Sequence Analysis and Potentials of the Native RbcS Promoter in the Development of an Alternative Eukaryotic Expression System Using Green Microalga Ankistrodesmus convolutus

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Sequence Analysis of AcRbcS Promoter
2.2. Determination of the Transcription Start Site
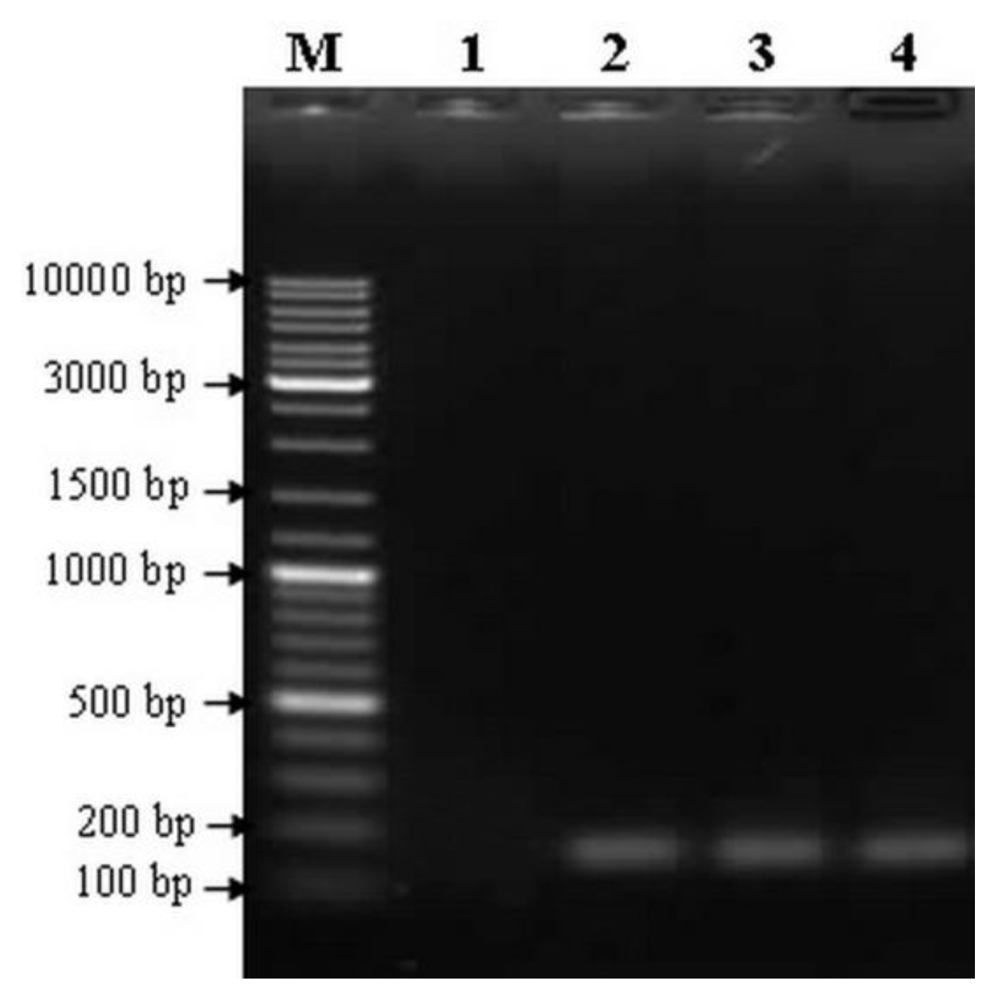
2.3. Detection of Transgenes Using PCR Analysis
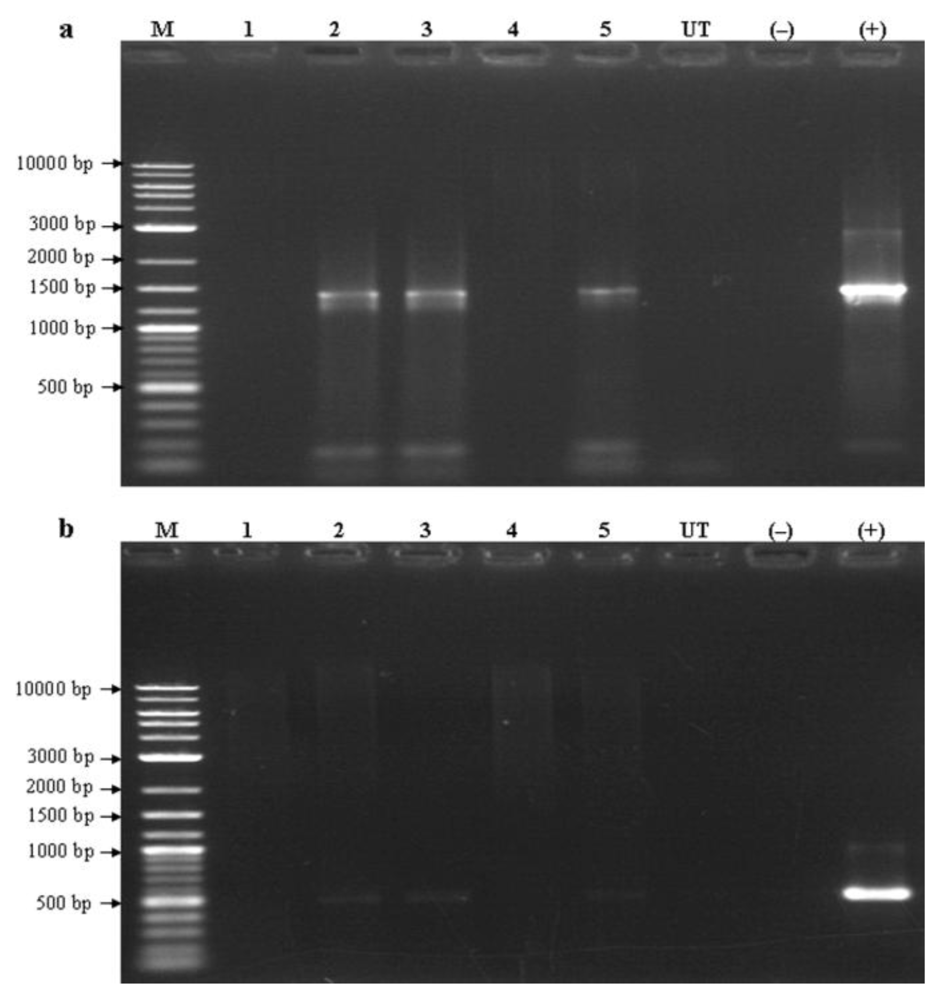
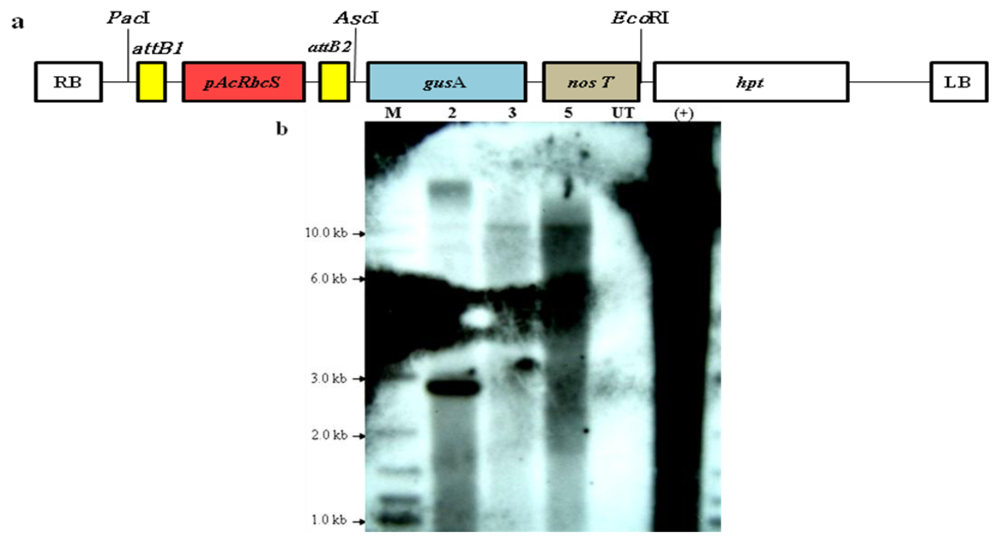
2.4. Integration of the Transgenes Driven by the AcRbcS Promoter
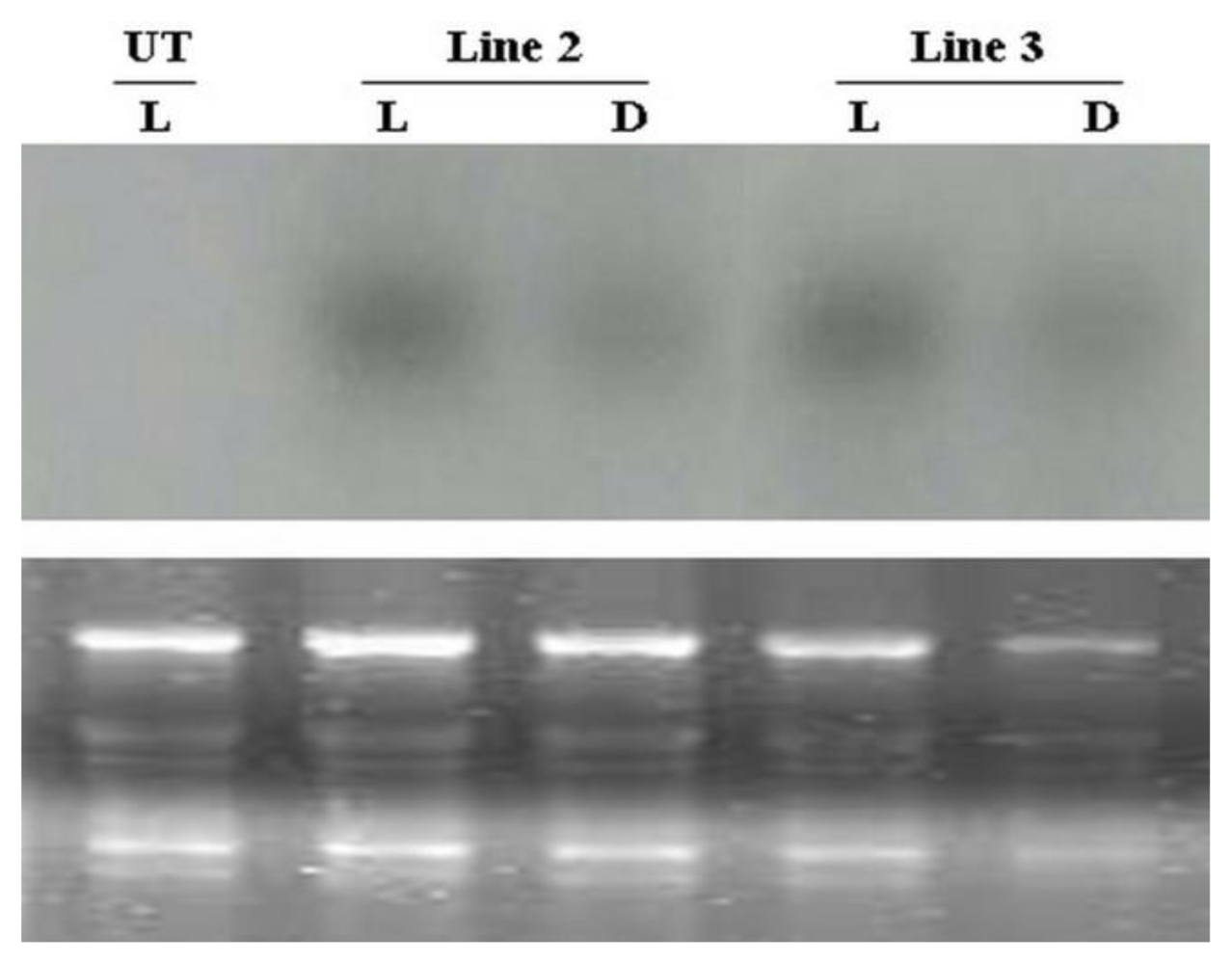
2.5. Influence of Light on the Regulation of AcRbcS Promoter Activity
3. Experimental Section
3.1. Culture Conditions
3.2. Isolation and Sequence Analysis of AcRbcS Promoter
3.3. Identification of the Transcription Start Site by 5′-Rapid Amplification of cDNA Ends (5′-RACE)
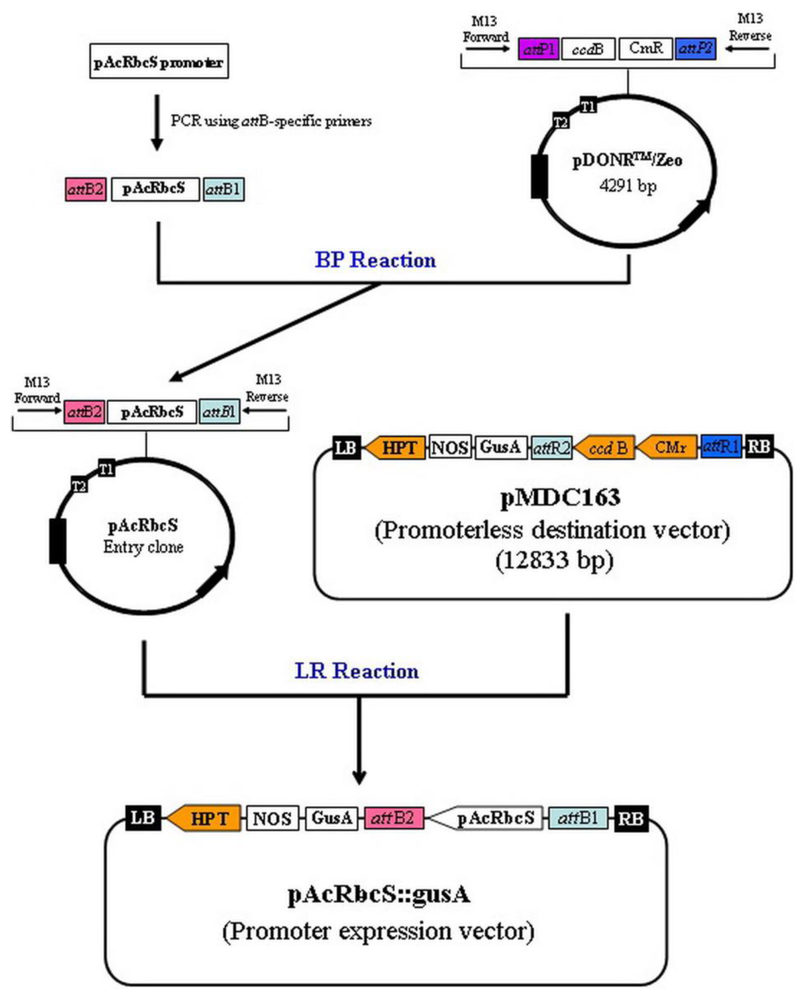
3.4. Construction of Promoter::gusA Fusing Vector
3.5. Nuclear Transformation of A. convolutus
3.6. Analysis of T-DNA Integration and Transgenes Expression
4. Conclusions
Acknowledgments
References
- Mayfield, S.P.; Franklin, S.E.; Lerner, R.A. Expression and assembly of a fully active antibody in algae. Proc. Natl. Acad. Sci. USA 2003, 100, 438–442. [Google Scholar]
