A Single-Chamber Microbial Fuel Cell without an Air Cathode

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
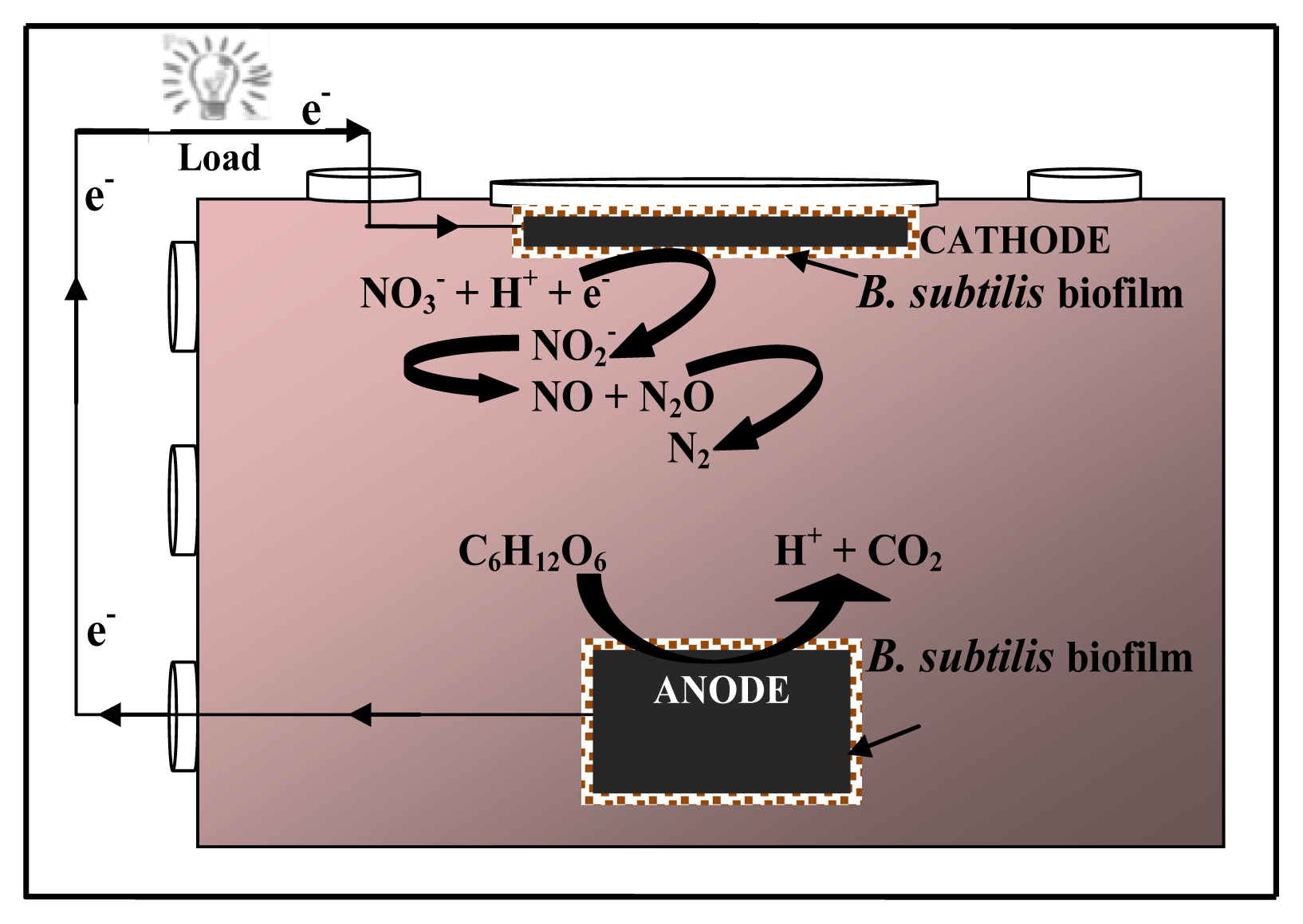
2.1. Nitrate Respiration and Current Generation
2.2. Effect of Nitrate Addition During Different Batch Cycles and Its Relationship with Current Generation and Nitrate Reduction
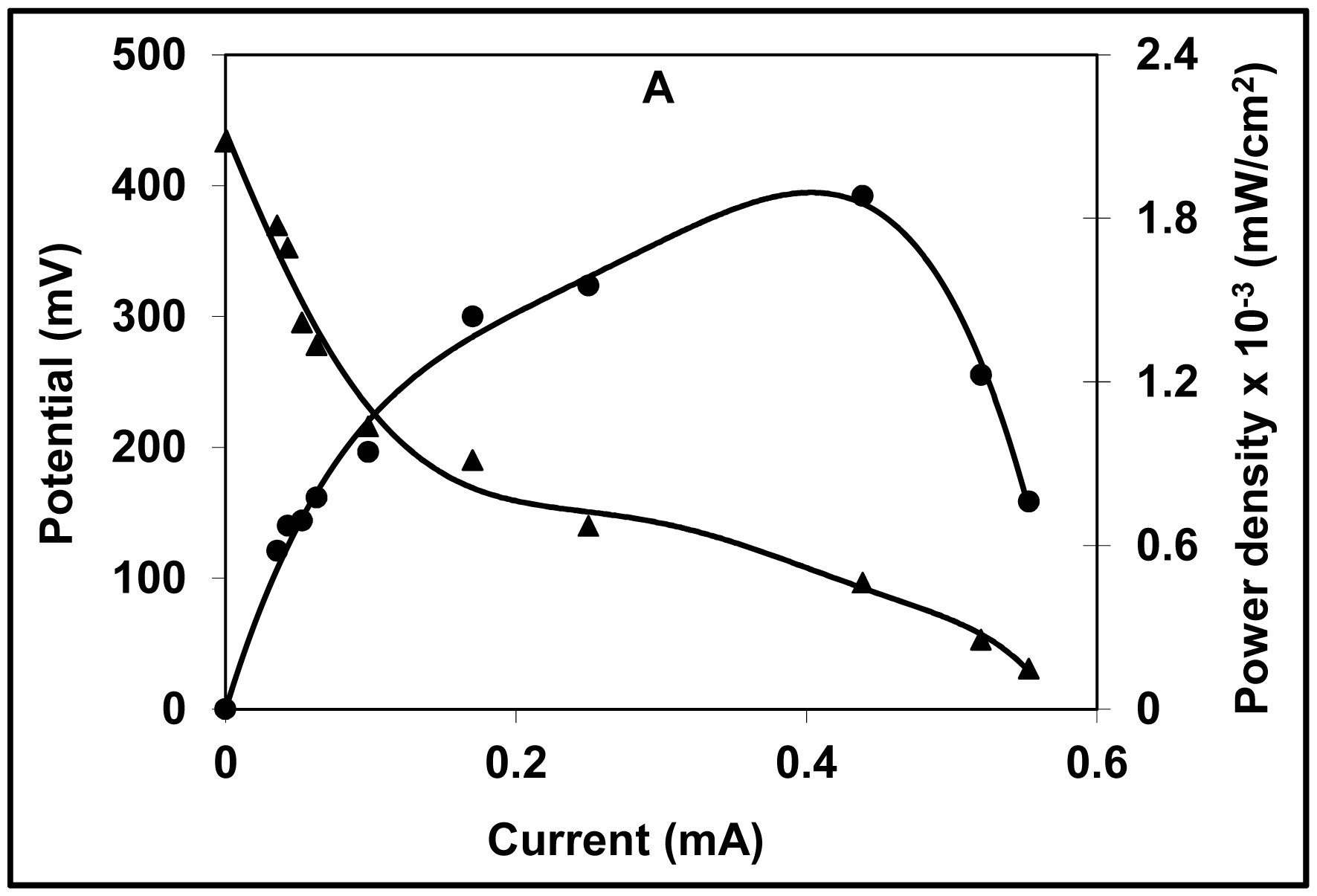
2.3. Polarization Curve
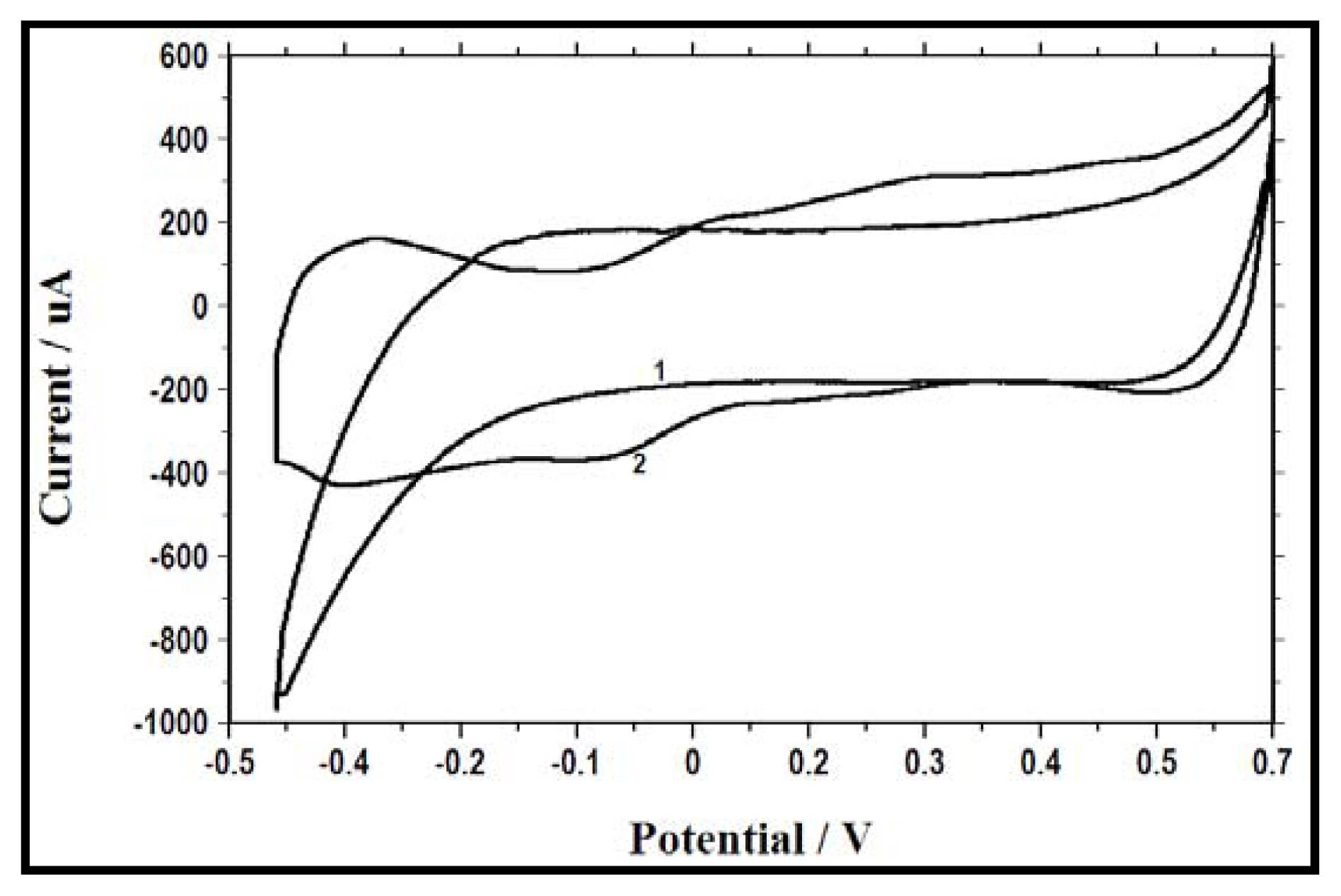
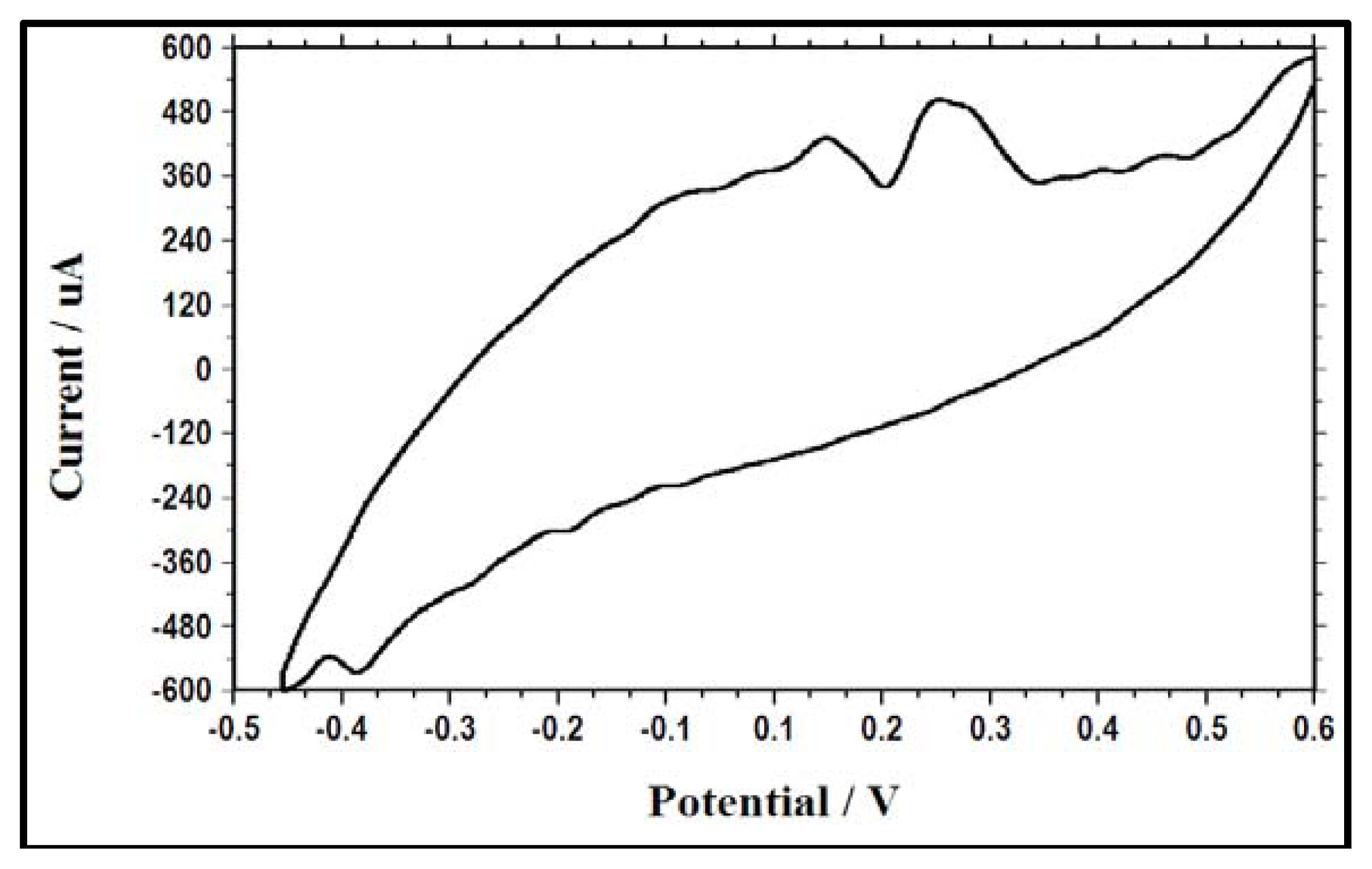
2.4. Electrochemical Activity as a Result of Change in Electron Acceptor
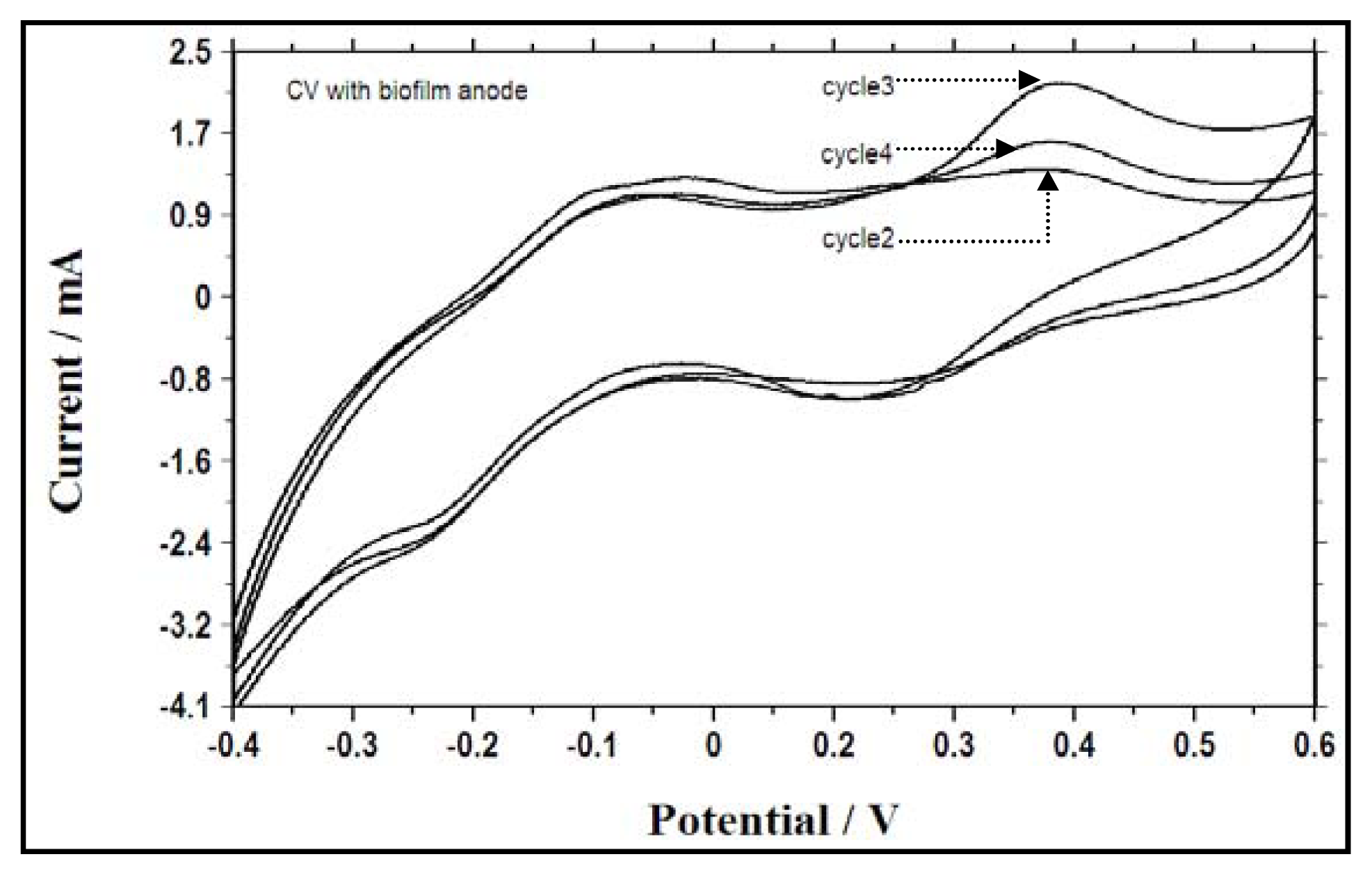
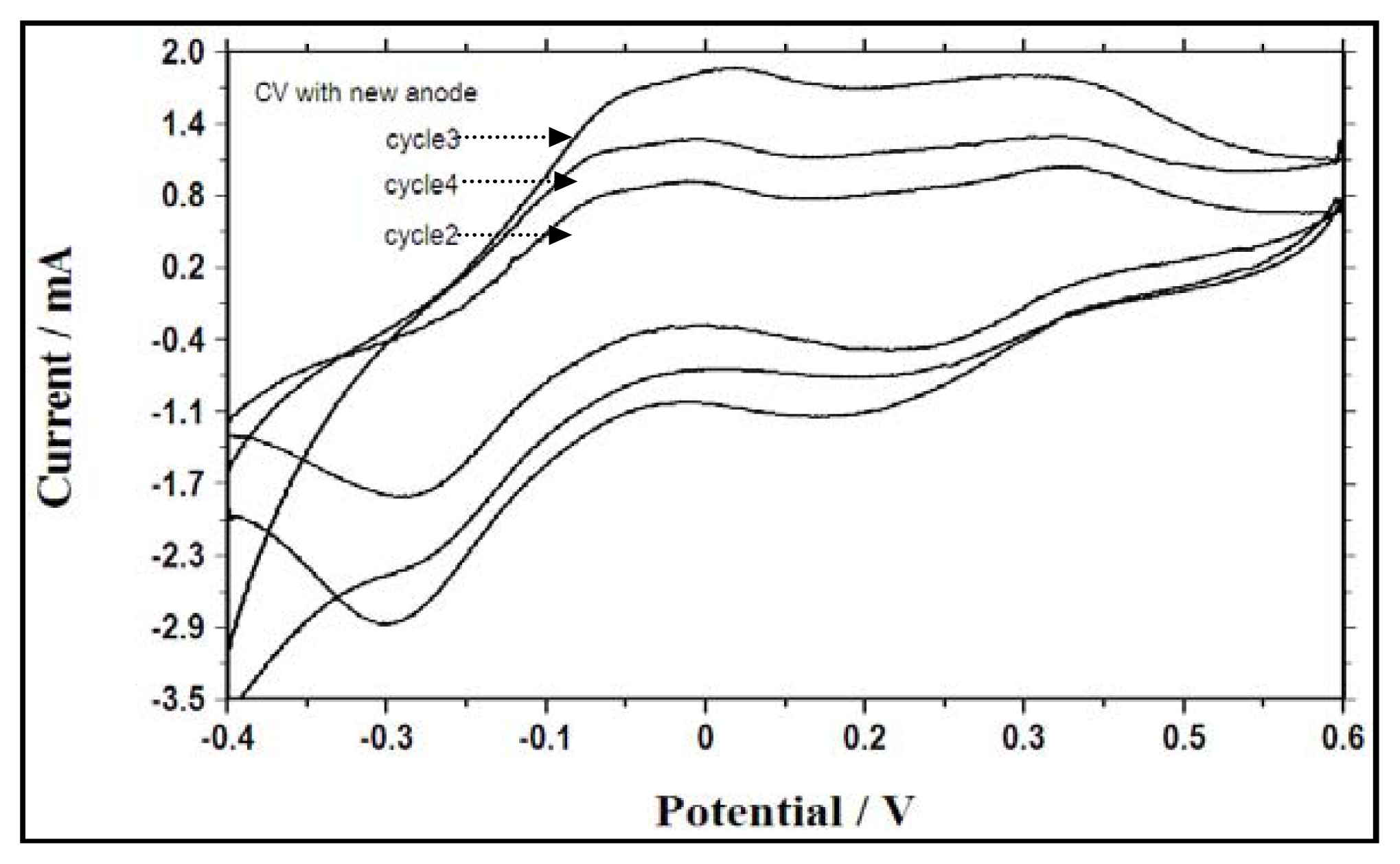
2.5. Electrochemical Activity During Different Fed Batch Cycles and Electron Transfer Mechanisms
2.6. Performance of the MFC at the End of the Batch Test
2.7. Coulombic Efficiency (CE)
3. Experimental Section
3.1. Cultivation of Bacteria
3.2. Microbial Fuel Cell Construction and Operation
3.3. Biofilm Growth and Anaerobic Condition
3.4. Optical Density Measurement
3.5. Nitrate Measurement
3.6. Data Acquisition and Electrochemical Technique
3.7. Cyclic Voltammetry
4. Conclusions
Acknowledgments
References
