Bufalin Induces Lung Cancer Cell Apoptosis via the Inhibition of PI3K/Akt Pathway

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
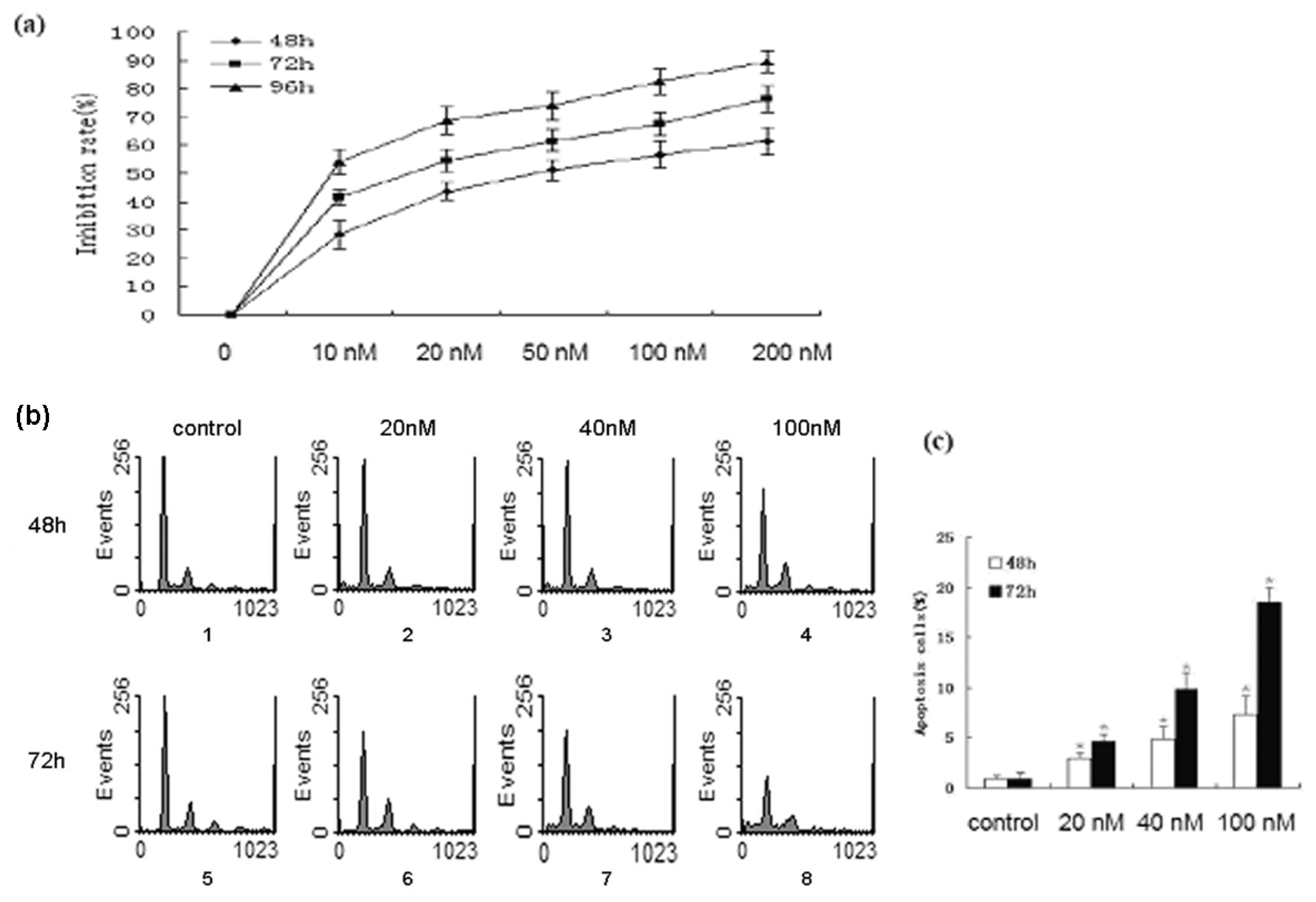
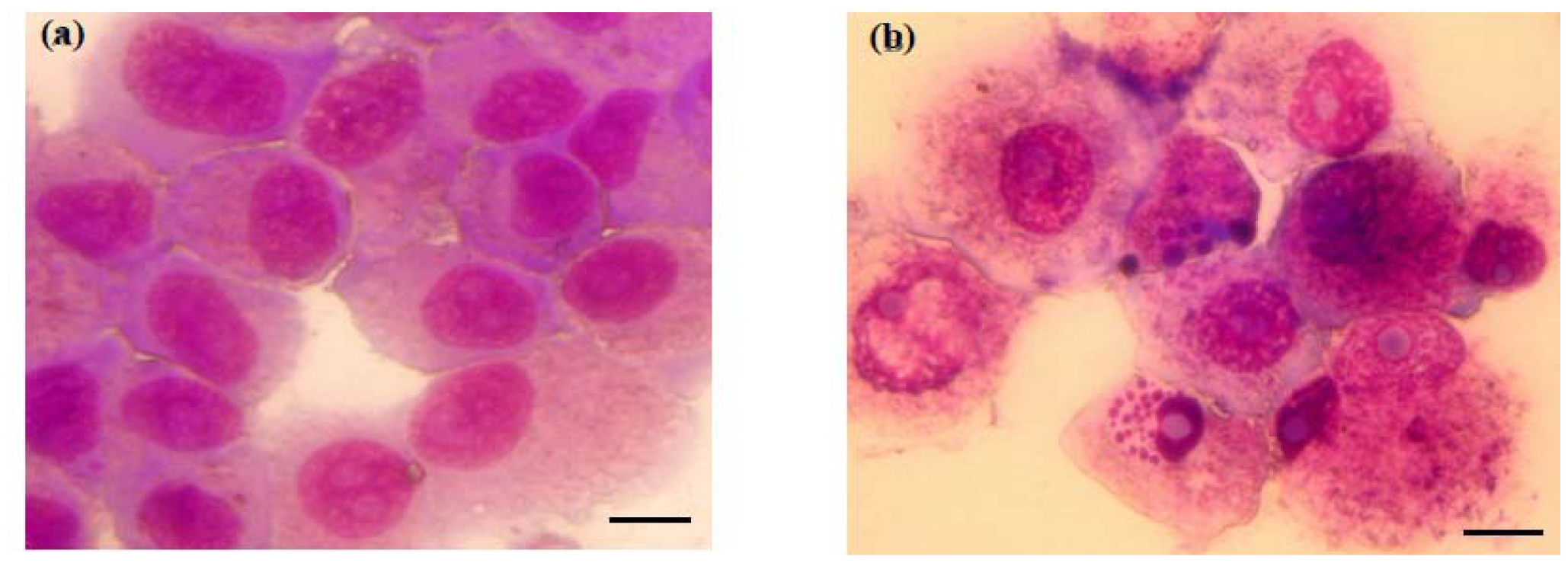
2.1. Bufalin Inhibits the Proliferation and Induces the Apoptosis of A549 Cells
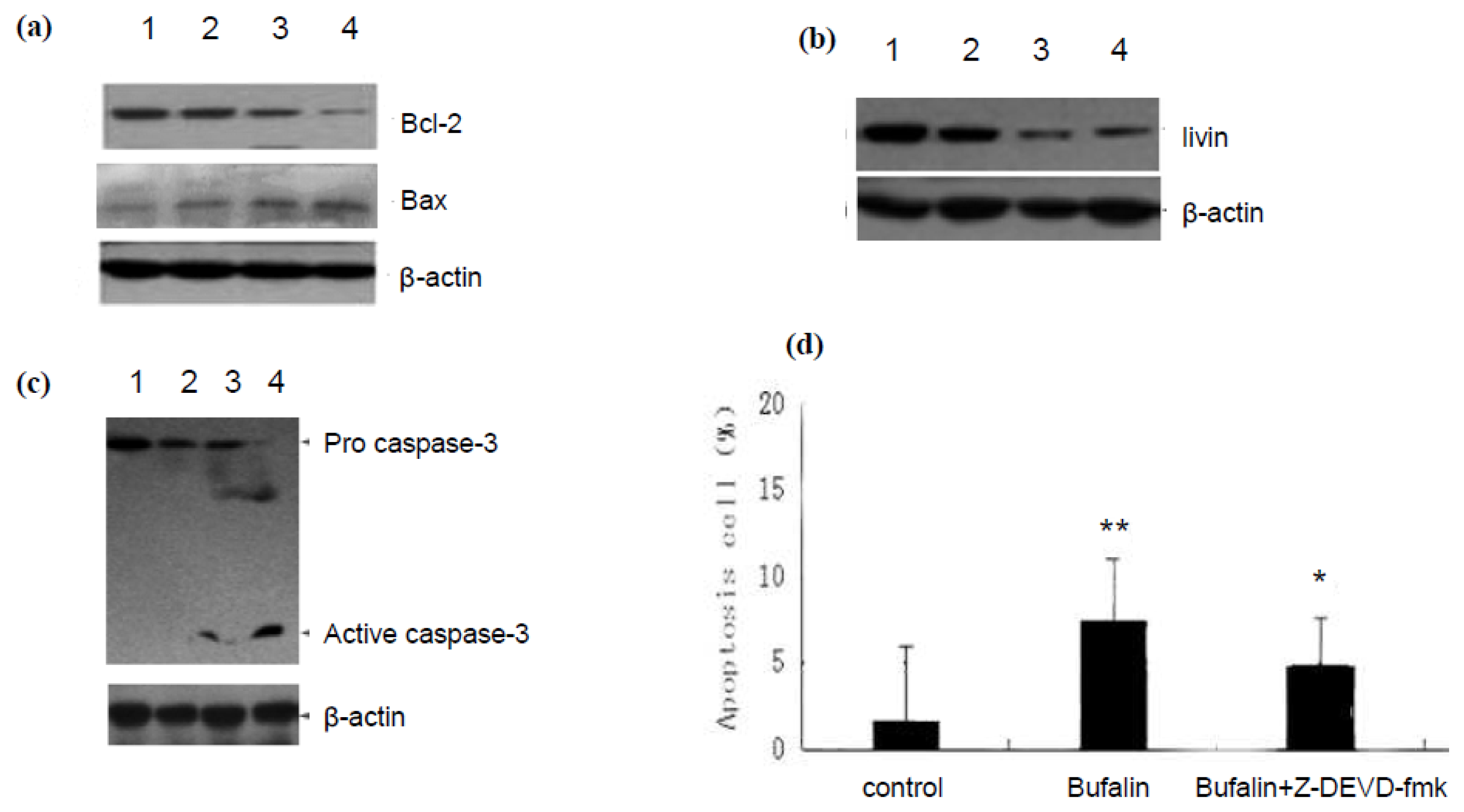
2.2. Bufalin Regulates the Expression of Apoptosis Related Proteins in A549 Cells
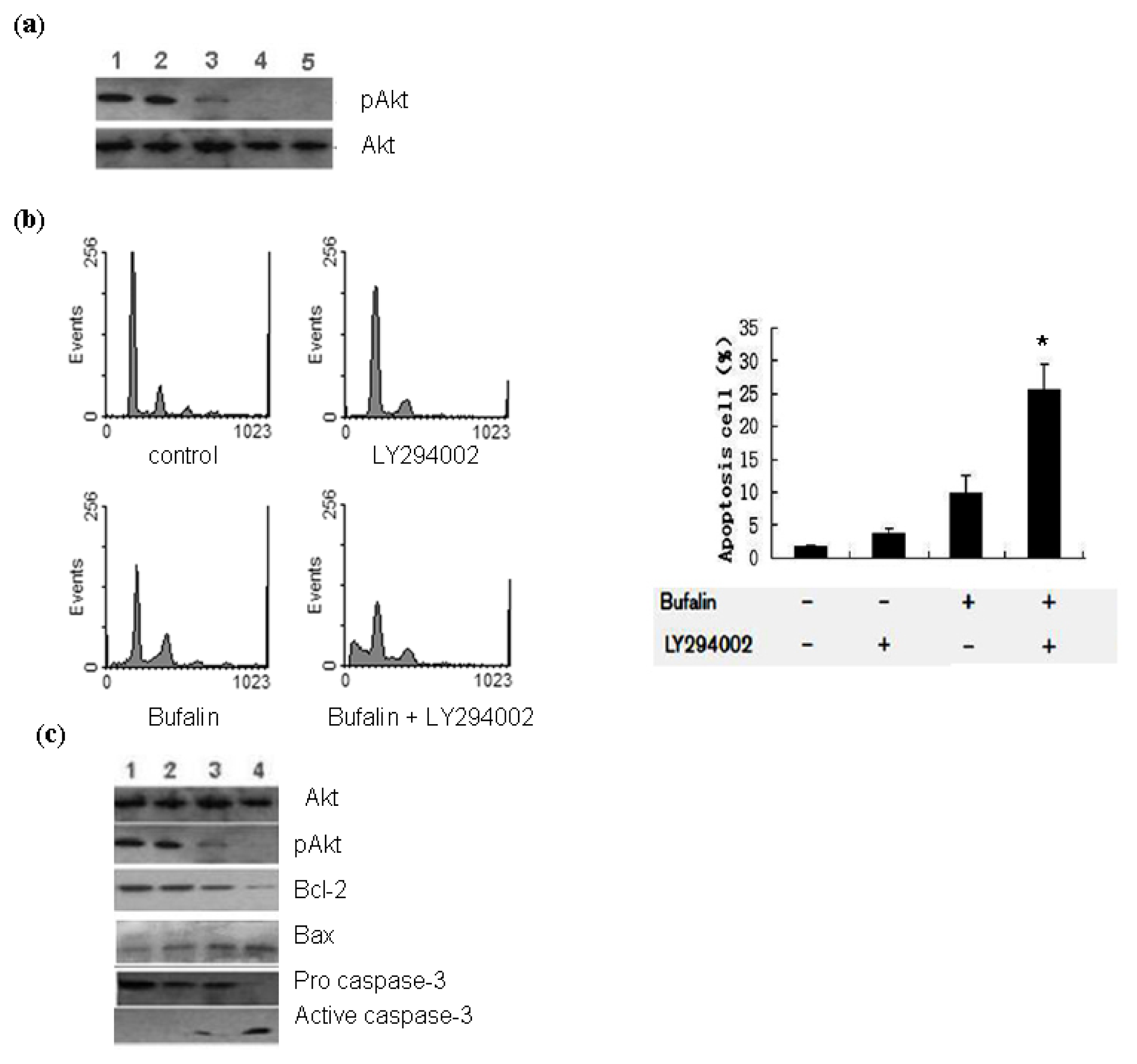
2.3. Bufalin Modulates the Activation of PI3K/Akt Pathway in A549 Cells
3. Materials and Methods
3.1. Reagents and Antibodies
3.2. Cell Culture
3.3. MTT Assay
3.4. Giemsa Staining
3.5. Flow Cytometry Analysis
3.6. Western Blot Analysis
3.7. Statistical Analysis
4. Conclusions
Acknowledgements
References
- Jemal, A.; Siegel, R.; Ward, E.; Hao, Y.; Xu, J.; Murray, T.; Thun, M.J. Cancer statistics. CA Cancer J. Clin. 2008, 58, 71–96. [Google Scholar]
- Pignon, J.P.; Tribodet, H.; Scagliotti, G.V.; Douillard, J.Y.; Shepherd, F.A.; Stephens, R.J.; Dunant, A.; Torri, V.; Rosell, R.; Seymour, L.; et al. Lung adjuvant cisplatin evaluation: A pooled analysis by the LACE Collaborative Group. J. Clin. Oncol 2008, 26, 3552–3559. [Google Scholar]
