Variation in Stability of Endogenous Reference Genes in Fallopian Tubes and Endometrium from Healthy and Ectopic Pregnant Women

Abstract
:1. Introduction
2. Results and Discussion
2.1. Patient Characteristics
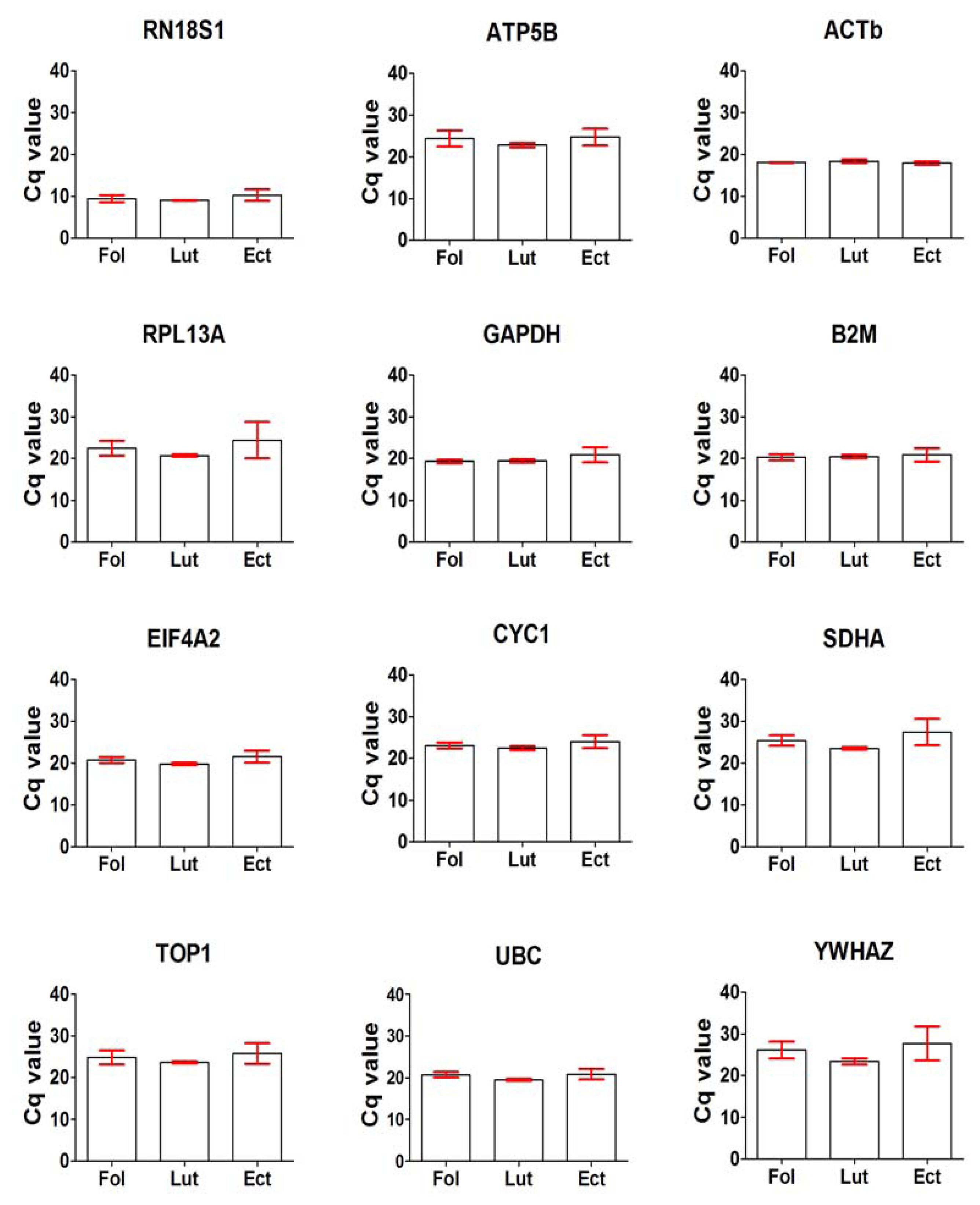
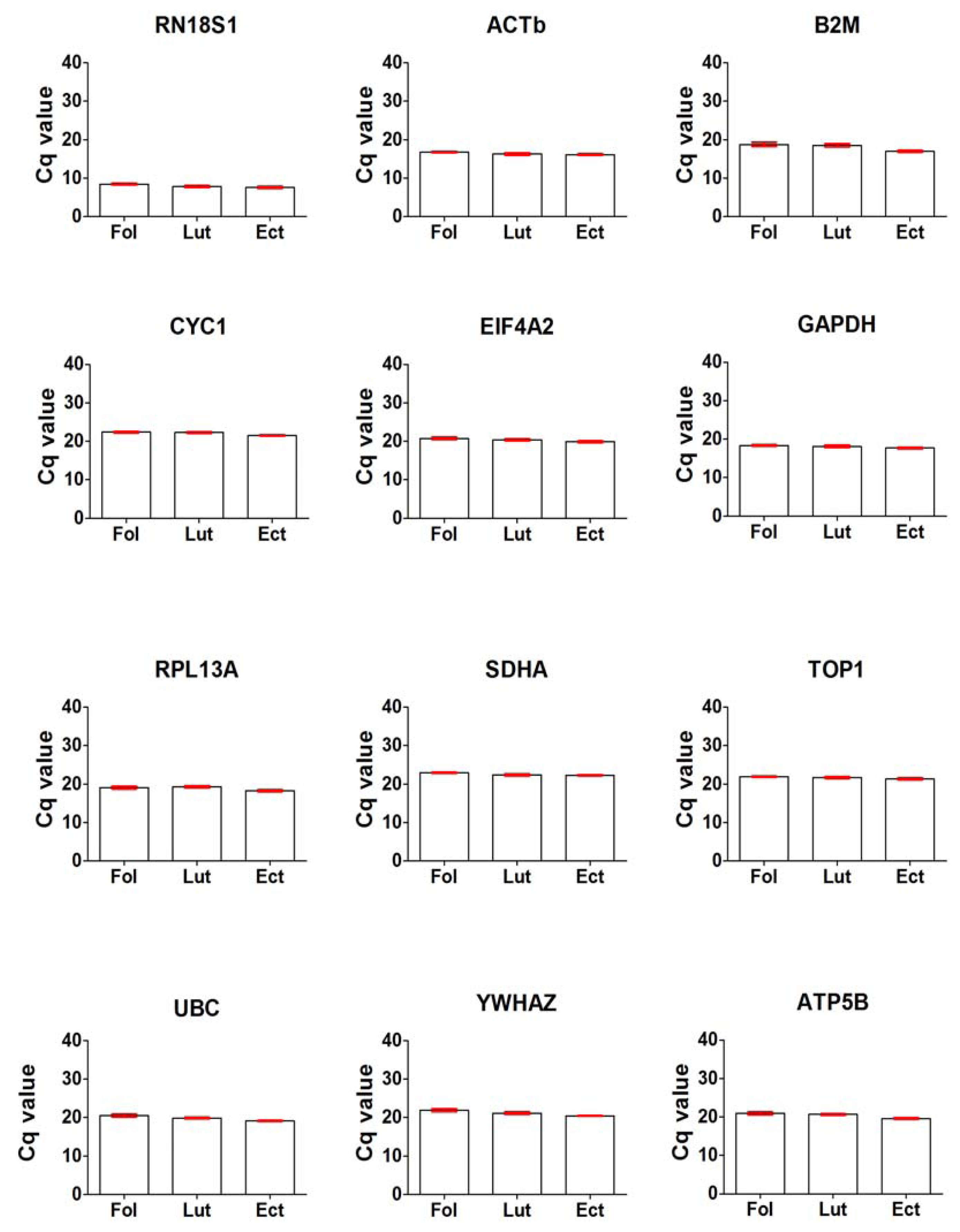
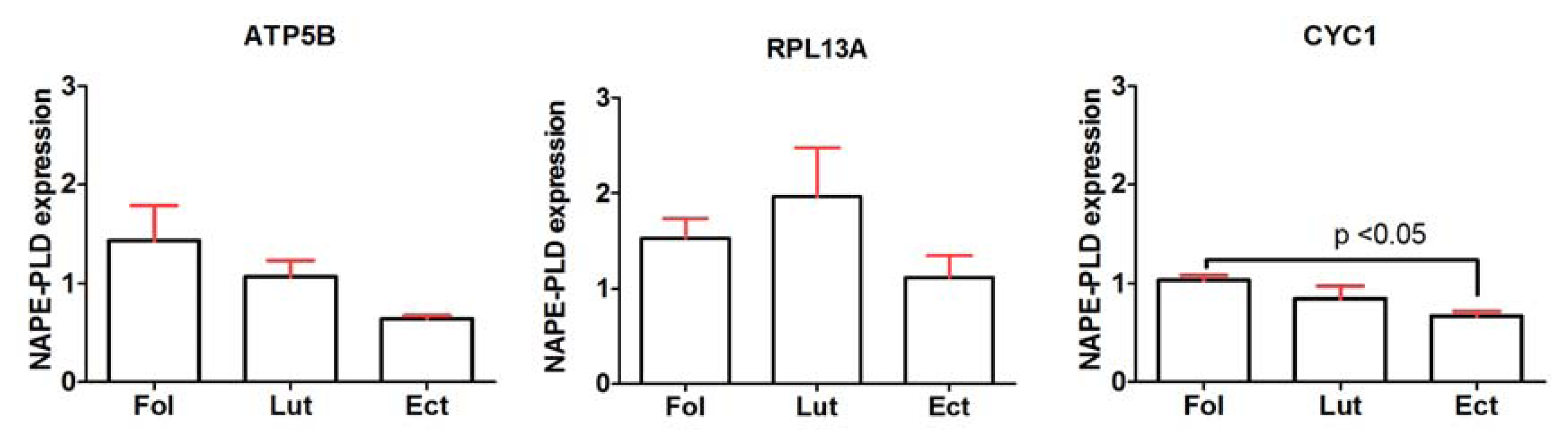
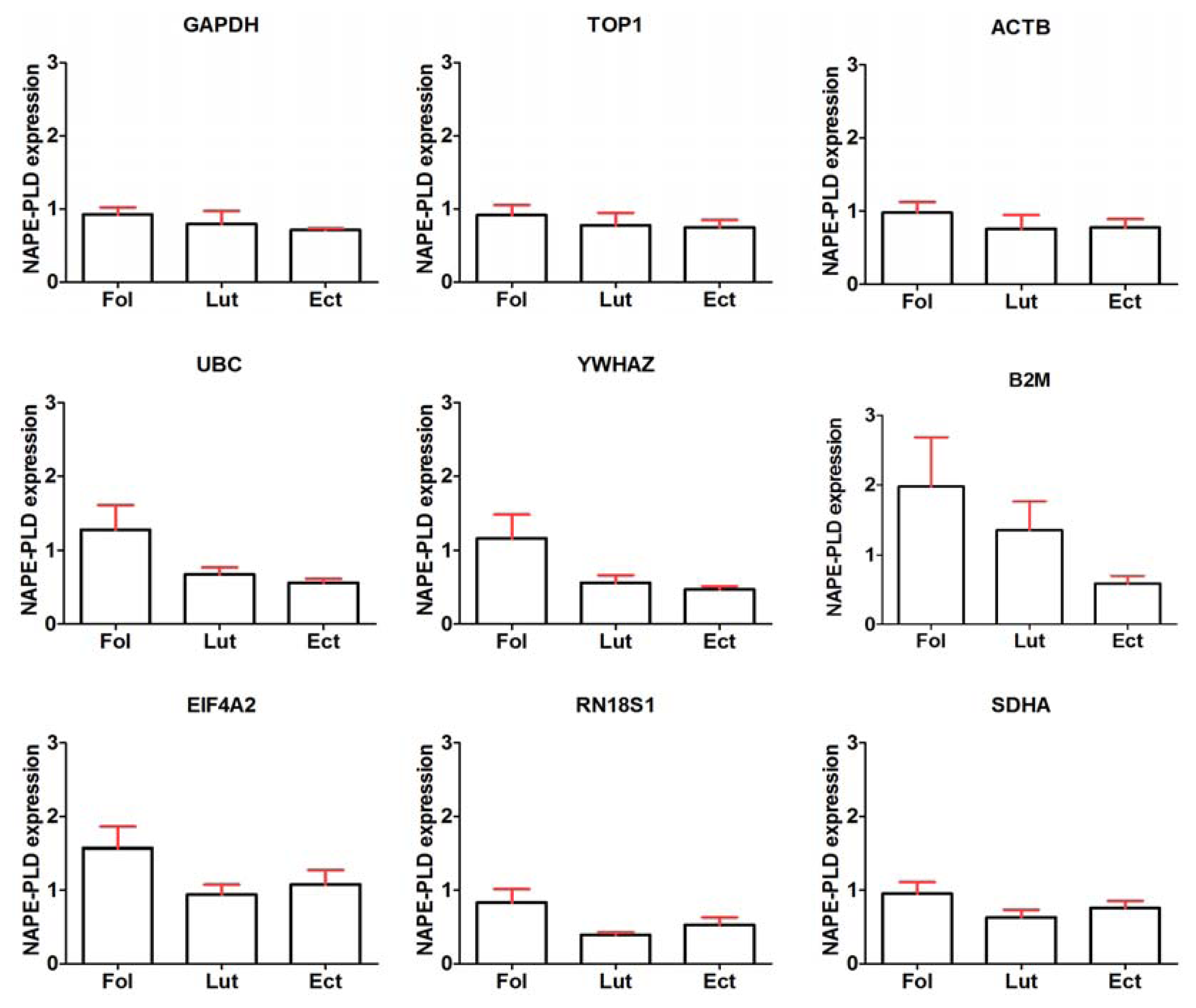
2.2. General Expression Levels of Endogenous Reference Genes
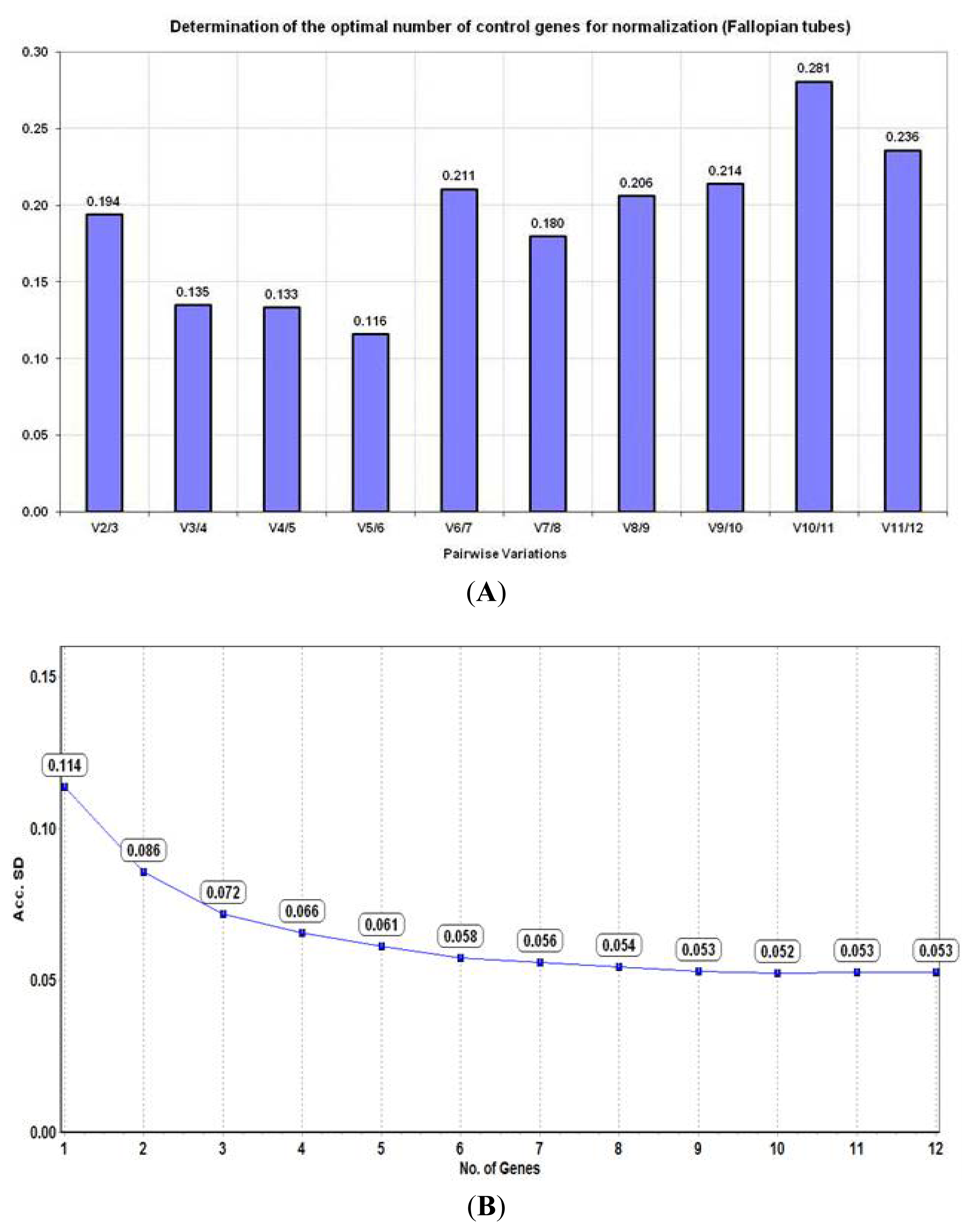
2.3. Analysis of Reference Gene Stability in Fallopian Tubes
2.3.1. geNorm Algorithm
2.3.2. NormFinder Algorithm
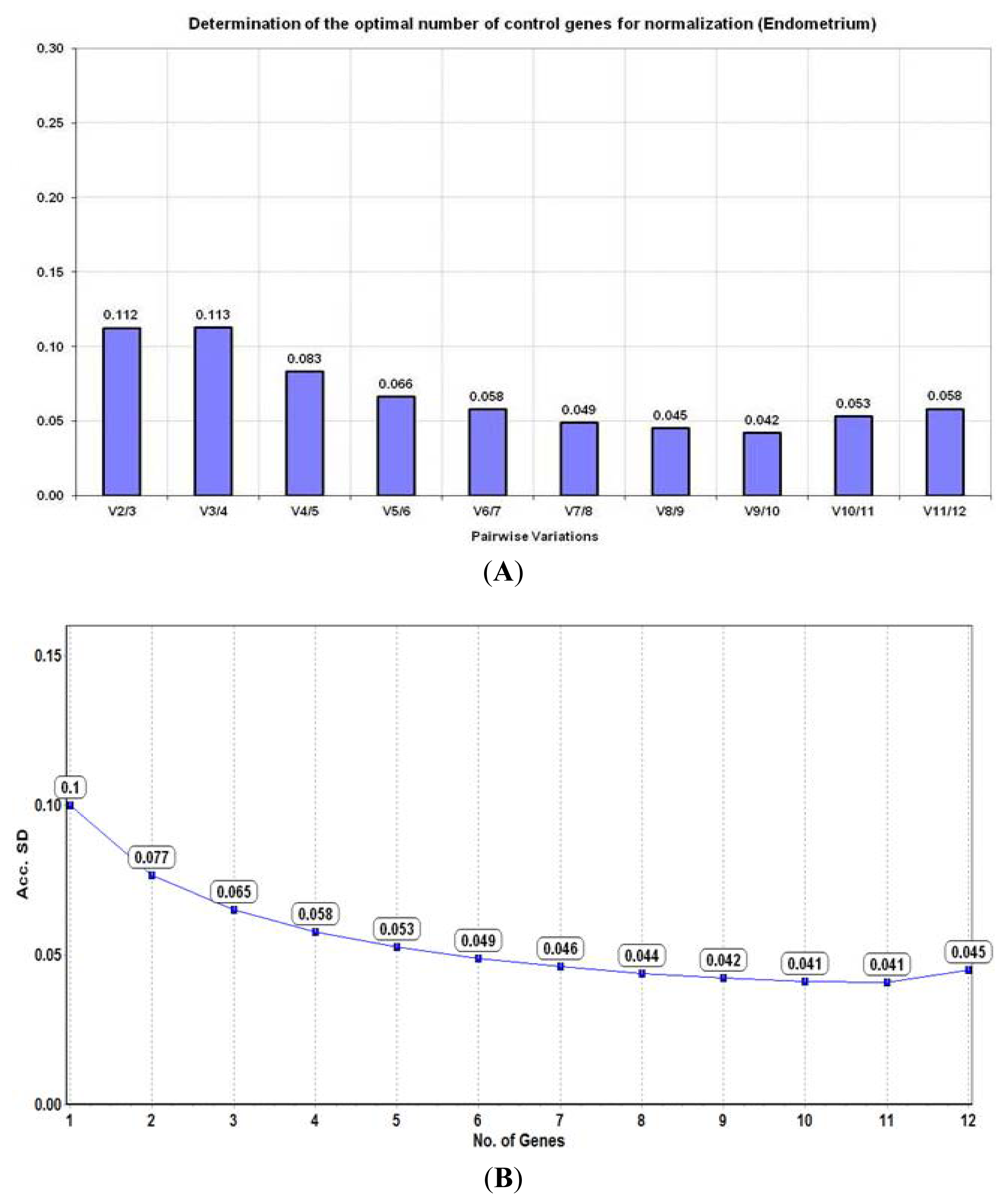
2.4. Analysis of Reference Gene Stability in Endometrium
2.4.1. geNorm Analysis
2.4.2. NormFinder Algorithm
2.5. The Effect of Choice of Using Different Reference Genes to Normalise Data for Genes of Interest
2.6. General Discussion of Results