- Ra, C.S.; Lo, K.V.; Shin, J.S.; Oh, J.S.; Hong, B.J. Biological nutrient removal with an integral organic carbon source in piggery wastewater treatment. Water Res 2000, 34, 965–973. [Google Scholar]
- Shukla, A.K.; Suresh, P.; Berchmans, S.; Rajendran, A. Biological fuel cells and their applications. Curr. Sci 2004, 87, 455–468. [Google Scholar]
- Clauwaert, P.; Rabaey, K.; Aelterman, P.; de Schamphelaire, L.; Pham, T.H.; Boeckx, P.; Boon, N.; Verstraete, W. Biological denitrification in microbial fuel cells. Environ. Sci. Technol 2007, 41, 3354–3360. [Google Scholar]
- Bond, D.R.; Lovely, D.R. Electricity production by Geobacter sulfurreducens attached to electrodes. Appl. Environ. Microb 2003, 69, 1548–1555. [Google Scholar]
- Sukkasem, C.; Xu, S.; Park, S.; Boonsawang, P.; Liu, H. Effect of nitrate on the performance of single chamber air cathode microbial fuel cells. Water Res 2008, 42, 4743–4750. [Google Scholar]
- Rabaey, K.; Rodríguez1, J.; Blackall, L.L.; Keller, J.; Gross, P.; Batstone, D.; Verstraete, W.; Nealson, K.H. Microbial ecology meets electrochemistry: Electricity-driven and driving communities. ISME J 2007, 1, 9–18. [Google Scholar]