- Strauss, G.M.; Herndon, J.E.; Maddaus, M.A.; Johnstone, D.W.; Johnson, E.A.; Harpole, D.H.; Gillenwater, H.H.; Watson, D.M.; Sugarbaker, D.J.; Schilsky, R.L.; et al. Adjuvant paclitaxel plus carboplatin compared with observation in stage IB non-small-cell lung cancer: CALGB 9633 with the Cancer and Leukemia Group B, Radiation Therapy Oncology Group, and North Central Cancer Treatment Group Study Groups. J. Clin. Oncol 2008, 26, 5043–5051. [Google Scholar]
- Qi, F.; Inagaki, Y.; Gao, B.; Cui, X.; Xu, H.; Kokudo, N.; Li, A.; Tang, W. Bufalin and cinobufagin induce apoptosis of human hepatocellular carcinoma cells via Fas- and mitochondria-mediated pathways. Cancer Sci 2011, 102, 951–958. [Google Scholar]
- Chen, A.; Yu, J.; Zhang, L.; Sun, Y.; Zhang, Y.; Guo, H.; Zhou, Y.; Mitchelson, K.; Cheng, J. Microarray and biochemical analysis of bufalin-induced apoptosis of HL-60 Cells. Biotechnol. Lett 2009, 31, 487–494. [Google Scholar]
- Takai, N.; Ueda, T.; Nishida, M.; Nasu, K.; Narahara, H. Bufalin induces growth inhibition, cell cycle arrest and apoptosis in human endometrial and ovarian cancer cells. Int. J. Mol. Med 2008, 21, 637–643. [Google Scholar]