3. Experimental Section
3.1. Ethics Statement
3.2. Subjects
3.3. Tissue Collection
3.4. Isolation of RNA and cDNA Synthesis
3.5. Quantitative Real-Time PCR
3.6. Software Determination of Reference Genes Stability
3.7. Gene Expression Stability Analysis
- The highest Cq value was subtracted from all other Cq values for each gene measured. This gave a delta Cq value (ΔCq) with the highest delta Cq value being 0. All other values were less than 0.
- To each ΔCq value, the equation (2−ΔCq) was applied. Hence all data were expressed relative to the expression of the least expressed gene. Relative expression values were exported to geNorm and NormFinder for analysis.
3.8. Statistical Analysis
4. Conclusions
Acknowledgments
- Conflicts of InterestThe authors declare no conflicts of interest.
References
- Farquhar, C.M. Ectopic pregnancy. Lancet 2005, 366, 583–591. [Google Scholar]
- Cantwell, R.; Clutton-Brock, T.; Cooper, G.; Dawson, A.; Drife, J.; Garrod, D.; Harper, A.; Hulbert, D.; Lucas, S.; McClure, J.; et al. Saving Mothers’ Lives: Reviewing maternal deaths to make motherhood safer: 2006–2008. The Eighth Report of the Confidential Enquiries into Maternal Deaths in the United Kingdom. BJOG 2011, 118, 1–203. [Google Scholar]
- Pisarska, M.D.; Carson, S.A. Incidence and risk factors for ectopic pregnancy. Clin. Obstet. Gynecol 1999, 42, 2–8. [Google Scholar]
- Gebeh, A.; Willets, J.; Marczylo, T.; Lam, P.; Taylor, A.; Konje, J. Molecular mechanisms of tubal pregnancy. Expert Rev. Obstet. Gynecol 2010, 5, 727–739. [Google Scholar]
- Radonic, A.; Thulke, S.; Mackay, I.M.; Landt, O.; Siegert, W.; Nitsche, A. Guideline to reference gene selection for quantitative real-time PCR. Biochem. Biophys. Res. Commun 2004, 313, 856–862. [Google Scholar]
