Computational Analysis of Axonal Transport: A Novel Assessment of Neurotoxicity, Neuronal Development and Functions

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Physiological Functions of Axonal Transport
2.1. The Role of Axonal Transport in Neural Survival
2.2. The Role of Axonal Transport in Neural Network Formation
2.3. Axonal Transport and Neural Plasticity
2.4. Pathophysiology of Axonal Transport
2.5. Comparison of the Analytical Methods for Axonal Transport
2.6. Difficulties in Axonal Transport Analysis
- The gray scale of the vesicle is not similar and the images have a lot of random noise, which made vesicle detection difficult using simple image processing technologies.
- The vesicles move bi-directionally along the axon with different velocities and/or opposite directions.
- Since the vesicles move in close proximity in a narrow passageway, the vesicles may appear as a single particle on an image frame and then as several particles on the next.
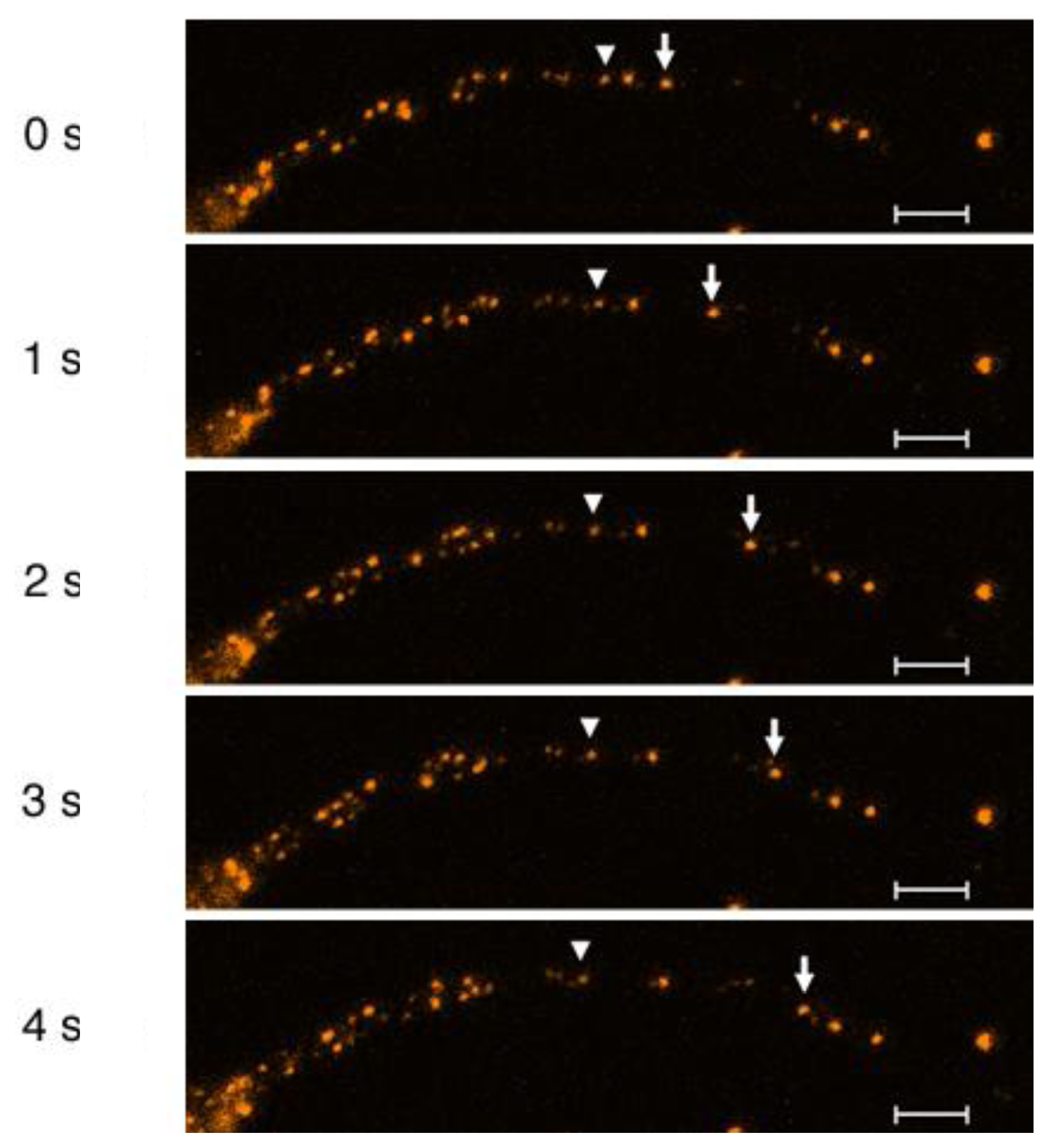
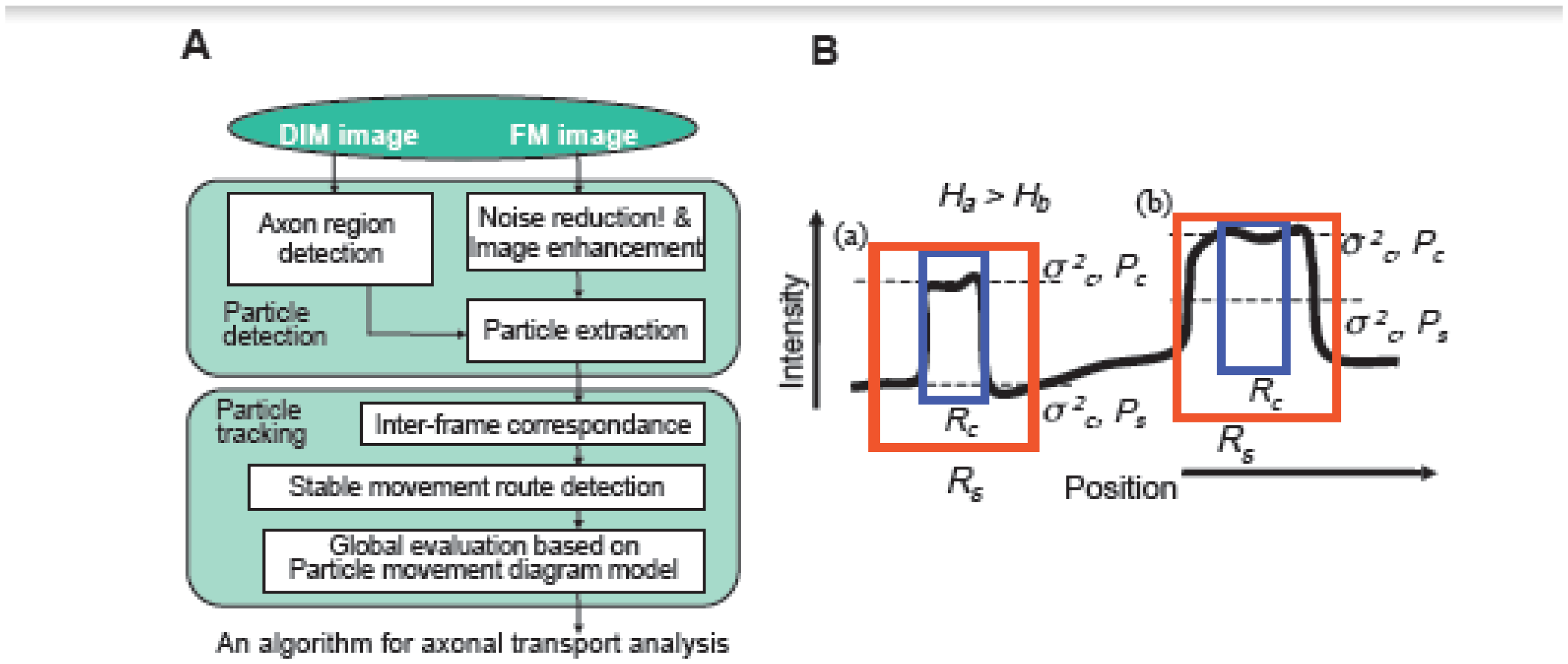
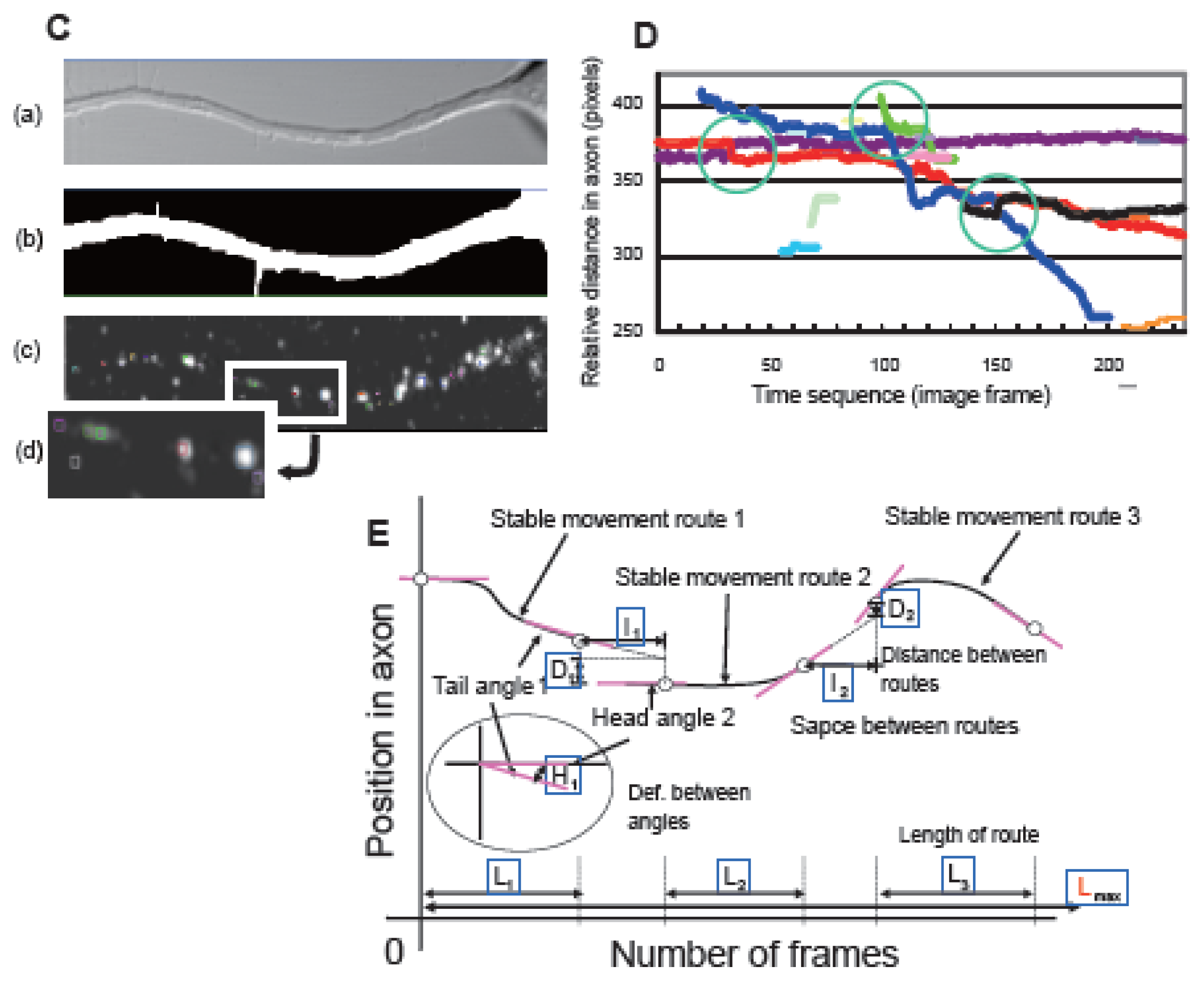
