The Effect of Osmolytes on Protein Fibrillation

Abstract
:1. Introduction
1.1. Osmolytes: Chemical Chaperones Which Can Stabilize or Destabilize Proteins
1.2. Osmolytes and Protein Fibrillation
1.3. Glucagon: A Model System for Fibrillation
2. Results
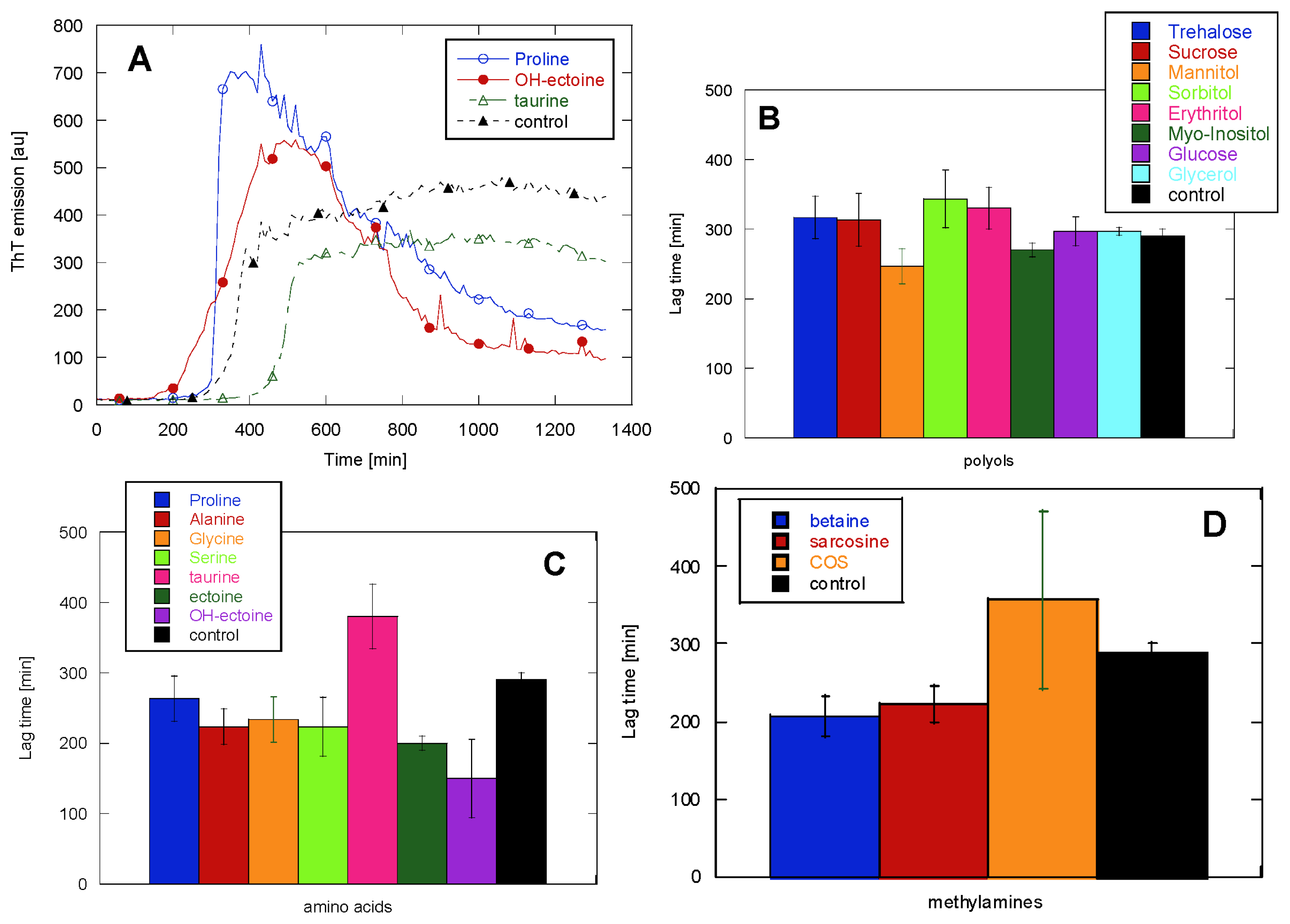
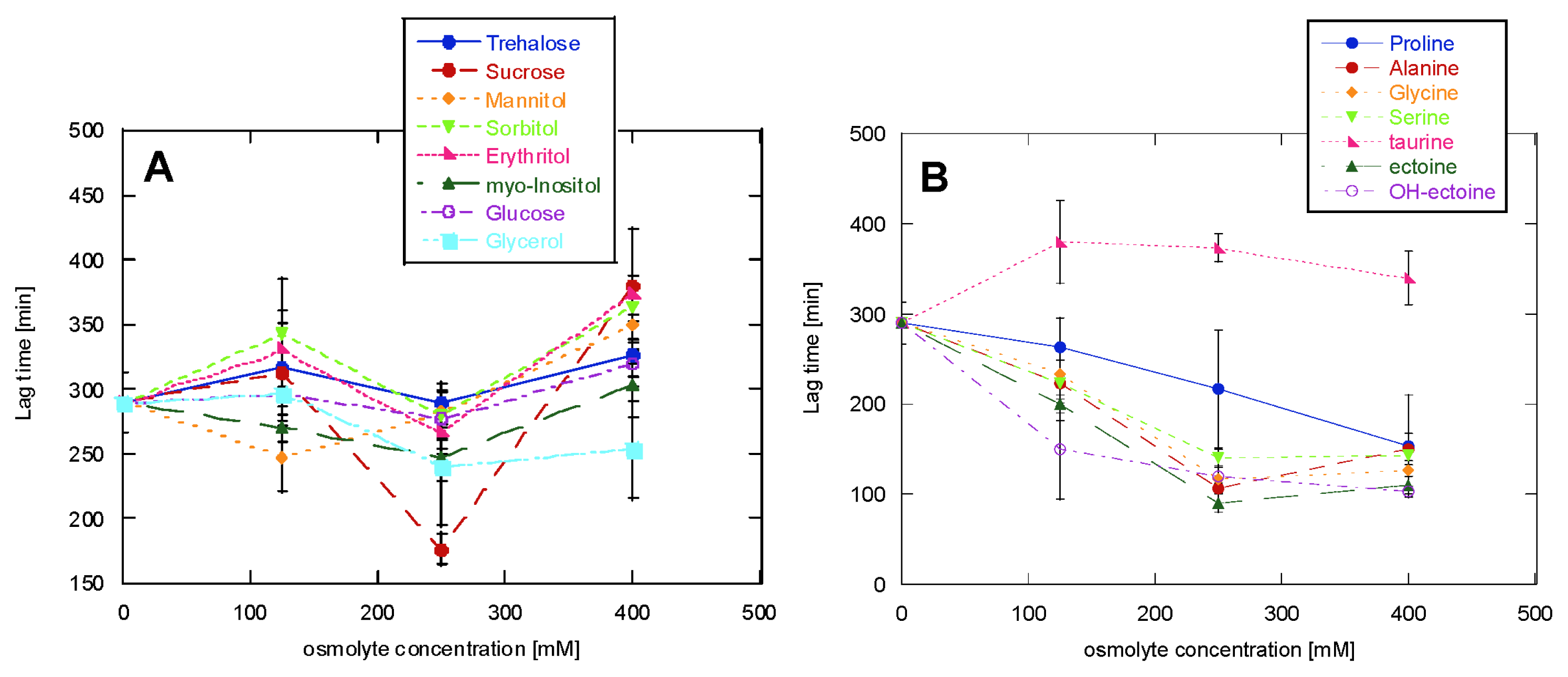
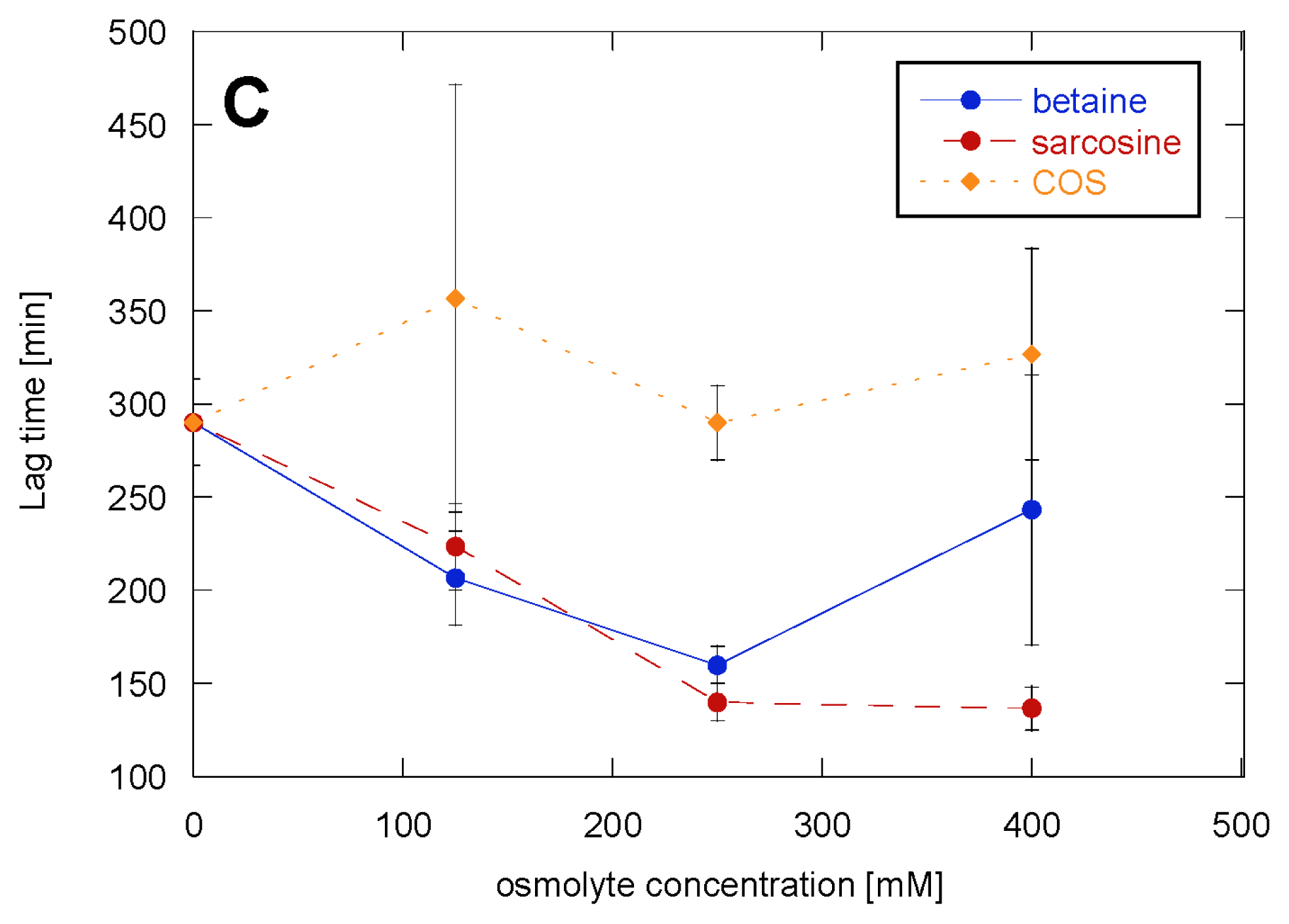
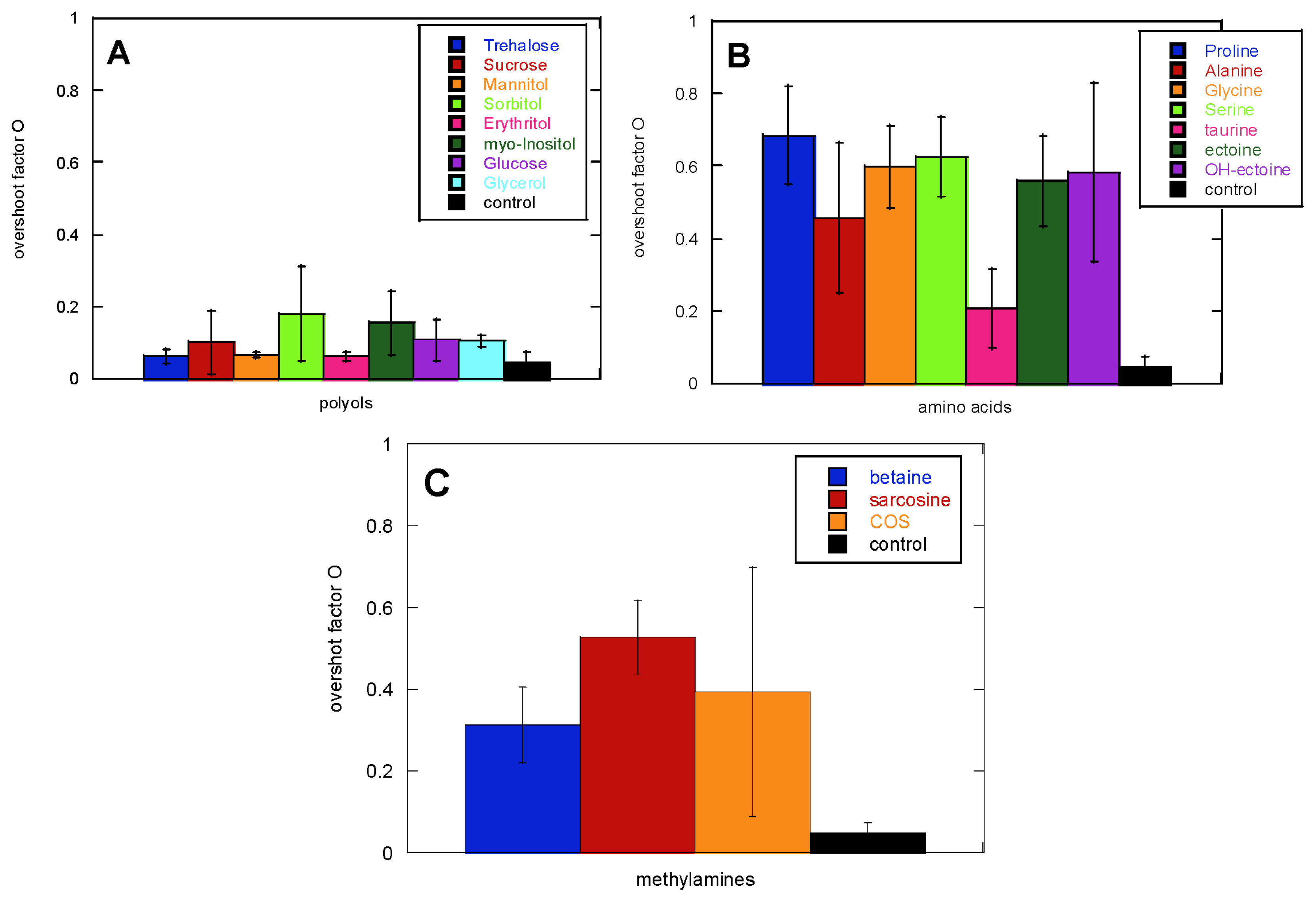
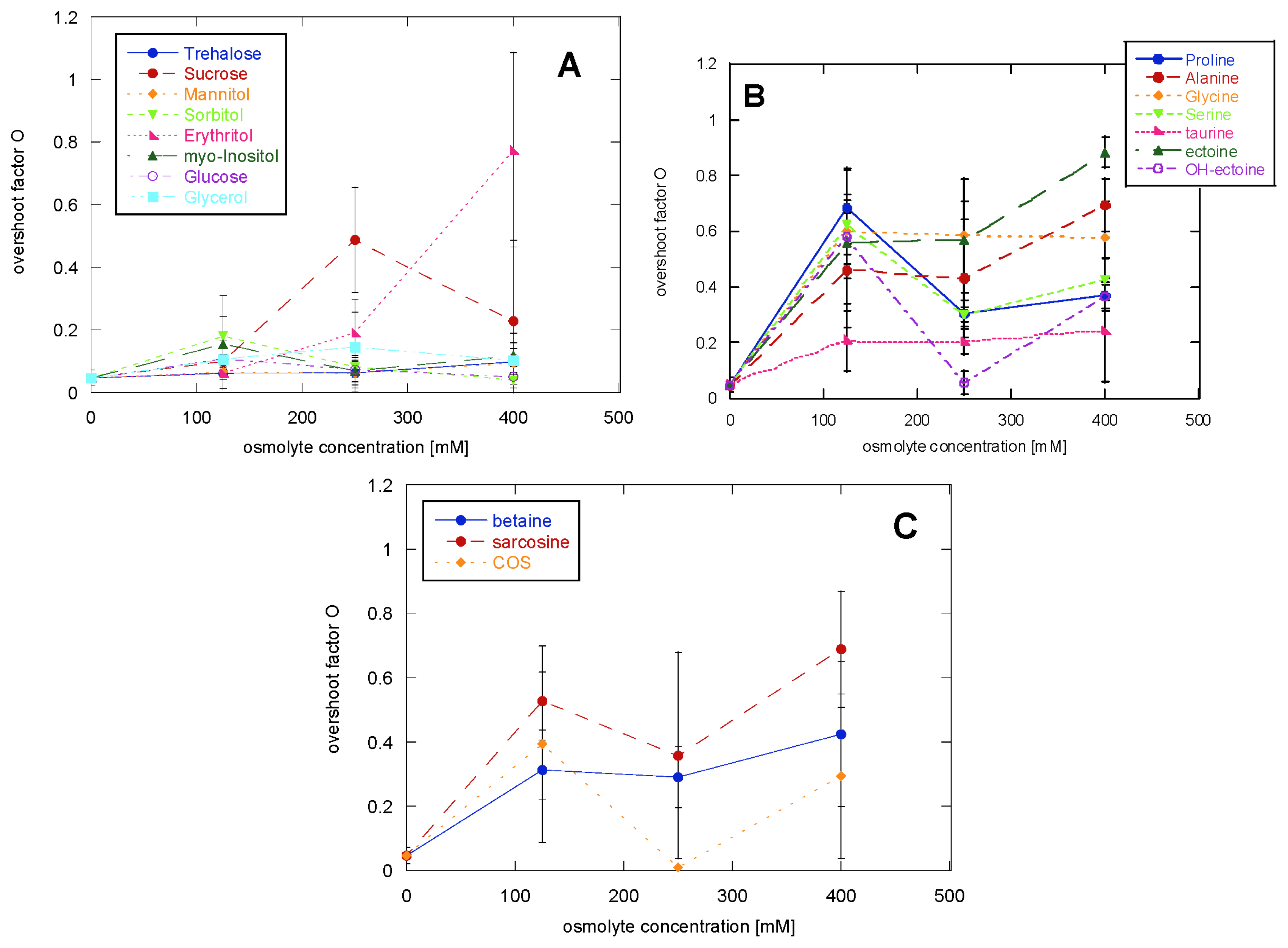
2.1. Glucagon Fibrillation Is Either Accelerated or Unaffected by Osmolytes
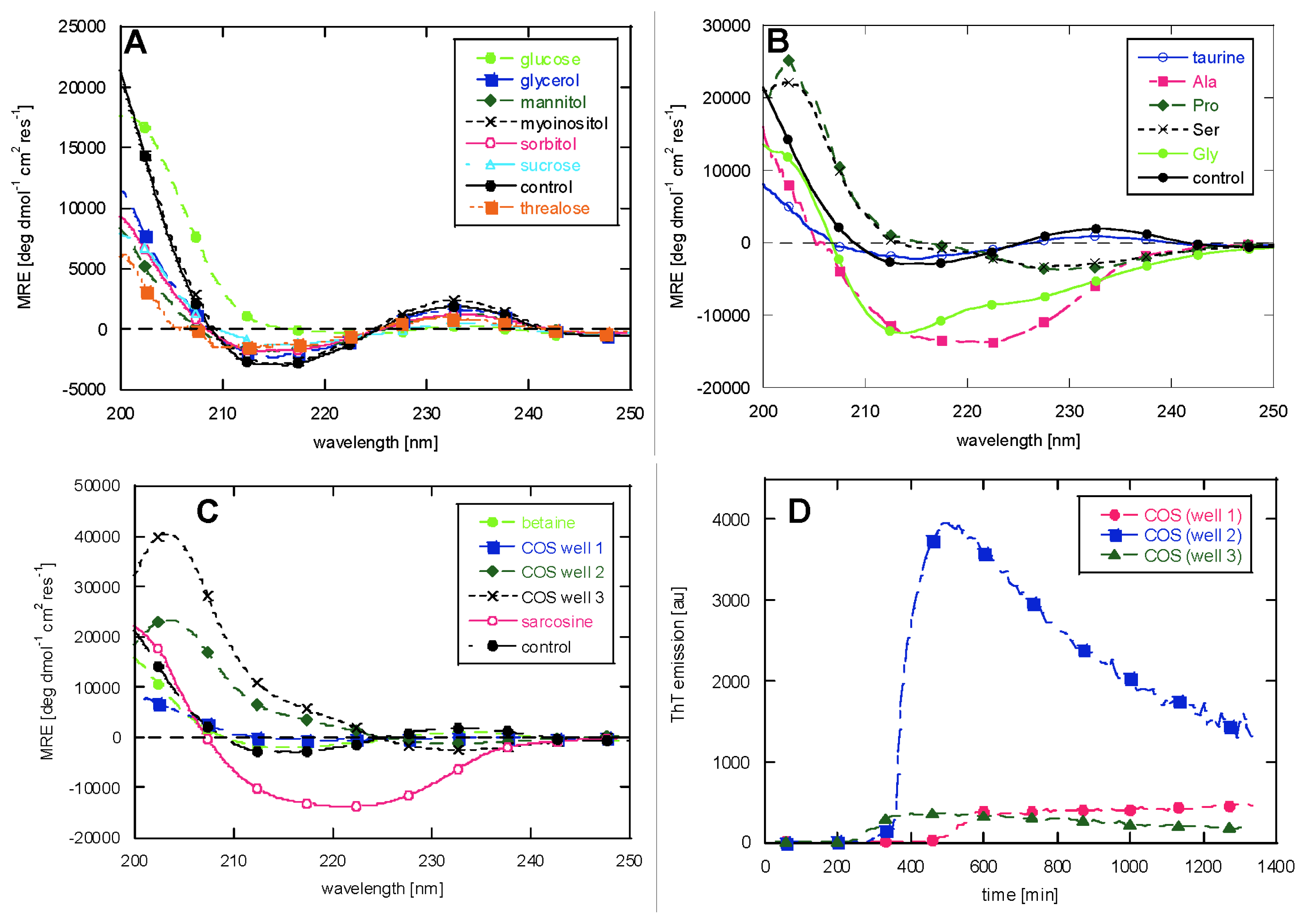
2.2. CD Reveals Glucagon Polymorphism in the Presence of Osmolytes
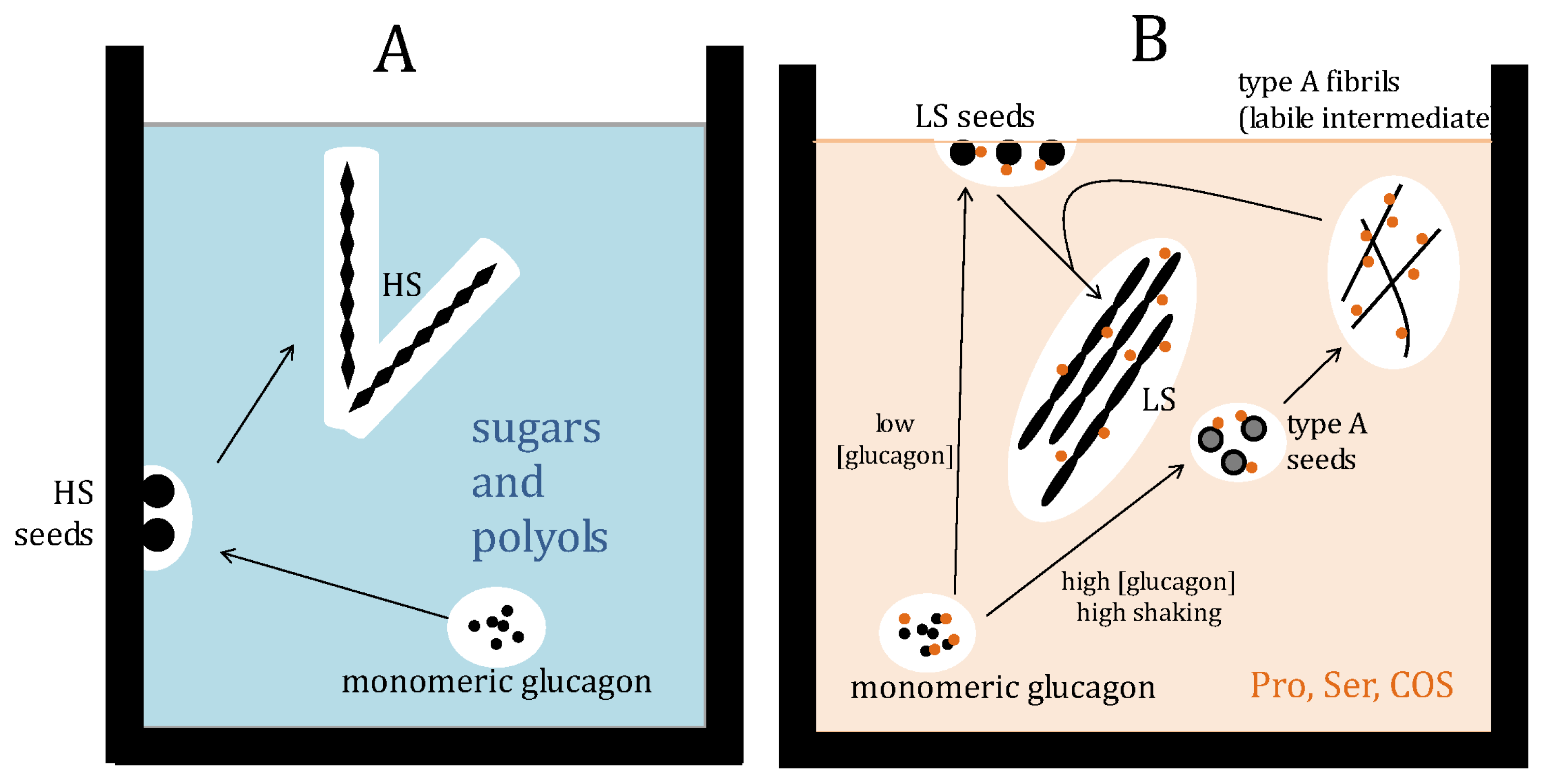
3. Discussion
4. Experimental Section
4.1. Glucagon Fibrillation in the Plate Reader
4.2. Circular Dichroism
5. Conclutions
Acknowledgments
References
- Bowlus, R.D.; Somero, G.N. Solute compatibility with enzyme function and structure: Rationales for the selection of osmotic agents and end-products of anaerobic metabolism in marine invertebrates. J. Exp. Zool 1979, 208, 137–151. [Google Scholar]
- Clark, M.E.; Hinke, J.A.M.; Todd, M.E. Studies on water in barnacle muscle fibres II. Role of ions and organic solutes in swelling of chemically-skinned fibres. J. Exp. Biol 1981, 90, 43–63. [Google Scholar]
- Yancey, P.H.; Clark, M.E.; Hand, S.C.; Bowlus, R.D.; Somero, G.N. Living with water stress: Evolution of osmolyte systems. Science 1982, 217, 1214–1222. [Google Scholar]
- Welch, W.J.; Brown, C.R. Influence of molecular and chemical chaperones on protein folding. Cell Stress Chaperones 1996, 1, 109–115. [Google Scholar]
- Meng, F.-G.; Park, Y.-D.; Zhou, H.-M. Role of proline, glycerol, and heparin as protein folding aids during refolding of rabbit muscle creatine kinase. Int. J. Biochem. Cell Biol 2001, 33, 701–709. [Google Scholar]
- Timasheff, S.N. Solvent stabilization of protein structure. Meth. Mol. Biol 1995, 40, 253–269. [Google Scholar]