- Huggett, J.; Dheda, K.; Bustin, S.; Zumla, A. Real-time RT-PCR normalisation; strategies and considerations. Genes Immun 2005, 6, 279–284. [Google Scholar]
- Bustin, S.A. Absolute quantification of mRNA using real-time reverse transcription polymerase chain reaction assays. J. Mol. Endocrinol 2000, 25, 169–193. [Google Scholar]
- Bustin, S.A. Quantification of mRNA using real-time reverse transcription PCR (RT-PCR): Trends and problems. J. Mol. Endocrinol 2002, 29, 23–39. [Google Scholar]
- Romanowski, T.; Markiewicz, A.; Bednarz, N.; Bielawski, K.P. Housekeeping genes as a reference in quantitative real-time RT-PCR. Postepy Hig. Med. Dosw 2007, 61, 500–510. [Google Scholar]
- Ginzinger, D.G. Gene quantification using real-time quantitative PCR: An emerging technology hits the mainstream. Exp. Hematol 2002, 30, 503–512. [Google Scholar]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE guidelines: Minimum information for publication of quantitative real-time PCR experiments. Clin. Chem 2009, 55, 611–622. [Google Scholar]
- Taylor, A.H.; Amoako, A.A.; Bambang, K.; Karasu, T.; Gebeh, A.; Lam, P.M.; Marzcylo, T.H.; Konje, J.C. Endocannabinoids and pregnancy. Clin. Chim. Acta 2010, 411, 921–930. [Google Scholar]
- Bambang, K.; Karasu, T.; Gebeh, A.; Taylor, A.; Marczylo, T.; Lam, P.; Willets, J.; Konje, J. From fertilisation to implantation in mammalian pregnancy-modulation of early human reproduction by the endocannabinoid system. Pharmaceuticals 2010, 3, 2910–2929. [Google Scholar]
- Feng, X.; Xiong, Y.; Qian, H.; Lei, M.; Xu, D.; Ren, Z. Selection of reference genes for gene expression studies in porcine skeletal muscle using SYBR green qPCR. J. Biotechnol 2010, 150, 288–293. [Google Scholar]