2.7. Computer-Based Particle Detection and Tracking Methods
2.8. Velocity Distribution of Particle Movements
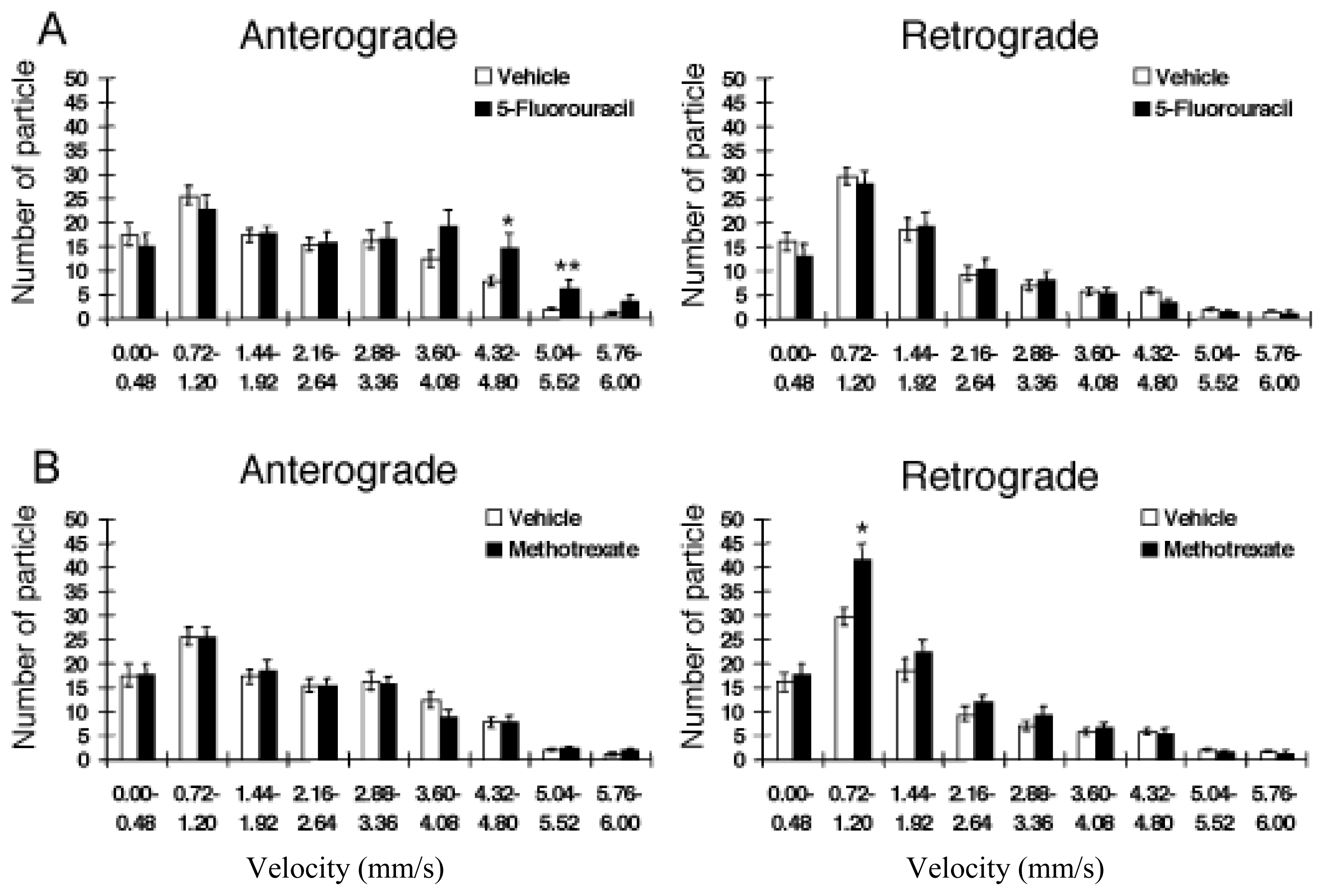
2.9. An Example of Computational Analysis of Axonal Transport: A Neurotoxicity Test
3. Experimental Section
4. Conclusions
Acknowledgements
- Conflict of InterestThe authors declare no conflict of interest.