- Gregory, K.B.; Bond, D.R.; Lovely, D.R. Graphite electrodes as electron donors for anaerobic respiration. Environ. Microbiol 2004, 6, 596–604. [Google Scholar]
- An, J.; Moon, H.; Lee, L.; Chang, I.S. Bifunctional silver nanoparticle cathode in microbial fuel cells for microbial growth inhibition with comparable oxygen reduction reaction activity. Environ. Sci. Technol 2011, 45, 5441–5446. [Google Scholar]
- An, J.; Moon, H.; Chang, I.S. Multiphase electrode microbial fuel cell system that simultaneously converts organics coexisting in water and sediment phases into electricity. Environ. Sci. Technol 2010, 44, 7145–7150. [Google Scholar]
- Harnisch, F.; Wirth, S.; Schröder, U. Effects of substrate and metabolite crossover on the cathodic oxygen reduction reaction in microbial fuel cells: Platinum vs. iron(II) phthalocyanine based electrodes. Electrochem. Commun 2009, 11, 2253–2256. [Google Scholar]
- Logan, B.E.; Hamelers, B.; Rozendal, R.; Schröder, U.; Keller, J.; Freguia, S.; Aelterman, P.; Verstraete, W.; Rabaey, K. Microbial fuel cell: Methodology and technology. Environ. Sci. Technol 2006, 40, 5181–5192. [Google Scholar]