- Hoogewijs, D.; Houthoofd, K.; Matthijssens, F.; Vandesompele, J.; Vanfleteren, J.R. Selection and validation of a set of reliable reference genes for quantitative sod gene expression analysis in Caenorhabditis elegans. BMC Mol. Biol 2008, 9. [Google Scholar] [CrossRef]
- Penna, I.; Vella, S.; Gigoni, A.; Russo, C.; Cancedda, R.; Pagano, A. Selection of candidate housekeeping genes for normalization in human postmortem brain samples. Int. J. Mol. Sci 2011, 12, 5461–5470. [Google Scholar]
- Taylor, A.H.; Abbas, M.S.; Habiba, M.A.; Konje, J.C. Histomorphometric evaluation of cannabinoid receptor and anandamide modulating enzyme expression in the human endometrium through the menstrual cycle. Histochem. Cell Biol 2010, 133, 557–565. [Google Scholar]
- geNorm, version 3.5; Center for Medical Genetics; Ghent University: Ghent, Belgium, 2007.
- NormFinder, version 0.953; Aarhus University Hospital: Aarhus, Denmark, 2005.
- GenEx software, version 5.3.6.170; MultiD Analyses AB: Göteborg, Sweden, 2011.
- Vandesompele, J.; de Preter, K.; Pattyn, F.; Poppe, B.; van Roy, N.; de Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol 2002, 3, RESEARCH0034.1–0034.11. [Google Scholar]
- Andersen, C.L.; Jensen, J.L.; Orntoft, T.F. Normalization of real-time quantitative reverse transcription-PCR data: A model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets. Cancer Res 2004, 64, 5245–5250. [Google Scholar]
- Mehta, R.; Birerdinc, A.; Hossain, N.; Afendy, A.; Chandhoke, V.; Younossi, Z.; Baranova, A. Validation of endogenous reference genes for qRT-PCR analysis of human visceral adipose samples. BMC Mol. Biol 2010, 11, 39. [Google Scholar]
- GraphPad Prism, version 5.00; GraphPad Software: San Diego, CA, USA, 2009.