- Griesbeck, C.; Kobl, I.; Heitzer, M. Chlamydomonas reinhardtii: A protein expression system for pharmaceutical and biotechnological proteins. Mol. Biotechnol 2006, 34, 213–223. [Google Scholar]
- Stevens, D.R.; Rochaix, J.D.; Purton, S. The bacterial phleomycin resistance gene ble as a dominant selectable marker in Chlamydomonas. Mol. Gen. Genet 1996, 251, 23–30. [Google Scholar]
- Schroda, M.; Blocker, D.; Beck, C.F. The HSP70A promoter as a tool for the improved expression of transgenes in Chlamydomonas. Plant J 2000, 21, 121–131. [Google Scholar]
- Walker, T.L.; Becker, D.K.; Collet, C. Characterisation of the Dunaliella tertiolecta RbcS genes and their promoter activity in Chlamydomonas reinhardtii. Plant Cell Rep 2005, 23, 727–735. [Google Scholar]
- Lu, Y.; Li, J.; Xue, L.; Yan, H.; Yuan, H.; Wang, C. A duplicated carbonic anhydrase 1 (DCA1) promoter mediates the nitrate reductase gene switch of Dunaliella salina. J. Appl. Phycol 2010. [Google Scholar] [CrossRef]
- Tang, D.K.H.; Qiao, S.Y.; Wu, M. Insertion mutagenesis of Chlamydomonas reinhardtii by electroporation and heterologous DNA. Biochem. Mol. Biol. Int 1995, 36, 1025–1035. [Google Scholar]