- Bolen, D.W.; Baskakov, I.V. The osmophobic effect: Natural selection of a thermodynamic force in protein folding. J. Mol. Biol 2001, 310, 955–963. [Google Scholar]
- Singh, L.R.; Poddar, N.K.; Dar, T.A.; Kumar, R.; Ahmad, F. Protein and DNA destabilization by osmolytes: The other side of the coin. Life Sci 2011, 88, 117–125. [Google Scholar]
- Orsini, G.; Goldberg, M.E. The renaturation of reduced chymotrypsinogen A in guanidine HCl. Refolding versus aggregation. J. Biol. Chem 1978, 253, 3453–3458. [Google Scholar]
- Maeda, Y.; Koga, H.; Yamada, H.; Ueda, T.; Imoto, T. Effective renaturation of reduced lysozyme by gentle removal of urea. Protein Eng 1995, 8, 201–205. [Google Scholar]
- Yancey, P.H.; Somero, G.N. Counteraction of urea destabilization of protein structure by methylamine osmoregulatory compounds of elasmobranch fishes. Biochem. J 1979, 183, 317–323. [Google Scholar]
- Santoro, M.M.; Liu, Y.; Khan, S.M.A.; Hou, L.X.; Bolen, D.W. Increased thermal stability of proteins in the presence of naturally occurring osmolytes. Biochemistry 1992, 31, 5278–5283. [Google Scholar]
- Anjum, F.; Rishi, V.; Ahmad, F. Compatibility of osmolytes with Gibbs energy of stabilization of proteins. Biochim. Biophys. Acta 2000, 1476, 75–84. [Google Scholar]
- Nayak, A.; Lee, C.-C.; McRae, G.J.; Belfort, G. Osmolyte controlled fibrillation kinetics of insulin: New insight into fibrillation using the preferential exclusion principle. Biotechnol. Prog 2009, 25, 1508–1514. [Google Scholar]
- Jamal, S.; Poddar, N.K.; Singh, L.R.; Dar, T.A.; Rishi, V.; Ahmad, F. Relationship between functional activity and protein stability in the presence of all classes of stabilizing osmolytes. FEBS J 2009, 276, 6024–6032. [Google Scholar]
- Kaushik, J.K.; Bhat, R. Why Is Trehalose an Exceptional Protein Stabilizer? J. Biol. Chem 2003, 278, 26458–26465. [Google Scholar]
