All-Trans Retinoic Acid Treatment Is Associated with Prohibitin Expression in Renal Interstitial Fibrosis Rats

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
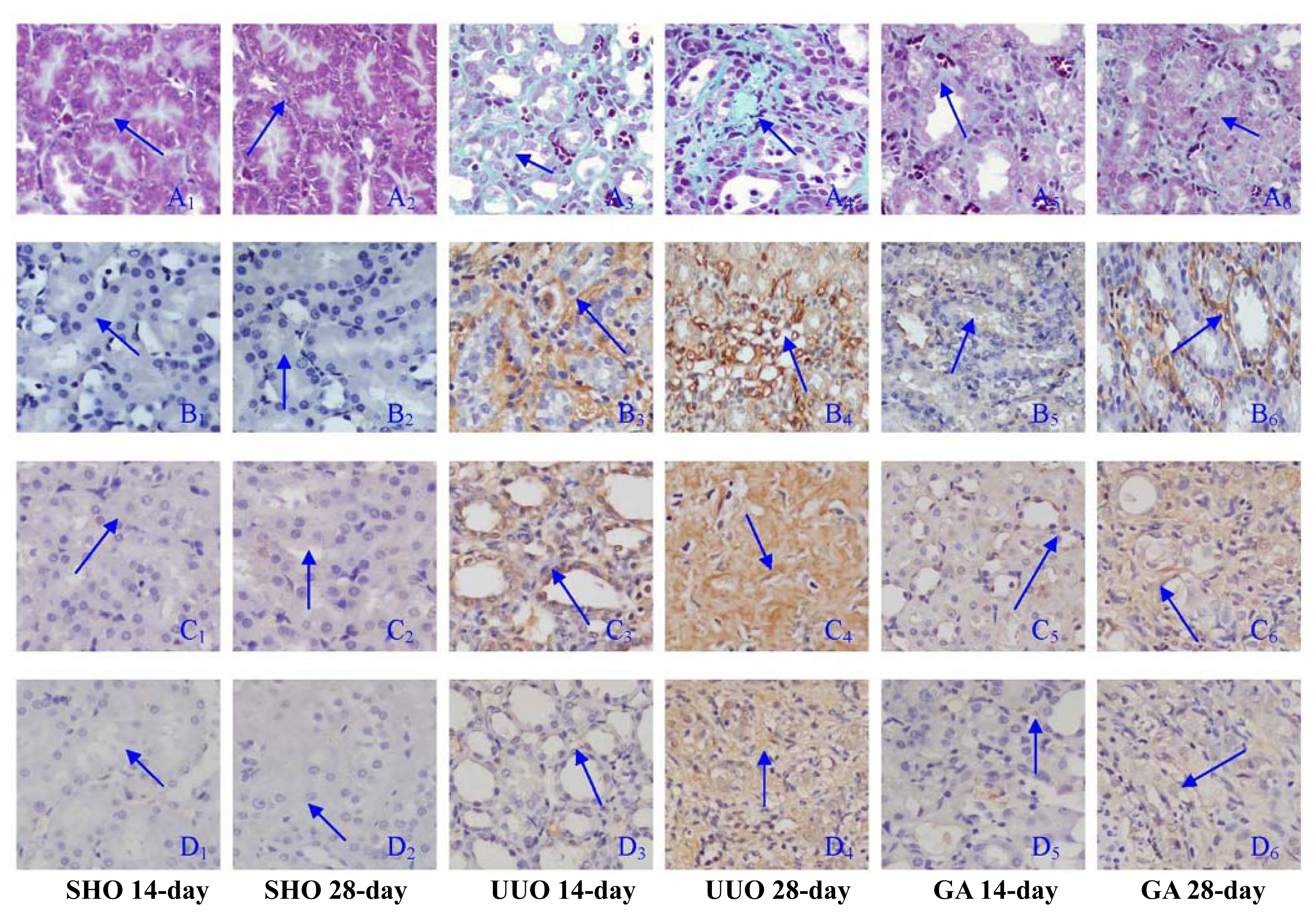
2.1. Renal Morphology
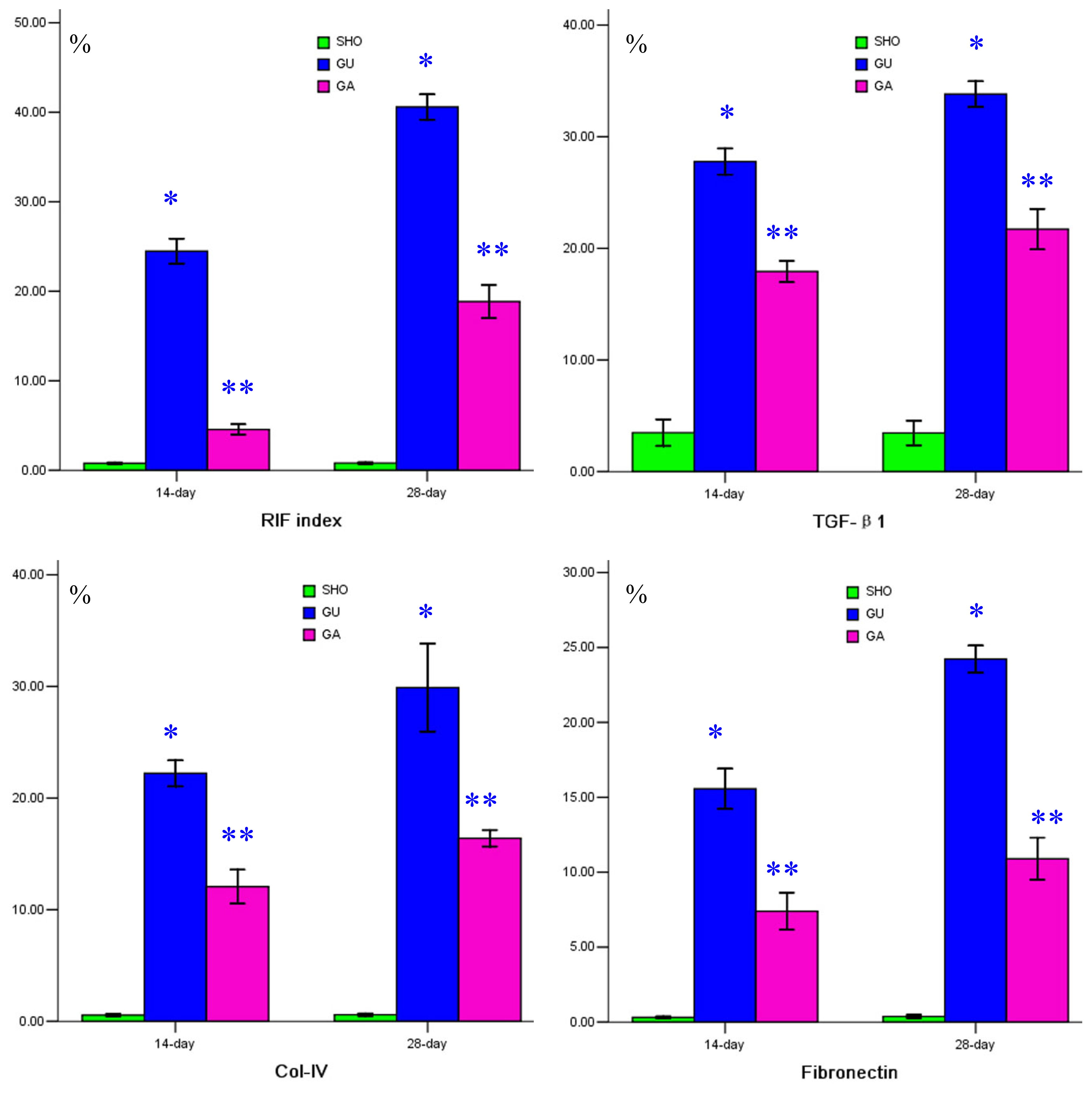
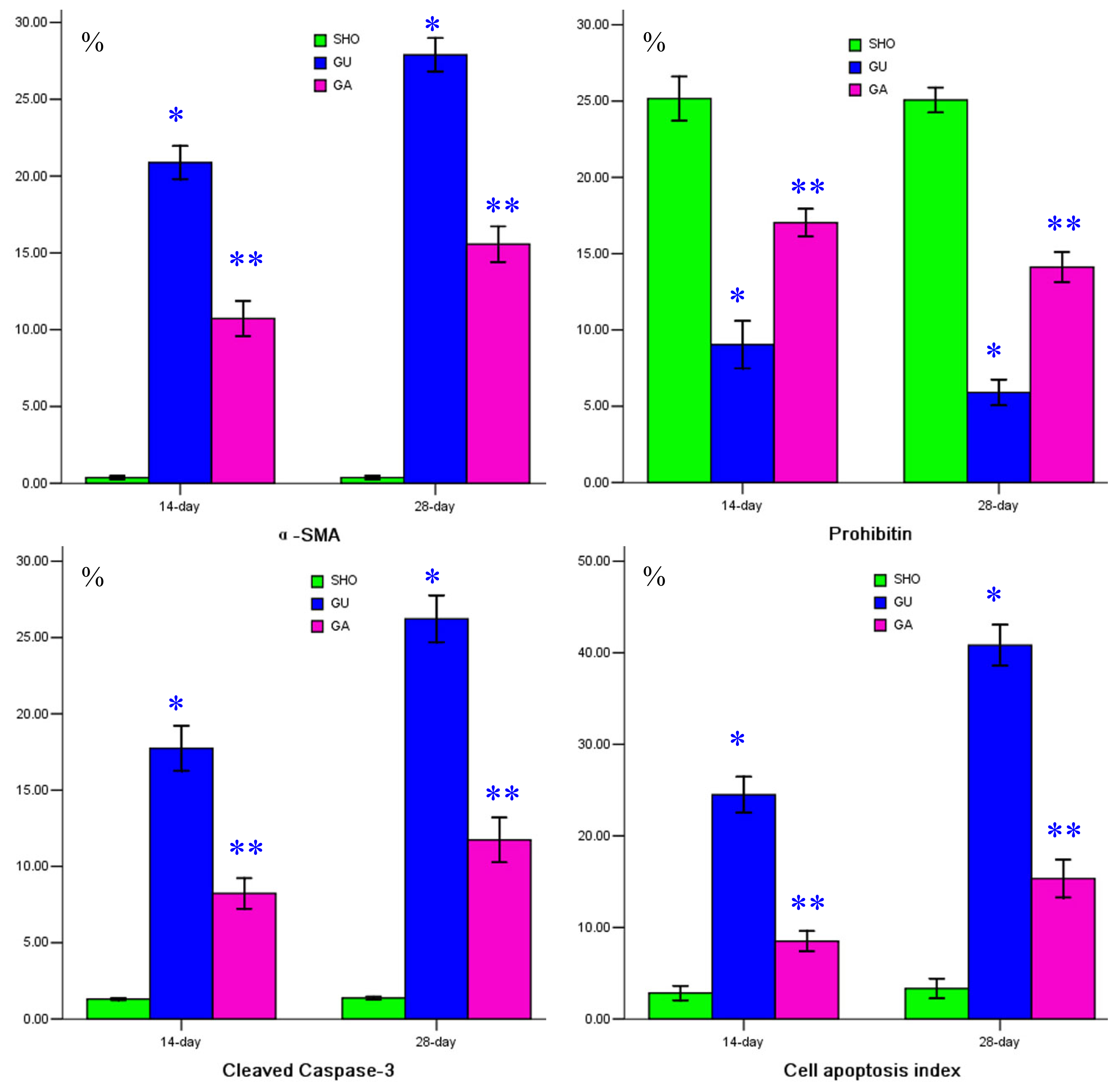
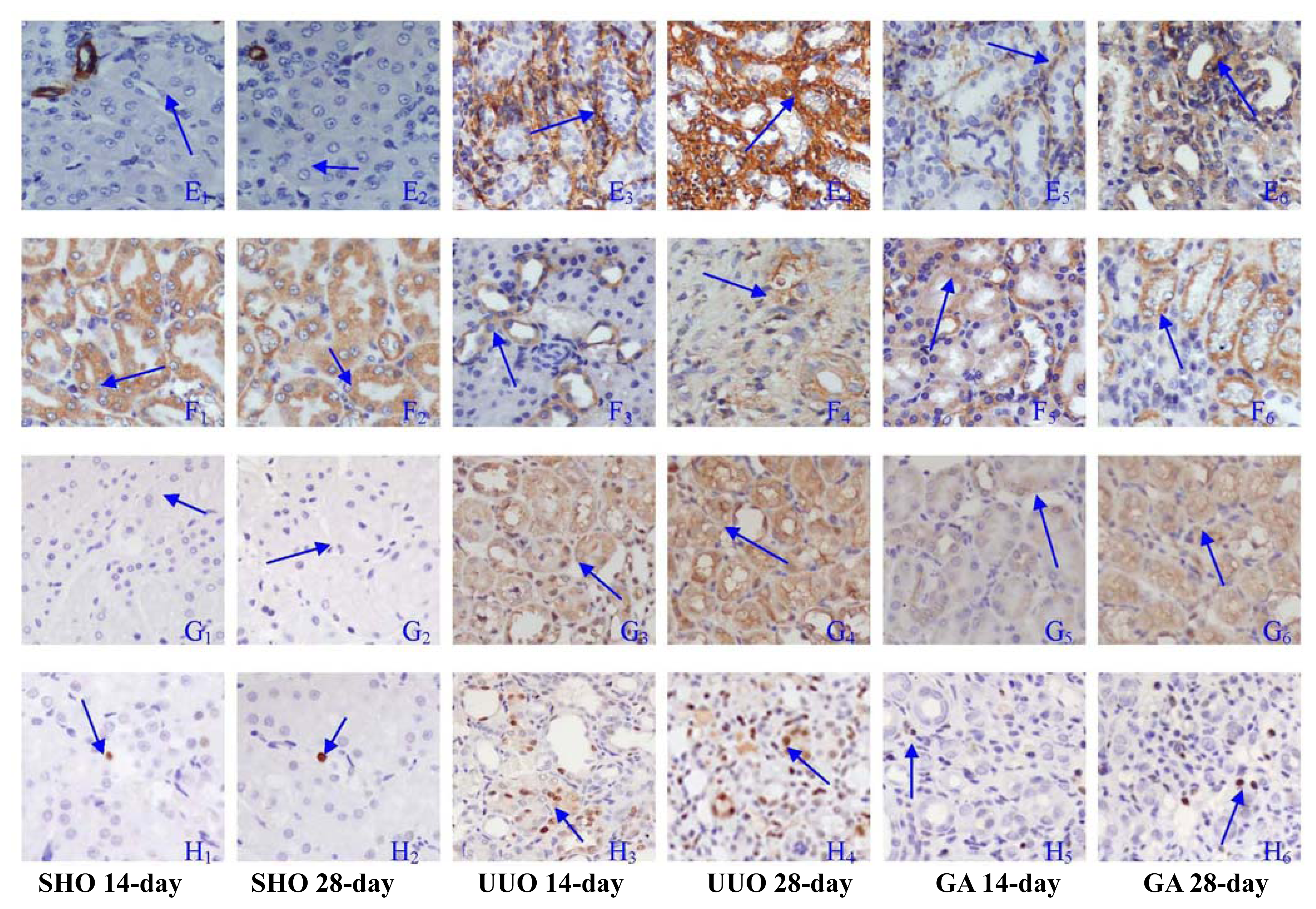
2.2. Association of ATRA Treatment with TGF-βl, Col-IV, Fibronectin, α-SMA, Prohibitin or Cleaved Caspase-3 Protein Expression
2.3. Association of ATRA Treatment with Cell Apoptosis
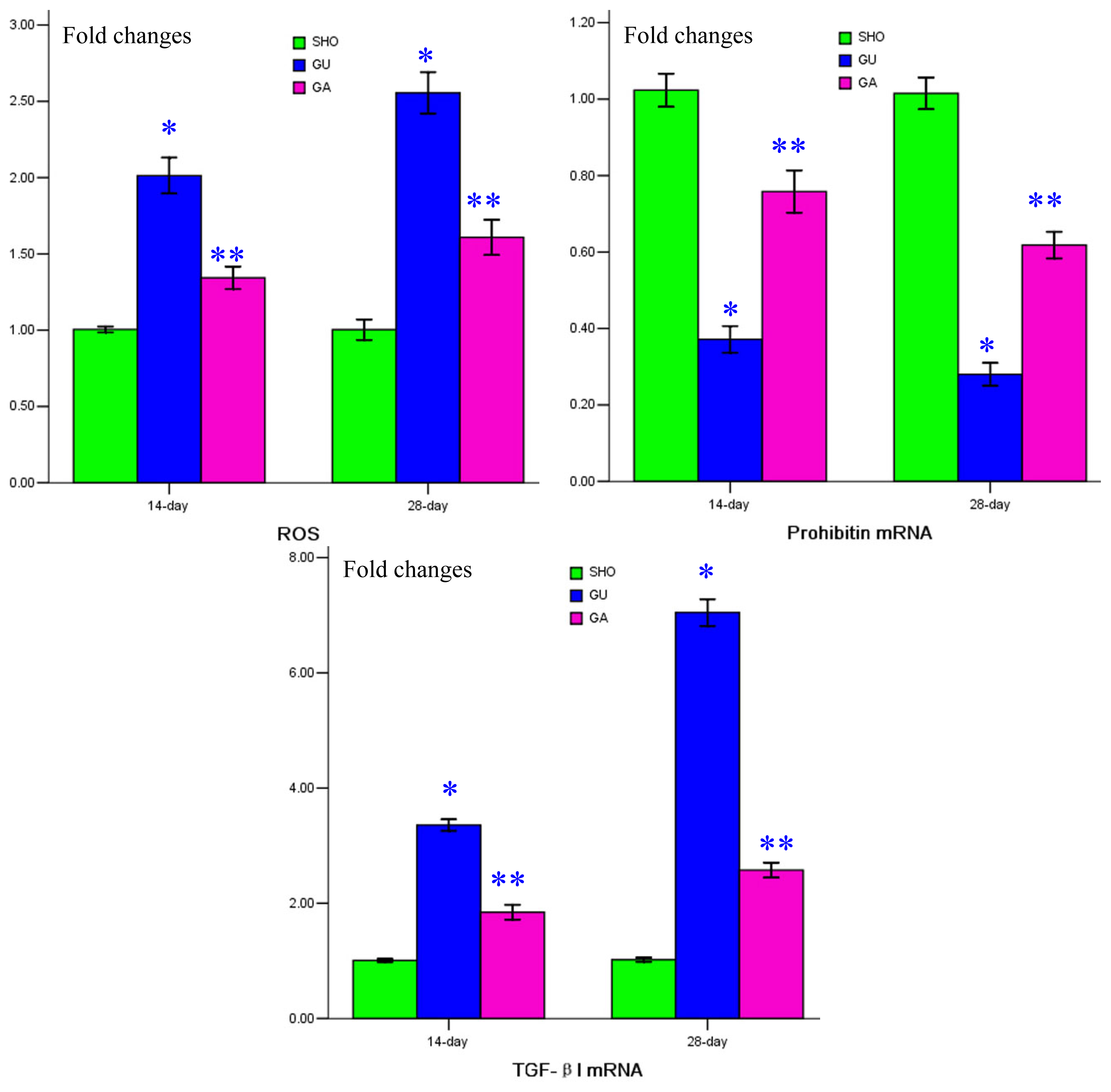
2.4. Association of ATRA Treatment with ROS Generation, mRNA Expressions of Prohibitin and TGF-βl
3. Discussion
4. Materials and Methods
4.1. Animal Model
4.2. Renal Morphology
4.3. Immunohistochemical Analysis of the Protein Expressions of Prohibitin, Transforming Growth Factor-β1 (TGF-β1), Collagen-IV (Col-IV), Fibronectin, α-Smooth Muscle Actin (α-SMA) and Cleaved Caspase-3
4.4. Apoptosis Assay
4.5. ROS Measurement
4.6. Real Time Reverse Transcription Polymerase Chain Reaction to Detect Prohibitin/TGF-β1 mRNA Expression in Renal Tissue
4.7. Statistical Analysis
5. Conclusion
Acknowledgments
- Conflict of InterestThe authors have declared that no competing interests exist.
References
- Bielesz, B.; Sirin, Y.; Si, H.; Niranjan, T.; Gruenwald, A.; Ahn, S.; Kato, H.; Pullman, J.; Gessler, M.; Haase, V.H.; et al. Epithelial Notch signaling regulates interstitial fibrosis development in the kidneys of mice and humans. J. Clin. Invest 2010, 120, 4040–4054. [Google Scholar]