- El-Sheekh, M.M. Stable transformation of the intact cells of Chlorella kessleri with high velocity microprojectiles. Biol. Plant 1999, 42, 209–216. [Google Scholar]
- Kim, D.H.; Kim, Y.T.; Cho, J.J.; Bae, J.H.; Hur, S.B.; Hwang, I.; Choi, T.J. Stable integration and functional expression of flounder growth hormone gene in transformed microalga, Chlorella ellipsoidea. Mar. Biotechnol 2002, 4, 63–73. [Google Scholar]
- Tan, C.; Qin, S.; Zhang, Q.; Jiang, P.; Zhao, F. Establishment of a microparticle bombardment transformation system for Dunaliella salina. J. Microbiol 2005, 43, 361–365. [Google Scholar]
- Feng, S.; Xue, L.; Liu, H.; Lu, P. Improvement of efficiency of genetic transformation for Dunaliella salina by glass beads method. Mol. Biol. Rep 2009, 36, 1433–1439. [Google Scholar]
- Blankenship, E.; Kindle, K. Expression of chimeric genes by the light-regulated cabII-1 promoter in Chlamydomonas reinhardtii: A cabII-1/nit1 gene functions as a dominant selectable marker in a nit1 nit2 strain. Mol. Cell. Biol 1992, 12, 5268–5279. [Google Scholar]
- Butanaev, A.M. Hygromicin phosphotransferase gene as a dominant selective marker for transformation of Chlamydomonas reinhardtii. Mol. Biol 1994, 28, 682–686. [Google Scholar]