- Granata, V.; Palladino, P.; Tizzano, B.; Negro, A.; Berisio, R.; Zagari, A. The effect of the osmolyte trimethylamine N-oxide on the stability of the prion protein at low pH. Biopolymers 2006, 82, 234–240. [Google Scholar]
- Natalello, A.; Liu, J.; Ami, D.; Doglia, S.M.; de Marco, A. The osmolyte betaine promotes protein misfolding and disruption of protein aggregates. Protein Struct. Funct. Bioinforma 2009, 75, 509–517. [Google Scholar]
- Singh, L.R.; Dar, T.A.; Rahman, S.; Jamal, S.; Ahmad, F. Glycine betaine may have opposite effects on protein stability at high and low pH values. Biochim. Biophys. Acta 2009, 1794, 929–935. [Google Scholar]
- Hincha, D.K. High concentrations of the compatible solute glycinebetaine destabilize model membranes under stress conditions. Cryobiology 2006, 53, 58–68. [Google Scholar]
- Poddar, N.K.; Ansari, Z.A.; Singh, R.K.B.; Moosavi-Movahedi, A.A.; Ahmad, F. Effect of monomeric and oligomeric sugar osmolytes on ΔGd, the Gibbs energy of stabilization of the protein at different pH values: Is the sum effect of monosaccharide individually additive in a mixture? Biophys. Chem 2008, 138, 120–129. [Google Scholar]
- Scaramozzino, F.; Peterson, D.W.; Farmer, P.; Gerig, J.T.; Graves, D.J.; Lew, J. TMAO promotes fibrillization and microtubule assembly activity in the C-terminal repeat region of tau. Biochemistry 2006, 45, 3684–3691. [Google Scholar]
- Munishkina, L.A.; Henriques, J.; Uversky, V.N.; Fink, A.L. Role of protein-water interactions and electrostatics in alpha-synuclein fibril formation. Biochemistry 2004, 43, 3289–3300. [Google Scholar]
- Fung, J.; Darabie, A.A.; McLaurin, J. Contribution of simple saccharides to the stabilization of amyloid structure. Biochem. Biophys. Res. Commun 2005, 328, 1067–1072. [Google Scholar]