References
- Hirokawa, N.; Takemura, R. Molecular motors and mechanisms of directional transport in neurons. Nat. Rev. Neurosci 2005, 6, 201–214. [Google Scholar]
- Hirokawa, N.; Noda, Y. Intracellular transport and kinesin superfamily proteins, KIFs: Structure, function, and dynamics. Physiol. Rev 2008, 88, 1089–1118. [Google Scholar]
- Vale, R.D.; Schnapp, B.J.; Mitchison, T.; Steuer, E.; Reese, T.S.; Sheetz, M.P. Different axoplasmic proteins generate movement in opposite directions along microtubules in vitro. Cell 1985, 43, 623–632. [Google Scholar]
- Goshima, Y.; Usui, H.; Shiozawa, T.; Hida, T.; Kuraoka, S.; Takeshita, S.; Yamashita, N.; Ichikawa, Y.; Kamiya, Y.; Gotoh, T. Computational analysis of the effects of antineoplastic agents on axonal transport. J. Pharmacol. Sci 2010, 114, 168–179. [Google Scholar]
- de Vos, K.J.; Grierson, A.J.; Ackerley, S.; Miller, C.C. Role of axonal transport in neurodegenerative diseases. Annu. Rev. Neurosci 2008, 31, 151–173. [Google Scholar]
- Malik, B.; Stillman, M. Chemotherapy-induced peripheral neuropathy. Curr. Neurol. Neurosci. Rep 2008, 8, 56–65. [Google Scholar]
- Nakata, T.; Terada, S.; Hirokawa, N. Visualization of the dynamics of synaptic vesicle and plasma membrane proteins in living axons. J. Cell Biol 1998, 140, 659–674. [Google Scholar]
- Perlson, E.; Maday, S.; Fu, M.M.; Moughamian, A.J.; Holzbaur, E.L. Retrograde axonal transport: Pathways to cell death? Trends Neurosci 2010, 33, 335–344. [Google Scholar]
- Li, C.; Sasaki, Y.; Takei, K.; Yamamoto, H.; Shouji, M.; Sugiyama, Y.; Kawakami, T.; Nakamura, F.; Yagi, T.; Ohshima, T.; et al. Correlation between semaphorin3A-induced facilitation of axonal transport and local activation of a translation initiation factor eukaryotic translation initiation factor 4E. J. Neurosci 2004, 24, 6161–6170. [Google Scholar]
- Goshima, Y.; Kawakami, T.; Hori, H.; Sugiyama, Y.; Takasawa, S.; Hashimoto, Y.; Kagoshima-Maezono, M.; Takenaka, T.; Misu, Y.; Strittmatter, S.M. A novel action of collapsin: Collapsin-1 increases antero- and retrograde axoplasmic transport independently of growth cone collapse. J. Neurobiol 1997, 33, 316–328. [Google Scholar]
- Goshima, Y.; Hori, H.; Sasaki, Y.; Yang, T.; Kagoshima-Maezono, M.; Li, C.; Takenaka, T.; Nakamura, F.; Takahashi, T.; Strittmatter, S.M.; et al. Growth cone neuropilin-1 mediates collapsin-1/Sema III facilitation of antero- and retrograde axoplasmic transport. J. Neurobiol 1999, 39, 579–589. [Google Scholar]
- Yamashita, N.; Chen, S.; Goshima, Y. Semaphorin3A drives AMPA receptor subunit GluR2 to dendrites by a retrograde transport of PlexinA4 along axon. Neurosci. Res 2011, 71S, e56. [Google Scholar]
- LaMonte, B.H.; Wallace, K.E.; Holloway, B.A.; Shelly, S.S.; Ascano, J.; Tokito, M.; van Winkle, T.; Howland, D.S.; Holzbaur, E.L. Disruption of dynein/dynactin inhibits axonal transport in motor neurons causing late-onset progressive degeneration. Neuron 2002, 34, 715–727. [Google Scholar]
- Hafezparast, M.; Klocke, R.; Ruhrberg, C.; Marquardt, A.; Ahmad-Annuar, A.; Bowen, S.; Lalli, G.; Witherden, A.S.; Hummerich, H.; Nicholson, S.; et al. Mutations in dynein link motor neuron degeneration to defects in retrograde transport. Science 2003, 300, 808–812. [Google Scholar]