- Kushiyama, T.; Oda, T.; Yamada, M.; Higashi, K.; Yamamoto, K.; Sakurai, Y.; Miura, S.; Kumagai, H. Alteration in the phenotype of macrophages in the repair of renal interstitial fibrosis in mice. Nephrology (Carlton) 2011, 16, 522–535. [Google Scholar]
- Morisada, N.; Nomura, M.; Nishii, H.; Furuno, Y.; Sakanashi, M.; Sabanai, K.; Toyohira, Y.; Ueno, S.; Watanabe, S.; Tamura, M.; et al. Complete disruption of all nitric oxide synthase genes causes markedly accelerated renal lesion formation following unilateral ureteral obstruction in mice in vivo. J. Pharmacol. Sci 2010, 114, 379–389. [Google Scholar]
- Correa-Costa, M.; Semedo, P.; Monteiro, A.P.; Silva, R.C.; Pereira, R.L.; Goncalves, G.M.; Marques, G.D.; Cenedeze, M.A.; Faleiros, A.C.; Keller, A.C.; et al. Induction of heme oxygenase-1 can halt and even reverse renal tubule-interstitial fibrosis. PLoS One 2010, 5, e14298. [Google Scholar]
- Manucha, W. Biochemical-molecular markers in unilateral ureteral obstruction. Biocell 2007, 31, 1–12. [Google Scholar]
- Manucha, W.; Carrizo, L.; Ruete, C.; Molina, H.; Valles, P. Angiotensin II type I antagonist on oxidative stress and heat shock protein 70 (HSP 70) expression in obstructive nephropathy. Cell Mol. Biol. (Noisy-le-Grand) 2005, 51, 547–555. [Google Scholar]
- Puppin, C.; Passon, N.; Franzoni, A.; Russo, D.; Damante, G. Histone deacetylase inhibitors control the transcription and alternative splicing of prohibitin in thyroid tumor cells. Oncol. Rep 2011, 25, 393–397. [Google Scholar]