- Samuel, D.; Ganesh, G.; Yang, P.-W.; Chang, M.-M.; Wang, S.-L.; Hwang, K.-C.; Yu, C.; Jayaraman, G.; Kumar, T.K.S.; Trivedi, V.D.; Chang, D.-K. Proline inhibits aggregation during protein refolding. Protein Sci 2000, 9, 344–352. [Google Scholar]
- Ignatova, Z.; Gierasch, L.M. Inhibition of protein aggregation in vitro and in vivo by a natural osmoprotectant. Proc. Natl. Acad. Sci. USA 2006, 103, 13357–13361. [Google Scholar]
- Borwankar, T.; Röthlein, C.; Zhang, G.; Techen, A.; Dosche, C.; Ignatova, Z. Natural osmolytes remodel the aggregation pathway of mutant huntingtin exon 1. Biochemistry 2011, 50, 2048–2060. [Google Scholar]
- Uversky, V.N.; Li, J.; Fink, A.L. Trimethylamine-N-oxide-induced folding of α-synuclein. FEBS Lett 2001, 509, 31–35. [Google Scholar]
- Morar, A.S.; Olteanu, A.; Young, G.B.; Pielak, G.J. Solvent-induced collapse of α-synuclein and acid-denatured cytochrome c. Protein Sci 2001, 10, 2195–2199. [Google Scholar]
- Bomhoff, G.; Sloan, K.; McLain, C.; Gogol, E.P.; Fisher, M.T. The effects of the flavonoid baicalein and osmolytes on the Mg 2+ accelerated aggregation/fibrillation of carboxymethylated bovine 1SS-α-lactalbumin. Arch. Biochem. Biophys 2006, 453, 75–86. [Google Scholar]
- Nandi, P.K.; Bera, A.; Sizaret, P.Y. Osmolyte trimethylamine N-oxide converts recombinant α-helical prion protein to its soluble β-structured form at high temperature. J. Mol. Biol 2006, 362, 810–820. [Google Scholar]
- Tatzelt, J.; Prusiner, S.; Welch, W. Chemical chaperones interfere with the formation of scrapie prion protein. EMBO J 1996, 15, 6363–6373. [Google Scholar]
- Yang, D.-S.; Yip, C.M.; Huang, T.H.J.; Chakrabartty, A.; Fraser, P.E. Manipulating the amyloid-β aggregation pathway with chemical chaperones. J. Biol. Chem 1999, 274, 32970–32974. [Google Scholar]
- Scheibel, T.; Lindquist, S.L. The role of conformational flexibility in prion propagation and maintenance for Sup35p. Nat. Struct. Mol. Biol 2001, 8, 958–962. [Google Scholar]