- Nimje, V.R.; Chen, C.Y.; Chen, C.C.; Chen, H.R.; Tseng, M.J.; Jean, J.S; Chang, Y.-F. Glycerol degradation in single chamber microbial fuel cell. Bioresour. Technol. 2011, 102, 2629–2634. [Google Scholar]
- Nimje, V.R.; Chen, C.Y.; Chen, C.C.; Jean, J.S.; Reddy, A.S.; Fan, C.W.; Pan, K.Y.; Liu, H.T.; Chen, J.L. Stable and high energy generation by a strain of Bacillus subtilis in a microbial fuel cell. J. Power Sources 2009, 190, 258–263. [Google Scholar]
- Nakano, M.M.; Zuber, P. Anaerobic growth of a “strict aerobe” (Bacillus subtilis). Annu. Rev. Microbiol 1998, 52, 165–190. [Google Scholar]
- Park, H.S.; Kim, B.H.; Kim, H.S.; Kim, H.J.; Kim, G.T.; Kim, M.; Chang, I.S.; Park, Y.K.; Chang, H.I. A novel electrochemically active and Fe(III)-reducing bacterium phylogenetically related to Clostridium butyricum isolated from a microbial fuel cell. Anaerobe 2001, 7, 297–306. [Google Scholar]
- Broman, K.; Lauwers, N.; Stalon, V.; Wiame, J.M. Oxygen and nitrate in utilization by Bacillus licheniformis of the arginase and arginine deiminase routes of arginine catabolism and other factors affecting their syntheses. J. Bacteriol 1978, 135, 920–927. [Google Scholar]
