Antihyperglycemic and Antioxidative Effects of Hydroxyethyl Methylcellulose (HEMC) and Hydroxypropyl Methylcellulose (HPMC) in Mice Fed with a High Fat Diet


Abstract
:1. Introduction
2. Results and Discussion
2.1. Body Weight Gain
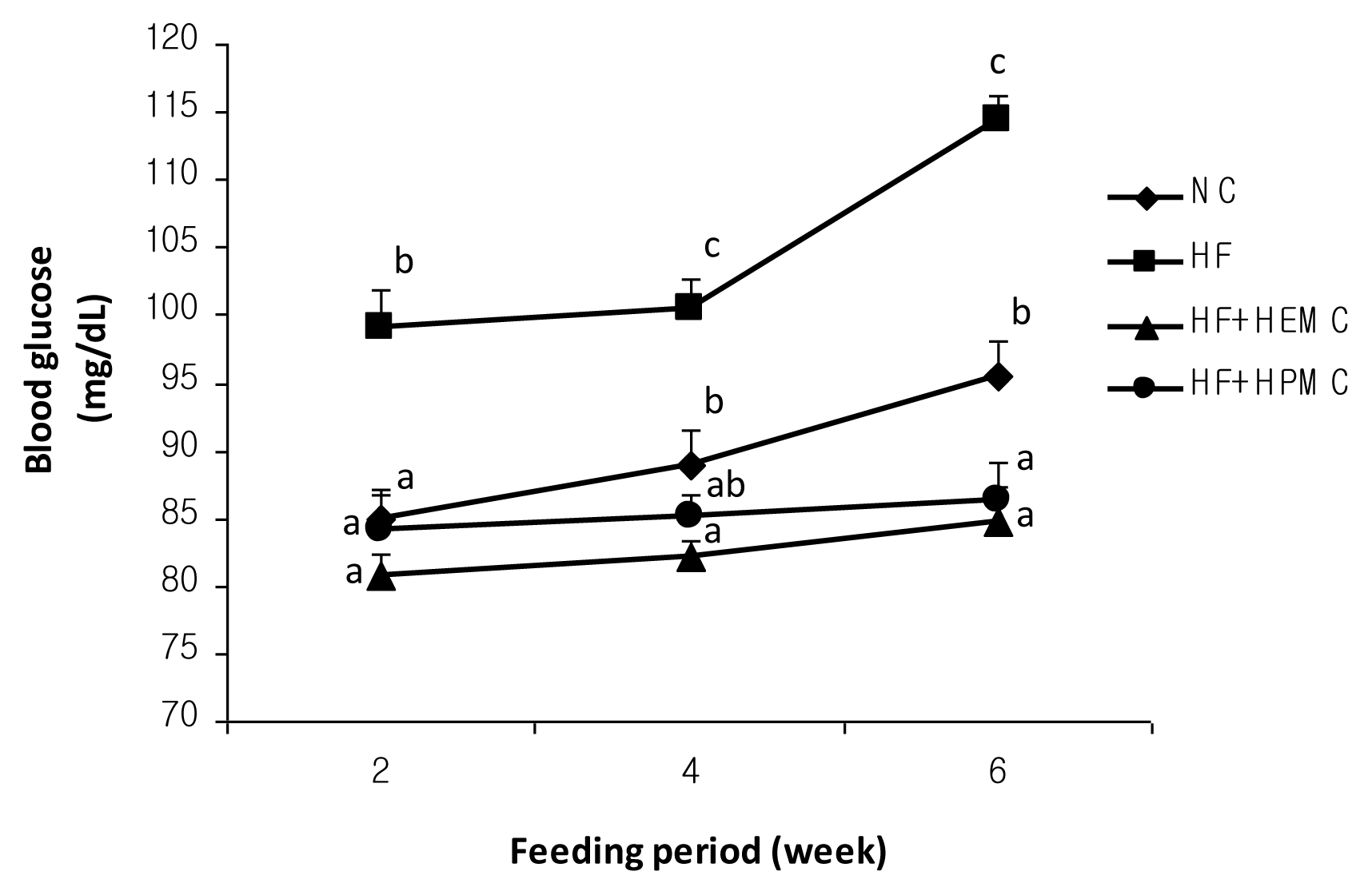
2.2. Blood Glucose Level
2.3. Glycogen and Insulin Concentrations
2.4. Plasma and Erythrocyte Lipid Peroxides
2.5. Hepatic Glucose-Regulating Enzyme Activities
2.6. Antioxidant Enzyme Activities
3. Experimental Section
3.1. Materials
3.2. Animals and Diet
3.3. Determination of Blood Glucose Level
3.4. Measurement of Glycogen and Insulin Levels
3.5. Determination of Lipid Peroxidation
3.6. Determination of Hepatic Glucose-Regulating Enzyme Activities
3.7. Measurement of Antioxidant Enzyme Activities
3.8. Statistical Analysis
4. Conclusions
Acknowledgments
References
- Burdock, G.A. Safety assessment of hydroxypropyl methylcellulose as a food ingredient. Food Chem. Toxicol 2007, 45, 2341–2351. [Google Scholar]
- Maki, K.C.; Carson, M.L.; Anderson, W.H.K.; Geohas, J.; Reeves, M.S.; Farmer, M.V.; Turowski, M.; Miller, M.; Kaden, V.N.; Dicklin, M.R.; et al. Lipid-altering effects of different formulations of hydroxymethylcellulose. J. Clin. Lipidol 2009, 3, 159–166. [Google Scholar]
- Maki, K.C.; Davidson, M.H.; Malik, K.C.; Albrecht, H.H.; O’Mullane, J.; Daggy, B.P. Cholesterol lowering with high-viscosity hydroxypropylmethylcellulose. Am. J. Cardiol 1999, 84, 1198–1203. [Google Scholar]
- Reppas, C.; Swidan, S.Z.; Tobey, S.W.; Turowski, M.; Dressman, J.B. Hydroxypropylmethylcellulose significantly lowers blood cholesterol in mildly hypercholesterolemic human subjects. Eur. J. Clin. Nutr 2009, 63, 71–77. [Google Scholar]
- Swidan, S.Z.; Reppas, C.; Barnett, J.L.; Grenwood, D.E.; Tallman, A.M.; Tobey, S.W.; Dressman, J.B. Ability of two comestible formulations of hydroxypropylmethylcellulose to lower serum cholesterol concentrations. Eur. J. Pharm. Sci 1996, 4, 239–245. [Google Scholar]
- Hung, S.C.; Anderson, W.H.K.; Albers, D.R.; Langhorst, M.L.; Young, S.A. Effect of hydroxypropyl methylcellulose on obesity and glucose metabolism in a diet-induced obesity mouse model. J. Diabetes 2011, 3, 158–167. [Google Scholar]
- Maki, K.C.; Carson, M.L.; Miller, M.P.; Turowski, M.; Bell, M.; Wilder, D.M.; Rains, T.M.; Reeves, M.S. Hydroxypropylmethylcellulose and methylcellulose consumption reduce postprandial insulinemia in overweight and obese men and women. J. Nutr 2008, 138, 292–296. [Google Scholar]