- Kim, H.Y.; Kim, Y.; Han, G.; Kim, D.J. Regulation of in vitro Aβ1–40 Aggregation mediated by small molecules. J. Alzheimer’s Dis 2010, 22, 73–85. [Google Scholar]
- Liu, R.; Barkhordarian, H.; Emadi, S.; Park, C.B.; Sierks, M.R. Trehalose differentially inhibits aggregation and neurotoxicity of beta-amyloid 40 and 42. Neurobiol. Dis 2005, 20, 74–81. [Google Scholar]
- Qi, W.; Zhang, A.; Good, T.A.; Fernandez, E.J. Two disaccharides and trimethylamine N-oxide affect Aβ aggregation differently, but all attenuate oligomer-induced membrane permeability. Biochemistry 2009, 48, 8908–8919. [Google Scholar]
- Singer, M.A.; Lindquist, S. Multiple effects of trehalose on protein folding in vitro and in vivo. Mol. Cell 1998, 1, 639–648. [Google Scholar]
- Tanaka, M.; Machida, Y.; Niu, S.; Ikeda, T.; Jana, N.R.; Doi, H.; Kurosawa, M.; Nekooki, M.; Nukina, N. Trehalose alleviates polyglutamine-mediated pathology in a mouse model of Huntington disease. Nat. Med 2004, 10, 148–154. [Google Scholar]
- Kanapathipillai, M.; Lentzen, G.; Sierks, M.; Park, C.B. Ectoine and hydroxyectoine inhibit aggregation and neurotoxicity of Alzheimer’s β-amyloid. FEBS Lett 2005, 579, 4775–4780. [Google Scholar]
- Hudson, S.A.; Ecroyd, H.; Kee, T.W.; Carver, J.A. The thioflavin T fluorescence assay for amyloid fibril detection can be biased by the presence of exogenous compounds. FEBS J 2009, 276, 5960–5972. [Google Scholar]
- Liu, K.-N.; Wang, H.-Y.; Chen, C.-Y.; Wang, S. l-Arginine reduces thioflavin T fluorescence but not fibrillation of bovine serum albumin. Amino Acids 2010, 39, 821–829. [Google Scholar]
- Pedersen, J.S.; Dikov, D.; Flink, J.L.; Hjuler, H.A.; Christiansen, G.; Otzen, D.E. The changing face of glucagon fibrillation: Structural polymorphism and conformational imprinting. J. Mol. Biol 2006, 355, 501–523. [Google Scholar]