- Zhu, B.; Zhai, J.; Zhu, H.; Kyprianou, N. Prohibitin regulates TGF-beta induced apoptosis as a downstream effector of Smad-dependent and -independent signaling. Prostate 2010, 70, 17–26. [Google Scholar]
- Li, Q.F.; Liang, Y.; Shi, S.L.; Liu, Q.R.; Xu, D.H.; Jing, G.J.; Wang, S.Y.; Kong, H.Y. Localization of prohibitin in the nuclear matrix and alteration of its expression during differentiation of human neuroblastoma SK-N-SH cells induced by retinoic acid. Cell. Mol. Neurobiol 2011, 31, 203–211. [Google Scholar]
- Liu, X.; Ren, Z.; Zhan, R.; Wang, X.; Wang, X.; Zhang, Z.; Leng, X.; Yang, Z.; Qian, L. Prohibitin protects against oxidative stress-induced cell injury in cultured neonatal cardiomyocyte. Cell Stress Chaperones 2009, 14, 311–319. [Google Scholar]
- Zhou, T.B.; Qin, Y.H.; Ou, C.; Lei, F.Y.; Su, L.N.; Huang, W.F.; Zhao, Y.J. All-trans retinoic acid can regulate the expressions of gelatinases and apolipoprotein E in glomerulosclerosis rats. Vascul. Pharmacol 2011, 55, 169–177. [Google Scholar]
- de Rosa, S.; Cirillo, P.; Paglia, A.; Sasso, L.; di Palma, V.; Chiariello, M. Reactive oxygen species and antioxidants in the pathophysiology of cardiovascular disease: Does the actual knowledge justify a clinical approach? Curr. Vasc. Pharmacol 2010, 8, 259–275. [Google Scholar]
- Lee, H.P.; Casadesus, G.; Zhu, X.; Lee, H.G.; Perry, G.; Smith, M.A.; Gustaw-Rothenberg, K.; Lerner, A. All-trans retinoic acid as a novel therapeutic strategy for Alzheimer’s disease. Expert Rev. Neurother 2009, 9, 1615–1621. [Google Scholar]