- Bray, G.A.; Paeratakul, S.; Popkin, B.M. Dietary fat and obesity: A review of animal, clinical and epidemiological studies. Physiol. Behav 2004, 83, 549–555. [Google Scholar]
- Park, J.; Rho, H.K.; Kim, K.H.; Choi, S.S.; Lee, Y.S.; Kim, J.B. Overexpression of glucose-6-phosphate dehydrogenase is associated with lipid dysregulation and insulin resistance in obesity. Mol. Cell. Biol 2005, 25, 5146–5157. [Google Scholar]
- Lichtenstein, A.H.; Schwab, U.S. Relationship of dietary fat to glucose metabolism. Atherosclerosis 2000, 150, 227–243. [Google Scholar]
- Sharma, N.; Garg, V.; Paul, A. Antihyperglycemic, antihyperlipidemic and antioxidative potential of Prosopis cineraria bark. Indian J. Clin. Biochem 2010, 25, 193–200. [Google Scholar]
- Ban, S.J.; Rico, C.W.; Um, I.C.; Kang, M.Y. Comparative evaluation of the hypolipidemic effects of hydroxyethyl methylcellulose (HEMC) and hydroxypropyl methylcellulose (HPMC) in high fat-fed mice. Food Chem. Toxicol 2012, 50, 130–134. [Google Scholar]
- Amrani, A.; Durant, S.; Throsby, M.; Coulaud, J.; Dardenne, M.; Homo-Delarche, F. Glucose homeostasis in the nonobese diabetic mouse at the prediabetic stage. Endoctrinology 1998, 139, 1115–1124. [Google Scholar]
- Hung, S.C.; Bartley, G.; Young, S.A.; Albers, D.R.; Dielman, D.R.; Anderson, W.H.K.; Yokohama, W. Dietary fiber improves lipid homeostasis and modulates adipocytokines in hamsters. J. Diabetes 2009, 1, 194–206. [Google Scholar]
- Reppas, C.; Adair, C.H.; Barnett, J.L.; Berardi, R.R.; DuRoss, D.; Swidan, S.Z.; Thill, P.F.; Tobey, S.W.; Dressman, J.B. High viscosity hydroxypropylmethylcellulose reduces postprandial blood glucose concentrations in NIDDM patients. Diabetes Res. Clin. Pract 1993, 22, 61–69. [Google Scholar]
- Reppas, C.; Greenwood, D.E.; Dressman, J.B. Longitudinal versus radial effects of hydroxypropyl methylcellulose on gastrointestinal glucose absorption in dogs. Eur. J. Pharmacol. Sci 1999, 8, 211–219. [Google Scholar]
- Topping, D.L.; Oakenfull, D.; Trimble, R.P.; Illman, R.J. A viscous fibre (methylcellulose) lowers blood glucose and plasma triacylglycerols and increases liver glycogen independently of volatile fatty acid production in the rat. Br. J. Nutr 1988, 59, 21–30. [Google Scholar]
- Maki, K.C.; Carson, M.L.; Miller, M.P.; Turowski, M.; Bell, M.; Wilder, D.M.; Reeves, M.S. High-viscosity hydroxypropylmethylcellulose blunts postprandial glucose and insulin responses. Diabetes Care 2007, 30, 1039–1043. [Google Scholar]