- Teng, C.; Qin, S.; Liu, J.; Yu, D.; Liang, C.; Tseng, C. Transient expression of lacZ in bombarded unicellular green alga Haematococcus pluvialis. J. Appl. Phycol 2002, 14, 495–500. [Google Scholar]
- Chen, Y.; Wang, Y.; Sun, Y.; Zhang, L.; Li, W. Highly efficient expression of rabbit neutrophil peptide-1 gene in Chlorella ellipsoidea cells. Curr. Genet 2001, 39, 365–370. [Google Scholar]
- Franklin, S.; Ngo, B.; Efuet, E.; Mayfield, S.P. Development of a GFP reporter gene for Chlamydomonas reinhardtii chloroplast. Plant J 2002, 30, 733–744. [Google Scholar]
- Shao, N.; Bock, R. A codon-optimized luciferase from Gaussia princeps facilitates the in vivo monitoring of gene expression in the model alga Chlamydomonas reinhardtii. Curr. Genet 2008, 53, 381–388. [Google Scholar]
- Fischer, N.; Rochaix, J.D. The flanking regions of PsaD drive efficient gene expression in the nucleus of the green alga Chlamydomonas reinhardtii. Mol. Genet. Genomics 2001, 265, 888–894. [Google Scholar]
- Kindle, K.L. High-frequency nuclear transformation of Chlamydomonas reinhardtii. Proc. Natl. Acad. Sci. USA 1990, 87, 1228–1232. [Google Scholar]
- Cerutti, H.; Johnson, A.M.; Gillham, N.W.; Boynton, J.E. A eubacterial gene conferring spectinomycin resistance on Chlamydomonas reinhardtii: integration into the nuclear genome and gene expression. Genetics 1997, 145, 97–110. [Google Scholar]