- Xie, C.M.; Chan, W.Y.; Yu, S.; Zhao, J.; Cheng, C.H. Bufalin induces autophagy-mediated cell death in human colon cancer cells through reactive oxygen species generation and JNK activation. Free Radic. Biol. Med 2011, 51, 1365–1375. [Google Scholar]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar]
- Li, D.; Qu, X.; Hou, K.; Zhang, Y.; Dong, Q.; Teng, Y.; Zhang, J.; Liu, Y. PI3K/Akt is involved in bufalin-induced apoptosis in gastric cancer cells. Anticancer Drugs 2009, 20, 59–64. [Google Scholar]
- Takeuchi, H.; Kim, J.; Fujimoto, A.; Umetani, N.; Mori, T.; Bilchik, A.; Turner, R.; Tran, A.; Kuo, C.; Hoon, D.S. X-Linked inhibitor of apoptosis protein expression level in colorectal cancer is regulated by hepatocyte growth factor/C-met pathway via Akt signaling. Clin. Cancer Res 2005, 11, 7621–7628. [Google Scholar]
- Lee, S.M.; Lee, C.T.; Kim, Y.W.; Han, S.K.; Shim, Y.S.; Yoo, C.G. Hypoxia confers protection against apoptosis via PI3K/Akt and ERK pathways in lung cancer cells. Cancer Lett 2006, 242, 231–238. [Google Scholar]
- Bak, Y.; Kim, H.; Kang, J.W.; Lee, D.H.; Kim, M.S.; Park, Y.S.; Kim, J.H.; Jung, K.Y.; Lim, Y.; Hong, J.; Yoon, D.Y. A synthetic naringenin derivative, 5-hydroxy-7, 4′-diacetyloxyflavanone-N-phenyl hydrazone (N101-43), induces apoptosis through up-regulation of Fas/FasL expression and inhibition of PI3K/Akt signaling pathways in non-small-cell lung cancer cells. J. Agric. Food Chem 2011, 59, 10286–10297. [Google Scholar]