- Liu, X.; Lu, L.; Tao, B.B.; Zhou, A.L.; Zhu, Y.C. Amelioration of glomerulosclerosis with all-trans retinoic acid is linked to decreased plasminogen activator inhibitor-1 and alpha-smooth muscle actin. Acta Pharmacol. Sin 2011, 32, 70–78. [Google Scholar]
- Ratnam, K.K.; Feng, X.; Chuang, P.Y.; Verma, V.; Lu, T.C.; Wang, J.; Jin, Y.; Farias, E.F.; Napoli, J.L.; Chen, N.; et al. Role of the retinoic acid receptor-alpha in HIV-associated nephropathy. Kidney Int 2011, 79, 624–634. [Google Scholar]
- Zhou, T.B.; Qin, Y.H.; Lei, F.Y.; Su, L.N.; Zhao, Y.J.; Huang, W.F. All-trans retinoic acid regulates the expression of apolipoprotein E in rats with glomerulosclerosis induced by Adriamycin. Exp. Mol. Pathol 2011, 90, 287–294. [Google Scholar]
- Rao, J.; Zhang, C.; Wang, P.; Lu, L.; Zhang, F. All-trans retinoic acid alleviates hepatic ischemia/ reperfusion injury by enhancing manganese superoxide dismutase in rats. Biol. Pharm. Bull 2010, 33, 869–875. [Google Scholar]
- Chen, C.L.; Chou, K.J.; Lee, P.T.; Chen, Y.S.; Chang, T.Y.; Hsu, C.Y.; Huang, W.C.; Chung, H.M.; Fang, H.C. Erythropoietin suppresses epithelial to mesenchymal transition and intercepts Smad signal transduction through a MEK-dependent mechanism in pig kidney (LLC-PK1) cell lines. Exp. Cell Res 2010, 316, 1109–1118. [Google Scholar]
- Djamali, A.; Vidyasagar, A.; Yagci, G.; Huang, L.J.; Reese, S. Mycophenolic acid may delay allograft fibrosis by inhibiting transforming growth factor-beta1-induced activation of Nox-2 through the nuclear factor-κB pathway. Transplantation 2010, 90, 387–393. [Google Scholar]