- Auchincloss, A.H.; Loroch, A.I.; Rochaix, J.D. The argininosuccinate lyase gene of Chlamydomonas reinhardtii: cloning of the cDNA and its characterization as a selectable shuttle marker. Mol. Gen. Genet 1999, 261, 21–30. [Google Scholar]
- Fuhrmann, M.; Oertel, W.; Hegemann, P. A synthetic gene coding for the green fluorescent protein (GFP) is a versatile reporter in Chlamydomonas reinhardtii. Plant J 1999, 19, 353–361. [Google Scholar]
- Kovar, J.L.; Zhang, J.; Funke, R.P.; Weeks, D.P. Molecular analysis of the acetolactate synthase gene of Chlamydomonas reinhardtii and development of a genetically engineered gene as a dominant selectable marker for genetic transformation. Plant J 2002, 29, 109–117. [Google Scholar]
- Sun, Y.; Yang, Z.Y.; Gao, X.S.; Li, Q.Y.; Zhang, Q.Q.; Xu, Z.K. Expression of foreign genes in Dunaliella by electroporation. Mol. Biotechnol 2005, 30, 185–192. [Google Scholar]
- Chu, W.L.; Phang, S.M.; Goh, S.H.; Blakebrough, N. Promising Microalgae for Production of Useful Chemicals. In Proceeding of the Conference on Medicinal Products from Tropical Rain Forests, Forest Research Institute of Malaysia, Kuala Lumpur, Malaysia, 13–15 May 1992; Shaari, K., Kadir, A.A., Ali, A.R.M., Eds.; pp. 338–345.