- Wauven, V.; Piérard, C.A.; Kley-Raymann, M.; Haas, D. Pseudomonas aeruginosa mutants affected in anaerobic growth on arginine: Evidence for a four-gene cluster encoding the arginine deiminase pathway. J. Bacteriol 1984, 160, 928–934. [Google Scholar]
- Cunin, R.; Glansdorff, N.; Pierard, A.; Stalon, V. Biosynthesis and metabolism of arginine in bacteria. Microbiol. Rev 1986, 50, 314–352. [Google Scholar]
- Nakano, M.M.; Dailly, Y.P.; Zuber, P.; Clark, D.P. Characterization of anaerobic fermentative growth of Bacillus subtilis: Identification of fermentation end products and genes required for growth. J. Bacteriol 1997, 179, 6749–6755. [Google Scholar]
- Benziger, J.B.; Satterfield, M.B.; Hogarth, W.H.J.; Nehlsen, J.P.; Kevrekidis, I.G. The power performance curve for engineering analysis of fuel cells. J. Power Sources 2006, 155, 272–285. [Google Scholar]
- Freguia, S.; Rabaey, K.; Yuan, Z.; Keller, J. Sequential anode-cathode configuration improves cathodic oxygen reduction and effluent quality of microbial fuel cells. Water Res 2008, 42, 1387–1396. [Google Scholar]