- Zhou, G.; Li, C.; Cai, L. Advanced glycation end-products induce connective tissue growth factor-mediated renal fibrosis predominantly through transforming growth factor beta-independent pathway. Am. J. Pathol 2004, 165, 2033–2043. [Google Scholar]
- Kelley, R.; Werdin, E.S.; Bruce, A.T.; Choudhury, S.; Wallace, S.M.; Ilagan, R.M.; Cox, B.R.; Tatsumi-Ficht, P.; Rivera, E.A.; Spencer, T.; et al. Tubular cell-enriched subpopulation of primary renal cells improves survival and augments kidney function in rodent model of chronic kidney disease. Am. J. Physiol. Renal. Physiol 2010, 299, 1026–1039. [Google Scholar]
- Ko, K.S.; Tomasi, M.L.; Iglesias-Ara, A.; French, B.A.; French, S.W.; Ramani, K.; Lozano, J.J.; Oh, P.; He, L.; Stiles, B.L.; et al. Liver-specific deletion of prohibitin 1 results in spontaneous liver injury, fibrosis, and hepatocellular carcinoma in mice. Hepatology 2010, 52, 2096–2108. [Google Scholar]
- Sato, S.; Murata, A.; Orihara, T.; Shirakawa, T.; Suenaga, K.; Kigoshi, H.; Uesugi, M. Marine natural product aurilide activates the OPA1-mediated apoptosis by binding to prohibitin. Chem. Biol 2011, 18, 131–139. [Google Scholar]
- Theiss, A.L.; Laroui, H.; Obertone, T.S.; Chowdhury, I.; Thompson, W.E.; Merlin, D.; Sitaraman, S.V. Nanoparticle-based therapeutic delivery of prohibitin to the colonic epithelial cells ameliorates acute murine colitis. Inflamm. Bowel Dis 2011, 17, 1163–1176. [Google Scholar]
- Muraguchi, T.; Kawawa, A.; Kubota, S. Prohibitin protects against hypoxia-induced H9c2 cardiomyocyte cell death. Biomed. Res 2010, 31, 113–122. [Google Scholar]