- Arora, A.; Ha, C.; Park, C.B. Inhibition of insulin amyloid formation by small stress molecules. FEBS Lett 2004, 564, 121–125. [Google Scholar]
- Ueda, T.; Nagata, M.; Imoto, T. Aggregation and chemical reaction in hen lysozyme caused by heating at pH 6 are depressed by osmolytes, sucrose and trehalose. J. Biochem 2001, 130, 491–496. [Google Scholar]
- Kim, Y.-S.; Cape, S.P.; Chi, E.; Raffen, R.; Wilkins-Stevens, P.; Stevens, F.J.; Manning, M.C.; Randolph, T.W.; Solomon, A.; Carpenter, J.F. Counteracting effects of renal solutes on amyloid fibril formation by immunoglobulin light chains. J. Biol. Chem 2001, 276, 1626–1633. [Google Scholar]
- Tanaka, M.; Machida, Y.; Nukina, N. A novel therapeutic strategy for polyglutamine diseases by stabilizing aggregation-prone proteins with small molecules. J. Mol. Med 2005, 83, 343–352. [Google Scholar]
- Dong, X.-Y.; Liu, J.-H.; Liu, F.-F.; Sun, Y. Self-interaction of native and denatured lysozyme in the presence of osmolytes, l-arginine and guanidine hydrochloride. Biochem. Eng. J 2009, 43, 321–326. [Google Scholar]
- Pedersen, J.S.; Andersen, C.B.; Otzen, D.E. Amyloid structure—One but not the same: The many levels of fibrillar polymorphism. FEBS J 2010, 277, 4591–4601. [Google Scholar]
- Hochacka, P.W.; Somero, G.N. Water-Solute Adaptations: The Evolution and Regulation of the Internal Milieu; Oxford University Press: New York NY, USA, 2002. [Google Scholar]
- Pedersen, J.S.; Dikov, D.; Flink, J.L.; Hjuler, H.A.; Christiansen, G.; Otzen, D.E. The changing face of glucagon fibrillation: Structural polymorphism and conformational imprinting. J. Mol. Biol 2006, 355, 501–523. [Google Scholar]
- Ghodke, S.; Nielsen, S.B.; Christiansen, G.; Hjuler, H.A.; Flink, J.; Otzen, D.E. Mapping out the multi-stage fibrillation of glucagon. FEBS J 2011, 279, 752–765. [Google Scholar]