- Heerssen, H.M.; Pazyra, M.F.; Segal, R.A. Dynein motors transport activated Trks to promote survival of target-dependent neurons. Nat. Neurosci 2004, 7, 596–604. [Google Scholar]
- Dechant, G.; Barde, Y.A. The neurotrophin receptor p75(NTR): Novel functions and implications for diseases of the nervous system. Nat. Neurosci 2002, 5, 1131–1136. [Google Scholar]
- Ye, H.; Kuruvilla, R.; Zweifel, L.S.; Ginty, D.D. Evidence in support of signaling endosome-based retrograde survival of sympathetic neurons. Neuron 2003, 39, 57–68. [Google Scholar]
- Wu, C.; Cui, B.; He, L.; Chen, L.; Mobley, W.C. The coming of age of axonal neurotrophin signaling endosomes. J. Proteomics 2009, 72, 46–55. [Google Scholar]
- McIntire, S.L.; Jorgensen, E.; Horvitz, H.R. The GABAergic nervous system of Caenorhabditis elegans. Nature 1993, 364, 334–337. [Google Scholar]
- Ogura, K.; Goshima, Y. The autophagy-related kinase UNC-51 and its binding partner UNC-14 regulate the subcellular localization of the Netrin receptor UNC-5 in Caenorhabditis elegans. Development 2006, 133, 3441–3450. [Google Scholar]
- Asakura, T.; Ogura, K.; Goshima, Y. UNC-6 expression by the vulval precursor cells of Caenorhabditis elegans is required for the complex axon guidance of the HSN neurons. Dev. Biol 2007, 304, 800–810. [Google Scholar]
- Asakura, T.; Waga, N.; Ogura, K.; Goshima, Y. Genes required for cellular UNC-6/netrin localization in Caenorhabditis elegans. Genetics 2010, 185, 573–585. [Google Scholar]
- Jia, Y.; Gall, C.M.; Lynch, G. Presynaptic BDNF promotes postsynaptic long-term potentiation in the dorsal striatum. J. Neurosci 2010, 30, 14440–14445. [Google Scholar]
- Ligon, L.A.; Steward, O. Role of microtubules and actin filaments in the movement of mitochondria in the axons and dendrites of cultured hippocampal neurons. J. Comp. Neurol 2000, 427, 351–361. [Google Scholar]
- Perlson, E.; Jeong, G.B.; Ross, J.L.; Dixit, R.; Wallace, K.E.; Kalb, R.G.; Holzbaur, E.L. A switch in retrograde signaling from survival to stress in rapid-onset neurodegeneration. J. Neurosci 2009, 29, 9903–9917. [Google Scholar]
- Her, L.S.; Goldstein, L.S. Enhanced sensitivity of striatal neurons to axonal transport defects induced by mutant huntingtin. J. Neurosci 2008, 28, 13662–13672. [Google Scholar]
- Gunawardena, S.; Her, L.S.; Brusch, R.G.; Laymon, R.A.; Niesman, I.R.; Gordesky-Gold, B.; Sintasath, L.; Bonini, N.M.; Goldstein, L.S. Disruption of axonal transport by loss of huntingtin or expression of pathogenic polyQ proteins in Drosophila. Neuron 2003, 40, 25–40. [Google Scholar]
- Stokin, G.B.; Lillo, C.; Falzone, T.L.; Brusch, R.G.; Rockenstein, E.; Mount, S.L.; Raman, R.; Davies, P.; Masliah, E.; Williams, D.S.; et al. Axonopathy and transport deficits early in the pathogenesis of Alzheimer’s disease. Science 2005, 307, 1282–1288. [Google Scholar]
- Mukherjee, A.; Jenkins, B.; Fang, C.; Radke, R.J.; Banker, G.; Roysam, B. Automated kymograph analysis for profiling axonal transport of secretory granules. Med. Image Anal 2011, 15, 354–367. [Google Scholar]
- Broeke, J.H.; Ge, H.; Dijkstra, I.M.; Cemgil, A.T.; Riedl, J.A.; Cornelisse, L.N.; Toonen, R.F.; Verhage, M.; Fitzgerald, W.J. Automated quantification of cellular traffic in living cells. J. Neurosci. Meth 2009, 178, 378–384. [Google Scholar]