- Li, W.; Wang, J.; Jiang, H.R.; Xu, X.L.; Zhang, J.; Liu, M.L.; Zhai, L.Y. Combined effects of cyclooxygenase-1 and cyclooxygenase-2 selective inhibitors on ovarian carcinoma in vivo. Int. J. Mol. Sci 2011, 12, 668–681. [Google Scholar]
- Khoo, B.Y.; Chua, S.L.; Balaram, P. Apoptotic effects of chrysin in human cancer cell lines. Int. J. Mol. Sci 2010, 11, 2188–2199. [Google Scholar]
- Donovan, M.; Cotter, T.G. Control of mitochondrial integrity by Bcl-2 family members and caspase-independent cell death. Biochim. Biophys. Acta 2004, 1644, 133–147. [Google Scholar]
- Watabe, M.; Kawazoe, N.; Masuda, Y.; Nakajo, S.; Nakaya, K. Bcl-2 protein inhibits bufalin-induced apoptosis through inhibition of mitogen-activated protein kinase activation in human leukemia U937 cells. Cancer Res 1997, 57, 3097–3100. [Google Scholar]
- Kasof, G.M.; Gomes, B.C. Livin, a novel inhibitor of apoptosis protein family member. J. Biol. Chem 2001, 276, 3238–3246. [Google Scholar]
- Sanna, M.G.; da Silva Correia, J.; Ducrey, O.; Lee, J.; Nomoto, K.; Schrantz, N.; Deveraux, Q.L.; Ulevitch, R.J. IAP suppression of apoptosis involves distinct mechanisms: The TAK1/JNK1 signaling cascade and caspase inhibition. Mol Cell Biol 2002, 22, 1754–1766. [Google Scholar]