- Macchi, F.; Hoffmann, S.V.; Carlsen, M.; Vad, B.; Imparato, A.; Rischel, C.; Otzen, D.E. Mechanical stress affects glucagon fibrillation kinetics and fibril structure. Langmuir 2011, 27, 12539–12549. [Google Scholar]
- Macchi, F.; Hoffmann, S.V.; Carlsen, M.; Vad, B.; Imparato, A.; Rischel, C.; Otzen, D.E. Mechanical Stress Affects Glucagon Fibrillation Kinetics and fibril Structure. Langmuir 2011, 27, 12539–12549. [Google Scholar]
- Woody, R.W. Contributions of tryptophan side chains to the far-ultraviolet circular dichroism of proteins. Eur. Biophys. J 1994, 23, 253–262. [Google Scholar]
- Andersen, C.B.; Hicks, M.R.; Vetri, V.; Vandahl, B.; Rahbek-Nielsen, H.; Thøgersen, H.; Thøgersen, I.B.; Enghild, J.J.; Serpell, L.C.; Rischel, C.; et al. Glucagon fibril polymorphism reflects differences in protofilament backbone structure. J. Mol. Biol 2010, 397, 932–946. [Google Scholar]
- Pedersen, J.S.; Dikov, D.; Otzen, D.E. N- and C-terminal hydrophobic patches are involved in fibrillation of glucagon. Biochemistry 2006, 45, 14503–14512. [Google Scholar]
- Uversky, V.N.; Cooper, M.E.; Bower, K.S.; Li, J.; Fink, A.L. Accelerated α-synuclein fibrillation in crowded milieu. FEBS Lett 2002, 515, 99–103. [Google Scholar]
- Sasahara, K.; McPhie, P.; Minton, A.P. Effect of dextran on protein stability and conformation attributed to macromolecular crowding. J. Mol. Biol 2003, 326, 1227–1237. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Osmolyte | Aβ40 | Aβ42 | α-synuclein | polyQ | Immunoglobulin light chains | Glucagon 1 | insulin | lysozyme |
|---|---|---|---|---|---|---|---|---|
| Glucose | oligomerization [24] | oligomerization [24] | collapse | no effect | minor effect [14] | |||
| Sucrose | oligomerization [24] | oligomerization [24] | no effect | no/minor effect [14,44] | reduced aggregation [25,45] | |||
| Fructose | no effect[24] | no effect [24] | minor effect [14] | |||||
| Galactose | induced fibrillation [24] | induced fibrillation [24] | ||||||