- Kishimoto, K.; Kinoshita, K.; Hino, S.; Yano, T.; Nagare, Y.; Shimazu, H.; Nozaki, Y.; Sugiyama, M.; Ikoma, S.; Funauchi, M. Therapeutic effect of retinoic acid on unilateral ureteral obstruction model. Nephron Exp. Nephrol 2011, 118, 69–78. [Google Scholar]
- Xie, P.; Sun, L.; Nayak, B.; Haruna, Y.; Liu, F.Y.; Kashihara, N.; Kanwar, Y.S. C/EBP-β modulates transcription of tubulointerstitial nephritis antigen in obstructive uropathy. J. Am. Soc. Nephrol 2009, 20, 807–819. [Google Scholar]
- Zhang, D.; Sun, L.; Xian, W.; Liu, F.; Ling, G.; Xiao, L.; Liu, Y.; Peng, Y.; Haruna, Y.; Kanwar, Y.S. Low-dose paclitaxel ameliorates renal fibrosis in rat UUO model by inhibition of TGF-β/Smad activity. Lab. Invest 2010, 90, 436–447. [Google Scholar]
- Radford, M.G., Jr; Donadio, J.V., Jr; Bergstralh, E.J.; Grande, J.P. Predicting renal outcome in IgA nephropathy. J. Am. Soc. Nephrol. 1997, 8, 199–207. [Google Scholar]
- Zhou, T.B.; Qin, Y.H.; Zhou, C.; Lei, F.Y.; Zhao, Y.J.; Chen, J.; Su, L.N.; Huang, W.F. Less expression of prohibitin is associated with increased Caspase-3 expression and cell apoptosis in renal interstitial fibrosis rats. Nephrology (Carlton) 2012, 17, 189–196. [Google Scholar]
- Hu, X.; Zhou, X.; He, B.; Xu, C.; Wu, L.; Cui, B.; Wen, H.; Lu, Z.; Jiang, H. Minocycline protects against myocardial ischemia and reperfusion injury by inhibiting high mobility group box 1 protein in rats. Eur. J. Pharmacol 2010, 638, 84–89. [Google Scholar]