- Ibrahim, W.; Lee, U.S.; Yeh, C.C.; Szabo, J.; Bruckner, G.; Chow, C.K. Oxidative stress and antioxidant status in mouse liver: Effects of dietary lipid, vitamin E and iron. J. Nutr 1997, 127, 1401–1406. [Google Scholar]
- Sanchez, D.; Quiñones, M.; Moulay, L.; Muguerza, B.; Miguel, M.; Aleixandre, A. Soluble fiber-enriched diets improve inflammation and oxidative stress biomarkers in Zucker fatty rats. Pharmacol. Res 2011, 64, 31–35. [Google Scholar]
- Thampi, B.S.; Manoj, G.; Leelamma, S.; Menon, V.P. Dietary fiber and lipid peroxidation: Effect of dietary fiber on levels of lipids and lipid peroxides in high fat diet. Indian J. Exp. Biol 1991, 29, 563–567. [Google Scholar]
- Coope, G.J.; Atkinson, A.M.; Allott, C.; McKerrecher, D.; Johnstone, C.; Pike, K.G.; Holme, P.C.; Vertigan, H.; Gill, D.; Coghlan, M.P.; et al. Predictive blood glucose lowering efficacy by glucokinase activators in high fat fed female zucker rats. Br. J. Pharmacol 2006, 149, 328–335. [Google Scholar]
- Friedman, J.E.; Sun, Y.; Ishizuka, T.; Farrell, C.J.; McCormack, S.E.; Herron, L.M.; Hakimi, P.; Lechner, P.; Yun, J.S. Phosphoenolpyruvate carboxykinase (GTP) gene transcription and hyperglycemia are regulated by glucocorticoids in genetically obese db/db transgenic mice. J. Biol. Chem 1997, 272, 31475–31481. [Google Scholar]
- Devi, G.S.; Prasad, M.H.; Saraswathi, I.; Raghu, D.; Rao, D.N.; Reddy, P.P. Free radicals antioxidant enzymes and lipid peroxidation in different types of leukemia. Clin. Chim. Acta 2000, 293, 53–62. [Google Scholar]
- Reiter, R.J.; Tan, D.; Burkhardt, S. Reactive oxygen and nitrogen species and cellular and organismal decline:amelioration with melatonin. Mech. Aging Dev 2002, 123, 1007–1019. [Google Scholar]
- Ng, C.J.; Shih, D.M.; Hama, S.Y.; Villa, N.; Navab, M.; Reddy, S.T. The paraoxonase gene family and atherosclerosis. Free Radic. Biol. Med 2005, 38, 153–163. [Google Scholar]
- Mullineaux, P.M.; Creissen, G.P. Glutathione Reductase: Regulation and Role in Oxidative Stress. In Oxidative Stress and the Molecular Biology of Antioxidant Defenses; Scandalios, J.G., Ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 1997; pp. 667–713. [Google Scholar]
- May, S.; de Haen, C. The insulin-like effect of hydrogen peroxide on pathways of lipid synthesis in rat adipocytes. J. Biol. Chem 1979, 254, 9017–9021. [Google Scholar]
- American Institute of Nutrition. Report of ad hoc committee on standards for nutritional studies. J. Nutr. 1980, 110, 1717–1726.