- Rabaey, K.; Boon, N; Siciliano, S.D.; Verhaege, M.; Verstraete, W. Biofuel cells select for microbial consortia that self-mediate electron transfer. Appl. Environ. Microb. 2004, 70, 5373–5382. [Google Scholar]
- Freguia, S.; Masuda, M.; Tsujimura, S.; Kano, K. Lactococcus lactis catalyses electricity generation at microbial fuel cell anodes via excretion of a soluble quinone. Bioelectrochemistry 2009, 76, 14–18. [Google Scholar]
- Liu, H.; Logan, B.E. Electricity generation using an air-cathode single chamber microbial fuel cell in the presence and absence of a proton exchange membrane. Environ. Sci. Technol 2004, 38, 4040–4046. [Google Scholar]
- Miller, H. Experiments in Molecular Genetics; CSHL Press: Plainview, NY, USA, 1972. [Google Scholar]
- Cheng, S.; Liu, H.; Logan, B.E. Increased performance of single-chamber microbial fuel cells using an improved cathode structure. Electrochem. Commun 2006, 8, 489–494. [Google Scholar]
- Nicholas, D.J.D.; Nason, A. Determination of Nitrate and Nitrite. In Methods in Enzymology; Academic Press Inc: New York NY, USA, 1957; Volume 3. [Google Scholar]
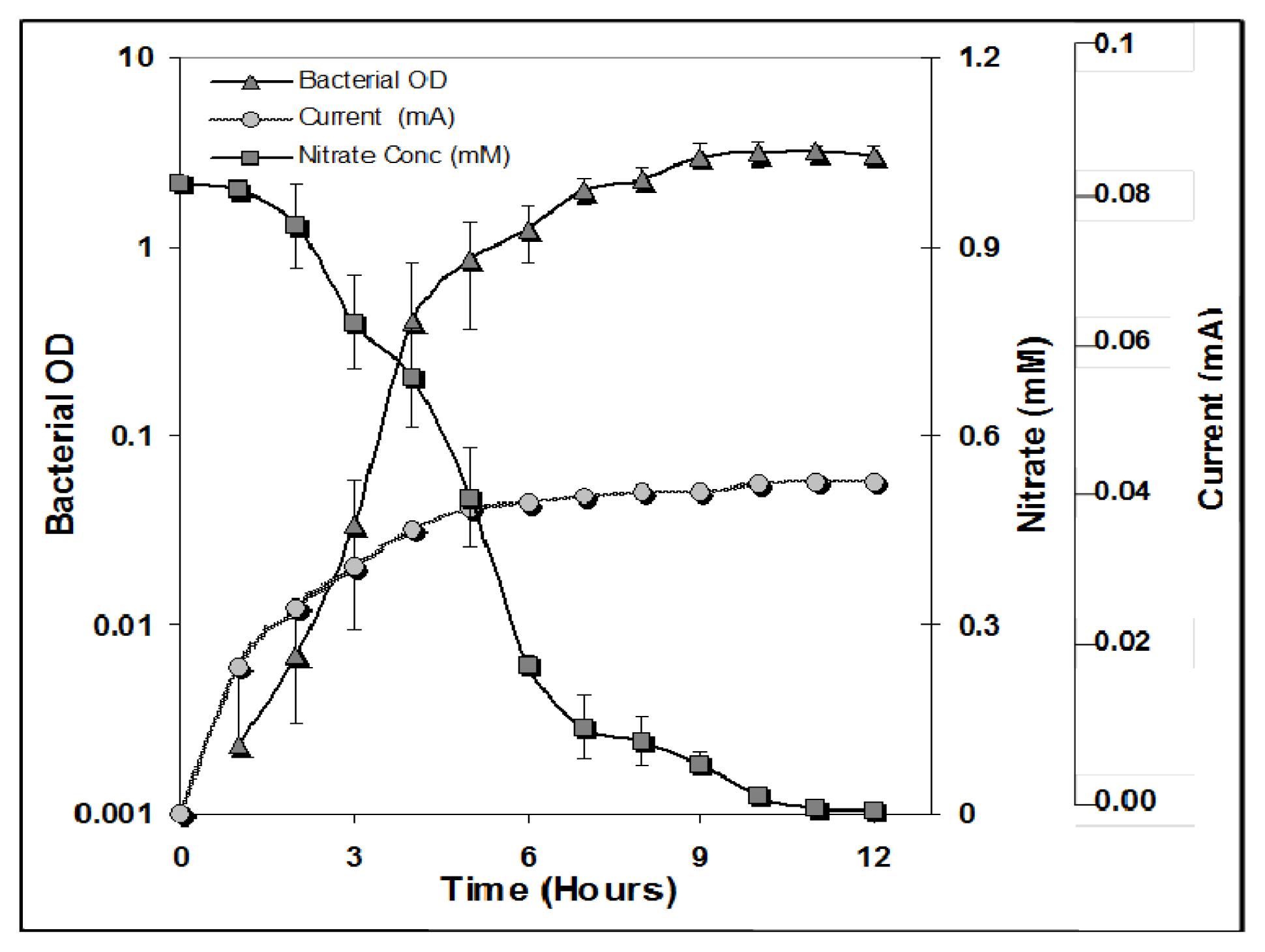
 ) with nitrate reduction (
) with nitrate reduction (
 ) and an increase in current (
) and an increase in current (
 ). The concentrations given were determined at the indicated time points in the Bacillus subtilis cultures with 1 mM nitrate and 1 mM glucose. The error bars indicate the standard deviation based on the average of three separate experiments.
) with nitrate reduction (
) and an increase in current (
). The concentrations given were determined at the indicated time points in the Bacillus subtilis cultures with 1 mM nitrate and 1 mM glucose. The error bars indicate the standard deviation based on the average of three separate experiments.
). The concentrations given were determined at the indicated time points in the Bacillus subtilis cultures with 1 mM nitrate and 1 mM glucose. The error bars indicate the standard deviation based on the average of three separate experiments.
) with nitrate reduction (
) and an increase in current (
). The concentrations given were determined at the indicated time points in the Bacillus subtilis cultures with 1 mM nitrate and 1 mM glucose. The error bars indicate the standard deviation based on the average of three separate experiments.