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Genbank Number | geNorm Ranking | M Value | NormFinder Ranking | Stability Value |
|---|---|---|---|---|---|
| EIF4A2 | NM_001967.3 | 1 | 0.41 | 6 | 0.119 |
| CYC1 | NM_001916.3 | 1 | 0.41 | 4 | 0.109 |
| UBC | NM_021009.5 | 3 | 0.55 | 1 | 0.098 |
| RN18S1 | NR_003286.2 | 4 | 0.59 | 8 | 0.137 |
| B2M | NM_004048.2 | 5 | 0.65 | 9 | 0.139 |
| GAPDH | NM_002046.3 | 6 | 0.70 | 5 | 0.115 |
| TOP1 | NM_003286.2 | 7 | 0.94 | 2 | 0.100 |
| ATP5B | NM_001686.3 | 8 | 1.10 | 7 | 0.122 |
| ACTB | NM_001101.3 | 9 | 1.28 | 12 | 0.154 |
| SDHA | NM_004168.2 | 10 | 1.48 | 11 | 0.152 |
| YWHAZ | NM_001135699.1 | 11 | 1.78 | 10 | 0.142 |
| RPL13A | NM_012423.2 | 12 | 1.98 | 3 | 0.103 |
| Best combination of genes | UBC, CYC1 & EIF4A2 | - | UBC & TOP1 | 0.064 |
| Gene Name | Genbank Number | geNorm Ranking | M Value | Normfinder Ranking | Stability Value |
|---|---|---|---|---|---|
| YWHAZ | NM_001135699.1 | 1 | 0.20 | 8 | 0.121 |
| UBC | NM_021009.5 | 1 | 0.20 | 3 | 0.088 |
| ATP5B | NM_001686.3 | 3 | 0.30 | 2 | 0.081 |
| SDHA | NM_004168.2 | 4 | 0.39 | 7 | 0.113 |
| RN18S1 | NR_003286.2 | 5 | 0.42 | 10 | 0.144 |
| EIF4A2 | NM_001967.3 | 6 | 0.44 | 11 | 0.148 |
| TOP1 | NM_003286.2 | 7 | 0.46 | 5 | 0.106 |
| CYC1 | NM_001916.3 | 8 | 0.47 | 1 | 0.074 |
| ACTB | NM_001101.3 | 9 | 0.48 | 9 | 0.125 |
| GAPDH | NM_002046.3 | 10 | 0.49 | 6 | 0.106 |
| RPL13A | NM_012423.2 | 11 | 0.52 | 12 | 0.148 |
| B2M | NM_004048.2 | 12 | 0.56 | 4 | 0.100 |
| Best combination of genes | UBC & YWHAZ | - | UBC & ATP5B | 0.056 |
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Gebeh, A.K.; Marczylo, E.L.; Amoako, A.A.; Willets, J.M.; Konje, J.C. Variation in Stability of Endogenous Reference Genes in Fallopian Tubes and Endometrium from Healthy and Ectopic Pregnant Women. Int. J. Mol. Sci. 2012, 13, 2810-2826. https://doi.org/10.3390/ijms13032810
Gebeh AK, Marczylo EL, Amoako AA, Willets JM, Konje JC. Variation in Stability of Endogenous Reference Genes in Fallopian Tubes and Endometrium from Healthy and Ectopic Pregnant Women. International Journal of Molecular Sciences. 2012; 13(3):2810-2826. https://doi.org/10.3390/ijms13032810
Chicago/Turabian StyleGebeh, Alpha K., Emma L. Marczylo, Akwasi A. Amoako, Jonathon M. Willets, and Justin C. Konje. 2012. "Variation in Stability of Endogenous Reference Genes in Fallopian Tubes and Endometrium from Healthy and Ectopic Pregnant Women" International Journal of Molecular Sciences 13, no. 3: 2810-2826. https://doi.org/10.3390/ijms13032810