- Seifter, S.; Dayton, S.; Navic, B.; Muntwyler, E. The estimation of glycogen with the anthrone reagent. Arch. Biochem 1950, 25, 191–200. [Google Scholar]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem 1979, 95, 351–358. [Google Scholar]
- Hulcher, F.H.; Oleson, W.H. Simplified spectrophotometric assay for microsomal 3-hydroxy-3-methylglutaryl CoA reductase by measurement of coenzyme A. J. Lipid Res 1973, 14, 625–631. [Google Scholar]
- Davidson, A.L.; Arion, W.J. Factors underlying significant underestimations of glucokinase activity in crude liver extracts: Physiological implications of higher cellular activity. Arch. Biochem. Biophys 1987, 253, 156–167. [Google Scholar]
- Alegre, M.; Ciudad, C.J.; Fillat, C.; Guinovart, J.J. Determination of glucose-6-phosphatase activity using the glucose dehydrogenase-coupled reaction. Anal. Biochem 1988, 173, 185–189. [Google Scholar]
- Bentle, L.A.; Lardy, H.A. Interaction of anions and divalent metal ions with phosphoenolpyruvate carboxykinase. J. Biol. Chem 1976, 251, 2916–2921. [Google Scholar]
- Marklund, S.; Marklund, G. Involvement of the superoxide anion radical in the autoxidation of pyrogallol and convenient assay for superoxide dismutase. Eur. J. Biochem 1974, 47, 469–474. [Google Scholar]
- Bradford, M.M. A rapid sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem 1976, 72, 248–254. [Google Scholar]
- Aebi, H. Catalase. In Method of Enzymatic Analysis; Bergmeyer, H.U., Ed.; Academic Press: New York NY, USA, 1974; Volume 2, pp. 673–684. [Google Scholar]
- Paglia, E.D.; Valentine, W.N. Studies on quantitative and qualitative characterization of erythrocyte glutathione peroxidase. J. Lab. Clin. Med 1967, 70, 158–169. [Google Scholar]
- Mize, C.E.; Langdon, R.G. Hepatic glutathione reductase, purification and general kinetic properties. J. Biol. Chem 1952, 237, 1589–1595. [Google Scholar]
- Mackness, M.I.; Arrol, S.; Durrington, P.N. Paraoxonase prevents accumulation of lipoperoxides in low-density lipoprotein. FEBS Lett 1991, 286, 152–154. [Google Scholar]


{kind=link}
| NC | HF | HF+HEMC | HF+HPMC | |
|---|---|---|---|---|
| Body weight gain (g) | 10.47 ± 0.72 a | 19.76 ± 0.36 d | 13.21 ± 0.83 b | 16.80 ± 0.61 c |
| Total feed intake (g) | 158.27 ± 3.12 a | 156.31 ± 2.03 a | 172.04 ± 4.63 b | 158.86 ± 2.99 a |
| Adipose tissue weight (g) | 7.62 ± 0.30 a | 12.02 ± 0.29 c | 10.30 ± 0.47 b | 10.27 ± 0.63 b |
| Dietary group | Glycogen (mg/g liver) | Insulin (ng/mL) |
|---|---|---|
| NC | 4.87 ± 0.56 a | 4.83 ± 0.30 a |
| HF | 4.28 ± 0.44 a | 9.96 ± 0.42 c |
| HF+HEMC | 5.96 ± 0.94 ab | 6.40 ± 0.32 b |
| HF+HPMC | 6.76 ± 0.23 b | 6.82 ± 0.30 b |
| Dietary group | Plasma TBARS (nmol/mL) | Erythrocyte TBARS (nmol/g Hb) |
|---|---|---|
| NC | 13.76 ± 0.48 c | 4.95 ± 0.31 b |
| HF | 13.41 ± 0.34 bc | 6.59 ± 0.68 c |
| HF+HEMC | 10.86 ± 0.30 a | 3.57 ± 0.02 a |
| HF+HPMC | 12.82 ± 0.16 b | 3.46 ± 0.19 a |
| Enzyme activity (nmol/min/mg protein) | |||
|---|---|---|---|