- Thanh, T.; Chi, V.T.Q.; Abdullah, M.P.; Omar, H.; Noroozi, M.; Ky, H.; Napis, S. Construction of cDNA library and preliminary analysis of expressed sequence tags from green microalga Ankistrodesmus convolutus Corda. Mol. Biol. Rep 2011, 38, 177–182. [Google Scholar]
- Giuliano, G.; Pechersky, E.; Malik, V.S.; Timko, M.P.; Scolnik, P.A.; Cashmore, A.R. An evolutionarily conserved protein binding sequence upstream of a plant light-regulated gene. Proc. Natl. Acad. Sci. USA 1988, 85, 7089–7093. [Google Scholar]
- Williams, M.E.; Foster, R.; Chua, N.H. Sequences flanking the hexameric G-box core CACGTG affect the specificity of protein binding. Plant Cell 1992, 4, 485–496. [Google Scholar]
- Hudson, M.E.; Quail, P.H. Identification of promoter motifs involved in the network of phytochrome A-regulated gene expression by combined analysis of genomic sequence and microarray data. Plant Physiol 2003, 133, 1605–1616. [Google Scholar]
- Li, T.; Gong, C.; Wang, T. The rice light-regulated gene RA68 encodes a novel protein interacting with oxygen-evolving complex PsbO mature protein. Plant Mol. Biol. Rep 2010, 28, 136–143. [Google Scholar]
- Ahrazem, O.; Rubio-Moraga, A.; López, R.C.; Gómez-Gómez, L. The expression of a chromoplast-specific lycopene beta cyclase gene is involved in the high production of saffron’s apocarotenoid precursors. J. Exp. Bot 2010, 61, 105–119. [Google Scholar]
- Nomura, M.; Katayama, K.; Nishimura, A.; Ishida, Y.; Ohta, S.; Komari, T.; Miyao-Tokutomi, M.; Tajima, S.; Matsuoka, M. The promoter of rbcS in a C3 plant (rice) directs organ-specific, light-dependent expression in a C4 plant (maize), but does not confer bundle sheath cell-specific expression. Plant Mol. Biol 2000, 44, 99–106. [Google Scholar]
- Higuchi, R.; Dollinger, G.; Walsh, P.S.; Griffith, R. Simultaneous amplification and detection of specific DNA sequences. Nat. Biotechnol 1992, 10, 413–417. [Google Scholar]
- Kumar, S.C.; Misqitta, R.W.; Reddy, V.S.; Rao, B.J.; Rajam, M.V. Genetic transformation of the green alga Chlamydomonas reinhardtii by Agrobacterium tumefaciens. Plant Sci 2004, 166, 731–738. [Google Scholar]
- Kota, R.; Holton, T.A.; Henry, R.J. Detection of transgenes in crop plants using molecular beacon assays. Plant Mol. Biol. Rep 1999, 17, 363–370. [Google Scholar]
- Jin, W.W.; Li, Z.Y.; Fang, Q.; Altosaar, I.; Liu, L.H.; Song, Y.C. Fluorescence in situ hybridization analysis of alien genes in Agrobacterium-mediated Cry1A(b)-transformed rice. Ann. Bot 2002, 90, 31–36. [Google Scholar]
- Wang, J.; Oard, J.H. Rice ubiquitin promoters: Deletion analysis and potential usefulness in plant transformation systems. Plant Cell Rep 2003, 22, 129–134. [Google Scholar]
- Gittins, J.R.; Pellny, T.K.; Hiles, E.R.; Rosa, C.; Biricolti, S.; James, D.J. Transgene expression driven by heterologous ribulose-1,5-bisphosphate carboxylase/oxygenase small-subunit gene promoters in the vegetative tissues of apple (Malus pumila Mill.). Planta 2000, 210, 232–240. [Google Scholar]