- Liu, C.; Wu, X.; Luo, C.; Hu, Z.; Yin, Z.; He, Y.; Du, H.; Zhang, W.; Jiang, Q.; Lin, Y. Antisense oligonucleotide targeting Livin induces apoptosis of human bladder cancer cell via a mechanism involving caspase 3. J. Exp. Clin. Cancer Res 2010, 29. [Google Scholar] [CrossRef]
- Sos, M.L.; Fischer, S.; Ullrich, R.; Peifer, M.; Heuckmann, J.M.; Koker, M.; Heynck, S.; Stuckrath, I.; Weiss, J.; Fischer, F.; et al. Identifying genotype-dependent efficacy of single and combined PI3K- and MAPK-pathway inhibition in cancer. Proc. Natl. Acad. Sci. USA 2009, 106, 18351–18356. [Google Scholar]
- Li, Y.; Liu, J.; Liu, X.; Xing, K.; Wang, Y.; Li, F.; Yao, L. Resveratrol-induced cell inhibition of growth and apoptosis in MCF7 human breast cancer cells are associated with modulation of phosphorylated Akt and caspase-9. Appl. Biochem. Biotechnol 2006, 135, 181–192. [Google Scholar]
- Zhang, G.; Li, M.; Zhu, X.; Bai, Y.; Yang, C. Knockdown of akt sensitizes osteosarcoma cells to apoptosis induced by Cisplatin treatment. Int. J. Mol. Sci 2011, 12, 2994–3005. [Google Scholar]