- Ha, T.; Hua, F.; Liu, X.; Ma, J.; McMullen, J.R.; Shioi, T.; Izumo, S.; Kelley, J.; Gao, X.; Browder, W.; et al. Lipopolysaccharide-induced myocardial protection against ischaemia/ reperfusion injury is mediated through a PI3K/Akt-dependent mechanism. Cardiovasc. Res 2008, 78, 546–553. [Google Scholar]
- Li, W.; Wang, J.; Jiang, H.R.; Xu, X.L.; Zhang, J.; Liu, M.L.; Zhai, L.Y. Combined effects of cyclooxygenase-1 and cyclooxygenase-2 selective inhibitors on ovarian carcinoma in vivo. Int. J. Mol. Sci 2011, 12, 668–681. [Google Scholar]
- Lai, J.Y.; Li, Y.T.; Wang, T.P. In vitro response of retinal pigment epithelial cells exposed to chitosan materials prepared with different cross-linkers. Int. J. Mol. Sci 2010, 11, 5256–5272. [Google Scholar]
- Olalekan, L.A.; Lawal, A.F.; Ologundudu, A.; Adeniran, O.Y.; Omonkhua, A.; Obi, F. Antioxidant effects of heated garlic juice on cadmium-induced liver damage in rats as compared to ascorbic acid. J. Toxicol. Sci 2011, 36, 549–557. [Google Scholar]
- Zhang, Y.; Peng, T.; Zhu, H.; Zheng, X.; Zhang, X.; Jiang, N.; Cheng, X.; Lai, X.; Shunnar, A.; Singh, M.; et al. Prevention of hyperglycemia-induced myocardial apoptosis by gene silencing of Toll-like receptor-4. J. Transl. Med 2010, 8. [Google Scholar] [CrossRef]
- Chen, N.; Ko, M. Schisandrin B-induced glutathione antioxidant response and cardioprotection are mediated by reactive oxidant species production in rat hearts. Biol. Pharm. Bull 2010, 33, 825–829. [Google Scholar]