- Marraccini, P.; Courjault, C.; Caillet, V.; Lausanne, F.; Lepage, B.; Rogers, W.J.; Tessereau, S.; Deshayes, A. Rubisco small subunit of Coffea arabica: cDNA sequence, gene cloning and promoter analysis in transgenic tobacco plants. Plant Physiol. Biochem 2003, 41, 17–25. [Google Scholar]
- Terzaghi, W.B.; Cashmore, A.R. Light-regulated transcription. Annu. Rev. Plant Physiol. Plant Mol. Biol 1995, 46, 445–474. [Google Scholar]
- Yamazaki, T.; Yamamoto, M.; Sakamoto, W.; Kawano, S. Isolation and molecular characterization of rbcS in the unicellular green alga Nannochloris bacillaris (Chlorophyta, Trebouxiophyceae). Phycol. Res 2005, 53, 67–76. [Google Scholar]
- Nichols, H.W. Growth Media-Freshwater. In Handbook of Phycological Methods: Culture Methods and Growth Measurements; Stein, J.R., Ed.; Cambridge University Press: Cambridge, UK, 1973; pp. 7–24. [Google Scholar]
- Thanh, T.; Chi, V.T.Q.; Abdullah, M.P.; Omar, H.; Napis, S. Efficiency of the ligation-mediated PCR and TAIL-PCR methods on isolation of RbcS promoter sequences from green microalga Ankistrodesmus convolutus. Mol. Biol 2011. In Press. [Google Scholar]
- Higo, K.; Ugawa, Y.; Iwamoto, M.; Korenaga, T. Plant cis-acting regulatory DNA elements (PLACE) database. Nucleic Acids Res 1999, 27, 297–300. [Google Scholar]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; van de Peer, Y.; Rouzé, P.; Rombauts, P. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res 2002, 30, 325–327. [Google Scholar]
- Thanh, T.; Omar, H.; Abdullah, M.P.; Chi, V.T.Q.; Noroozi, M.; Ky, H.; Napis, S. Rapid and effective method of RNA isolation from green microalga Ankistrodesmus convolutus. Mol. Biotechnol 2009, 43, 148–153. [Google Scholar]
- Thanh, T.; Chi, V.T.Q.; Abdullah, M.P.; Omar, H.; Noroozi, M.; Napis, S. Cloning and characterization of ribulose-1,5-bisphosphate carboxylase/oxygenase small subunit (RbcS) cDNA from green microalga Ankistrodesmus convolutus. Mol. Biol. Rep 2011. [Google Scholar] [CrossRef]






© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Thanh, T.; Chi, V.T.Q.; Omar, H.; Abdullah, M.P.; Napis, S. Sequence Analysis and Potentials of the Native RbcS Promoter in the Development of an Alternative Eukaryotic Expression System Using Green Microalga Ankistrodesmus convolutus. Int. J. Mol. Sci. 2012, 13, 2676-2691. https://doi.org/10.3390/ijms13032676
Thanh T, Chi VTQ, Omar H, Abdullah MP, Napis S. Sequence Analysis and Potentials of the Native RbcS Promoter in the Development of an Alternative Eukaryotic Expression System Using Green Microalga Ankistrodesmus convolutus. International Journal of Molecular Sciences. 2012; 13(3):2676-2691. https://doi.org/10.3390/ijms13032676
Chicago/Turabian StyleThanh, Tran, Vu Thi Quynh Chi, Hishamuddin Omar, Mohd Puad Abdullah, and Suhaimi Napis. 2012. "Sequence Analysis and Potentials of the Native RbcS Promoter in the Development of an Alternative Eukaryotic Expression System Using Green Microalga Ankistrodesmus convolutus" International Journal of Molecular Sciences 13, no. 3: 2676-2691. https://doi.org/10.3390/ijms13032676