| Mannose | induced fibrillation [24] | induced fibrillation [24] | ||||||
| Sorbitol | longer lag, more stable fibrils [46] | no effect | ||||||
| Glycerol | oligomerization [33] | no effect | no effect [25] | |||||
| Trehalose | slower/no fibrillation [36,37] | oligomerization (toxic) [36] | reduced aggregation in mice [47] | no effect | reduced/slower fibrillation [14,44] | increasing B1 [48], reduced aggregation [45] | ||
| Gly | faster fibrillation [35] | faster fibrilation | no effect [25] | |||||
| Pro | amorphous aggregates [27] | polymorphism | inhibits aggregation [25] | |||||
| Taurine | faster fibrillation [35] | slower fibrillation | ||||||
| Ectoine | longer lag time, more oligomers, lower toxicity[40] | faster fibrilation | highly reduced fibrillation [44] | |||||
| Betaine | faster fibrillation [27] | longer lag, more stable fibrils [46] | faster fibrilation | highly reduced fibrillation [44] | increasing B2 [48] | |||
| Sarcosine | faster fibrillation, polymorphism | Tm of unfolding increases by 22°, but the unfolded lysozyme aggregates [12] | ||||||
| TMAO | little effect [37]; oligomerization [33] | folded oligomer/enhanced fibrillation [28] | amorphous aggregates [27] |
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Macchi, F.; Eisenkolb, M.; Kiefer, H.; Otzen, D.E. The Effect of Osmolytes on Protein Fibrillation. Int. J. Mol. Sci. 2012, 13, 3801-3819. https://doi.org/10.3390/ijms13033801
Macchi F, Eisenkolb M, Kiefer H, Otzen DE. The Effect of Osmolytes on Protein Fibrillation. International Journal of Molecular Sciences. 2012; 13(3):3801-3819. https://doi.org/10.3390/ijms13033801
Chicago/Turabian StyleMacchi, Francesca, Maike Eisenkolb, Hans Kiefer, and Daniel E. Otzen. 2012. "The Effect of Osmolytes on Protein Fibrillation" International Journal of Molecular Sciences 13, no. 3: 3801-3819. https://doi.org/10.3390/ijms13033801