- Zhou, T.B.; Qin, Y.H.; Lei, F.Y.; Su, L.N.; Zhao, Y.J.; Huang, W.F. Less gelatinases is associated with apolipoprotein E accumulation in glomerulosclerosis rats. Histol. Histopathol 2012, 27, 249–256. [Google Scholar]
- Zhou, T.B.; Qin, Y.H.; Lei, F.Y.; Zhao, Y.J.; Huang, W.F. Association of PAX2 with cell apoptosis in unilateral ureteral obstruction rats. Ren. Fail 2012, 34, 194–202. [Google Scholar]





© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Zhou, T.-B.; Qin, Y.-H.; Li, Z.-Y.; Xu, H.-L.; Zhao, Y.-J.; Lei, F.-Y. All-Trans Retinoic Acid Treatment Is Associated with Prohibitin Expression in Renal Interstitial Fibrosis Rats. Int. J. Mol. Sci. 2012, 13, 2769-2782. https://doi.org/10.3390/ijms13032769
Zhou T-B, Qin Y-H, Li Z-Y, Xu H-L, Zhao Y-J, Lei F-Y. All-Trans Retinoic Acid Treatment Is Associated with Prohibitin Expression in Renal Interstitial Fibrosis Rats. International Journal of Molecular Sciences. 2012; 13(3):2769-2782. https://doi.org/10.3390/ijms13032769
Chicago/Turabian StyleZhou, Tian-Biao, Yuan-Han Qin, Zheng-Yi Li, Hui-Ling Xu, Yan-Jun Zhao, and Feng-Ying Lei. 2012. "All-Trans Retinoic Acid Treatment Is Associated with Prohibitin Expression in Renal Interstitial Fibrosis Rats" International Journal of Molecular Sciences 13, no. 3: 2769-2782. https://doi.org/10.3390/ijms13032769
APA StyleZhou, T. -B., Qin, Y. -H., Li, Z. -Y., Xu, H. -L., Zhao, Y. -J., & Lei, F. -Y. (2012). All-Trans Retinoic Acid Treatment Is Associated with Prohibitin Expression in Renal Interstitial Fibrosis Rats. International Journal of Molecular Sciences, 13(3), 2769-2782. https://doi.org/10.3390/ijms13032769