- Welzel, O.; Knorr, J.; Stroebel, A.M.; Kornhuber, J.; Groemer, T.W. A fast and robust method for automated analysis of axonal transport. Eur. Biophys. J 2011, 40, 1061–1069. [Google Scholar]
- Yamaguchi, O.; Fukui, K.; Maeda, K. Face Recognition Using Temporal Image Sequence. Proceedings of the 3rd IEEE International Conference on Automatic Face and Gesture Recognition, Nara, Japan, 14–16 April 1998; IEEE Computer Society: Washington, DC, USA, 1998; pp. 318–323. [Google Scholar]
- Schmitt, L. Theory of genetic algorithm. Theor. Comput. Sci 2001, 259, 1–61. [Google Scholar]
- Hough, P.V.C. Method and Means for Recognizing Complex Patterns. US Patent 3,969,654, 18 December 1962. [Google Scholar]
- Araki, Y.; Kawano, T.; Taru, H.; Saito, Y.; Wada, S.; Miyamoto, K.; Kobayashi, H.; Ishikawa, H.O.; Ohsugi, Y.; Yamamoto, T.; et al. The novel cargo Alcadein induces vesicle association of kinesin-1 motor components and activates axonal transport. EMBO J 2007, 26, 1475–1486. [Google Scholar]
- Kanai, A.; Hiruma, H.; Katakura, T.; Sase, S.; Kawakami, T.; Hoka, S. Low-concentration lidocaine rapidly inhibits axonal transport in cultured mouse dorsal root ganglion neurons. Anesthesiology 2001, 95, 675–680. [Google Scholar]
- Scripture, C.D.; Figg, W.D.; Sparreboom, A. Peripheral neuropathy induced by paclitaxel: Recent insights and future perspectives. Curr. Neuropharmacol 2006, 4, 165–172. [Google Scholar]
- Kaley, T.J.; Deangelis, L.M. Therapy of chemotherapy-induced peripheral neuropathy. Br. J. Haematol 2009, 145, 3–14. [Google Scholar]
- Price, R.D.; Milne, S.A.; Sharkey, J.; Matsuoka, N. Advances in small molecules promoting neurotrophic function. Pharmacol. Ther 2007, 115, 292–306. [Google Scholar]
- Wolf, S.; Barton, D.; Kottschade, L.; Grothey, A.; Loprinzi, C. Chemotherapy-induced peripheral neuropathy: Prevention and treatment strategies. Eur. J. Cancer 2008, 44, 1507–1515. [Google Scholar]
- Goshima, Y.; Nakamura, F.; Strittmatter, P.; Strittmatter, S.M. Collapsin-induced growth cone collapse mediated by an intracellular protein related to UNC-33. Nature 1995, 376, 509–514. [Google Scholar]
- Lee, G.; Papapetrou, E.P.; Kim, H.; Chambers, S.M.; Tomishima, M.J.; Fasano, C.A.; Ganat, Y.M.; Menon, J.; Shimizu, F.; Viale, A.; et al. Modelling pathogenesis and treatment of familial dysautonomia using patient-specific iPSCs. Nature 2009, 461, 402–406. [Google Scholar]






© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Goshima, Y.; Hida, T.; Gotoh, T. Computational Analysis of Axonal Transport: A Novel Assessment of Neurotoxicity, Neuronal Development and Functions. Int. J. Mol. Sci. 2012, 13, 3414-3430. https://doi.org/10.3390/ijms13033414
Goshima Y, Hida T, Gotoh T. Computational Analysis of Axonal Transport: A Novel Assessment of Neurotoxicity, Neuronal Development and Functions. International Journal of Molecular Sciences. 2012; 13(3):3414-3430. https://doi.org/10.3390/ijms13033414
Chicago/Turabian StyleGoshima, Yoshio, Tomonobu Hida, and Toshiyuki Gotoh. 2012. "Computational Analysis of Axonal Transport: A Novel Assessment of Neurotoxicity, Neuronal Development and Functions" International Journal of Molecular Sciences 13, no. 3: 3414-3430. https://doi.org/10.3390/ijms13033414