- Song, L.; Xiong, H.; Li, J.; Liao, W.; Wang, L.; Wu, J.; Li, M. Sphingosine kinase-1 enhances resistance to apoptosis through activation of PI3K/Akt/NF-kappaB pathway in human non-small cell lung cancer. Clin. Cancer Res 2011, 17, 1839–1849. [Google Scholar]
- Watabe, M.; Masuda, Y.; Nakajo, S.; Yoshida, T.; Kuroiwa, Y.; Nakaya, K. The cooperative interaction of two different signaling pathways in response to bufalin induces apoptosis in human leukemia U937 cells. J. Biol. Chem 1996, 271, 14067–14072. [Google Scholar]




© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Zhu, Z.; Sun, H.; Ma, G.; Wang, Z.; Li, E.; Liu, Y.; Liu, Y. Bufalin Induces Lung Cancer Cell Apoptosis via the Inhibition of PI3K/Akt Pathway. Int. J. Mol. Sci. 2012, 13, 2025-2035. https://doi.org/10.3390/ijms13022025
Zhu Z, Sun H, Ma G, Wang Z, Li E, Liu Y, Liu Y. Bufalin Induces Lung Cancer Cell Apoptosis via the Inhibition of PI3K/Akt Pathway. International Journal of Molecular Sciences. 2012; 13(2):2025-2035. https://doi.org/10.3390/ijms13022025
Chicago/Turabian StyleZhu, Zhitu, Hongzhi Sun, Guangyou Ma, Zhenghua Wang, Enze Li, Yangyang Liu, and Yunpeng Liu. 2012. "Bufalin Induces Lung Cancer Cell Apoptosis via the Inhibition of PI3K/Akt Pathway" International Journal of Molecular Sciences 13, no. 2: 2025-2035. https://doi.org/10.3390/ijms13022025