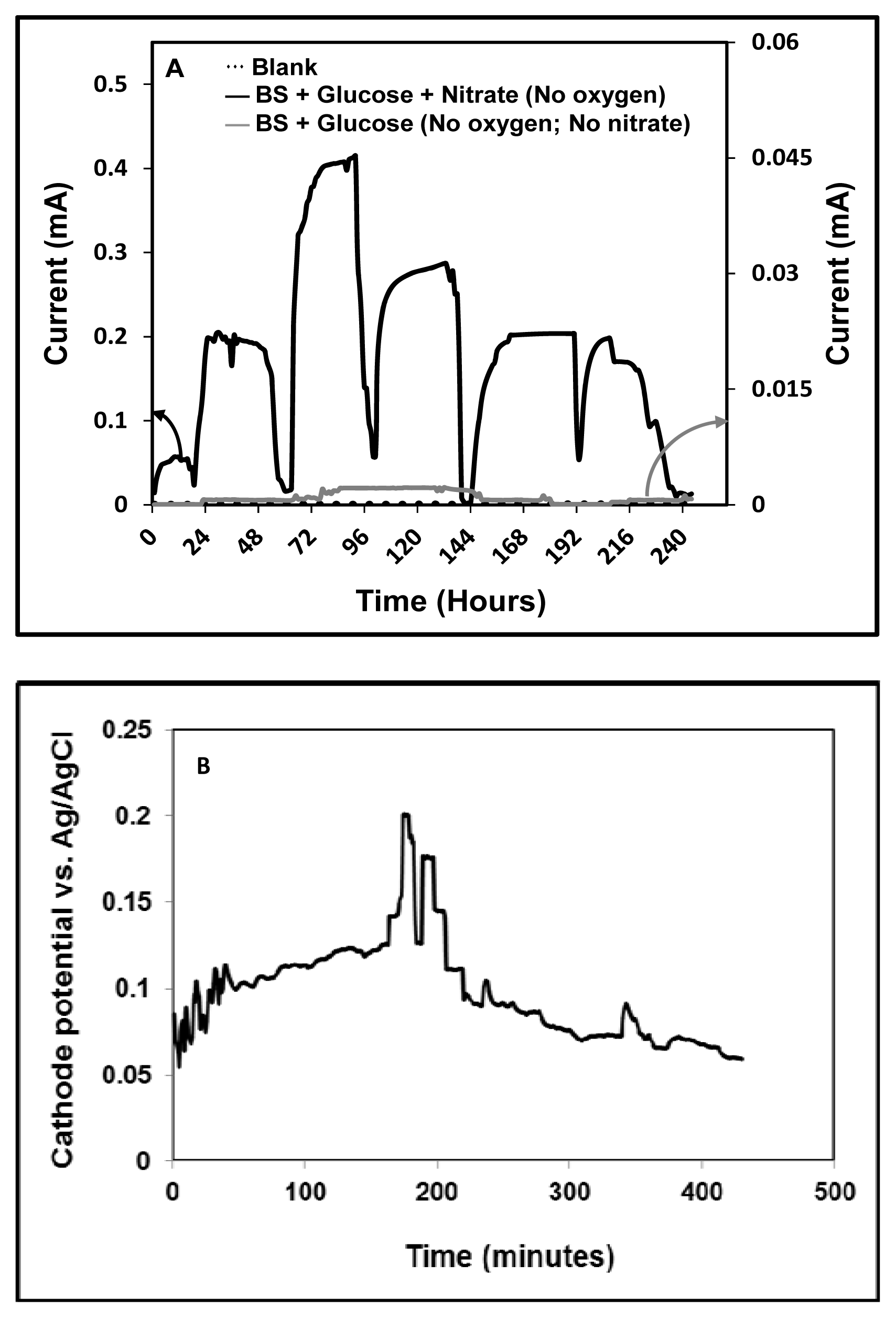
 ), and glucose with nitrate-fed (
), and glucose with nitrate-fed (
 ); (B) Cathode half-cell potential recorded at the end of the experiment.
), and glucose with nitrate-fed (
); (B) Cathode half-cell potential recorded at the end of the experiment.
); (B) Cathode half-cell potential recorded at the end of the experiment.
), and glucose with nitrate-fed (
); (B) Cathode half-cell potential recorded at the end of the experiment.







© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Nimje, V.R.; Chen, C.-C.; Chen, H.-R.; Chen, C.-Y.; Tseng, M.-J.; Cheng, K.-C.; Shih, R.-C.; Chang, Y.-F. A Single-Chamber Microbial Fuel Cell without an Air Cathode. Int. J. Mol. Sci. 2012, 13, 3933-3948. https://doi.org/10.3390/ijms13033933
Nimje VR, Chen C-C, Chen H-R, Chen C-Y, Tseng M-J, Cheng K-C, Shih R-C, Chang Y-F. A Single-Chamber Microbial Fuel Cell without an Air Cathode. International Journal of Molecular Sciences. 2012; 13(3):3933-3948. https://doi.org/10.3390/ijms13033933
Chicago/Turabian StyleNimje, Vanita Roshan, Chien-Cheng Chen, Hau-Ren Chen, Chien-Yen Chen, Min-Jen Tseng, Kai-Chien Cheng, Ruey-Chyuan Shih, and Young-Fo Chang. 2012. "A Single-Chamber Microbial Fuel Cell without an Air Cathode" International Journal of Molecular Sciences 13, no. 3: 3933-3948. https://doi.org/10.3390/ijms13033933