| Dietary group | GK | G6pase | PEPCK |
| NC | 2.09 ± 0.14 a | 162.00 ± 7.33 a | 3.16 ± 0.31 b |
| HF | 2.13 ± 0.06 a | 287.00 ± 13.00 c | 3.49 ± 0.14 b |
| HF+HEMC | 3.64 ± 0.07 b | 214.44 ± 7.39 b | 2.06 ± 0.51 a |
| HF+HPMC | 3.97 ± 0.14 c | 218.55 ± 8.53 b | 2.65 ± 0.23 ab |
| NC | HF | HF+HEMC | HF+HPMC | |
|---|---|---|---|---|
| Liver | ||||
| SOD (unit/mg protein) | 1.56 ± 0.14 a | 1.32 ± 0.12 a | 2.59 ± 0.15 b | 2.31 ± 0.25 b |
| GSH-Px (nmol/min/mg protein) | 4.03 ± 0.13 c | 3.75 ± 0.05 b | 3.14 ± 0.11 a | 3.30 ± 0.08 a |
| CAT (μmol/min/mg protein) | 1.17 ± 0.03 a | 1.14 ± 0.01 a | 1.46 ± 0.02 b | 1.49 ± 0.01 b |
| GR (nmol/min/mg protein) | 16.13 ± 0.67 b | 12.82 ± 1.00 a | 14.57 ± 0.99 ab | 14.74 ± 0.79 ab |
| PON (nmol/min/mg protein) | 2.48 ± 0.30 a | 2.26 ± 0.14 a | 3.85 ± 0.33 b | 3.86 ± 0.58 b |
| Erythrocyte | ||||
| SOD (unit/mg Hb) | 4.87 ± 0.33 a | 4.35 ± 0.13 a | 7.85 ± 1.02 b | 8.25 ± 1.00 b |
| GSH-Px (nmol/min/g Hb) | 2.15 ± 0.15 b | 1.62 ± 0.05 a | 1.97 ± 0.23 ab | 1.70 ± 0.05 a |
| CAT (μmol/min/g Hb) | 0.37 ± 0.01 a | 0.33 ± 0.01 a | 0.48 ± 0.04 b | 0.51 ± 0.02 b |
| GR (μmol/min/g Hb) | 0.27 ± 0.02 a | 0.27 ± 0.01 a | 0.82 ± 0.16 b | 0.73 ± 0.17 b |
| Sample | Viscosity (cps) | Degree of substitution (%) | ||
|---|---|---|---|---|
| Methyl group | Ethyl group | Propyl group | ||
| HEMC | 53900 | 28.30 | 8.34 | - |
| HPMC | 49000 | 22.21 | - | 8.68 |
| Component | Dietary group | |||
|---|---|---|---|---|
| NC | HF | HF+HEMC | HF+HPMC | |
| Casein | 20.0 | 20.0 | 20.0 | 20.0 |
| DL-Methionine | 0.3 | 0.3 | 0.3 | 0.3 |
| Sucrose | 50.0 | 50.0 | 48.0 | 48.0 |
| Cellulose | 5.0 | 5.0 | 5.0 | 5.0 |
| Corn oil | 5.0 | 3.0 | 3.0 | 3.0 |
| Choline bitartrate | 0.2 | 0.2 | 0.2 | 0.2 |
| Mineral mixture a | 3.5 | 3.5 | 3.5 | 3.5 |
| Vitamin mixture b | 1.0 | 1.0 | 1.0 | 1.0 |
| Corn starch | 15.0 | - | - | - |
| Lard | - | 17.0 | 17.0 | 17.0 |
| HEMC | - | - | 2.0 | - |
| HPMC | - | - | - | 2.0 |
| Total | 100.0 | 100.0 | 100.0 | 100.0 |
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ban, S.J.; Rico, C.W.; Um, I.C.; Kang, M.Y. Antihyperglycemic and Antioxidative Effects of Hydroxyethyl Methylcellulose (HEMC) and Hydroxypropyl Methylcellulose (HPMC) in Mice Fed with a High Fat Diet. Int. J. Mol. Sci. 2012, 13, 3738-3750. https://doi.org/10.3390/ijms13033738
Ban SJ, Rico CW, Um IC, Kang MY. Antihyperglycemic and Antioxidative Effects of Hydroxyethyl Methylcellulose (HEMC) and Hydroxypropyl Methylcellulose (HPMC) in Mice Fed with a High Fat Diet. International Journal of Molecular Sciences. 2012; 13(3):3738-3750. https://doi.org/10.3390/ijms13033738
Chicago/Turabian StyleBan, Su Jeong, Catherine W. Rico, In Chul Um, and Mi Young Kang. 2012. "Antihyperglycemic and Antioxidative Effects of Hydroxyethyl Methylcellulose (HEMC) and Hydroxypropyl Methylcellulose (HPMC) in Mice Fed with a High Fat Diet" International Journal of Molecular Sciences 13, no. 3: 3738-3750. https://doi.org/10.3390/ijms13033738