Dihydrolipoic Acid Induces Cytotoxicity in Mouse Blastocysts through Apoptosis Processes

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
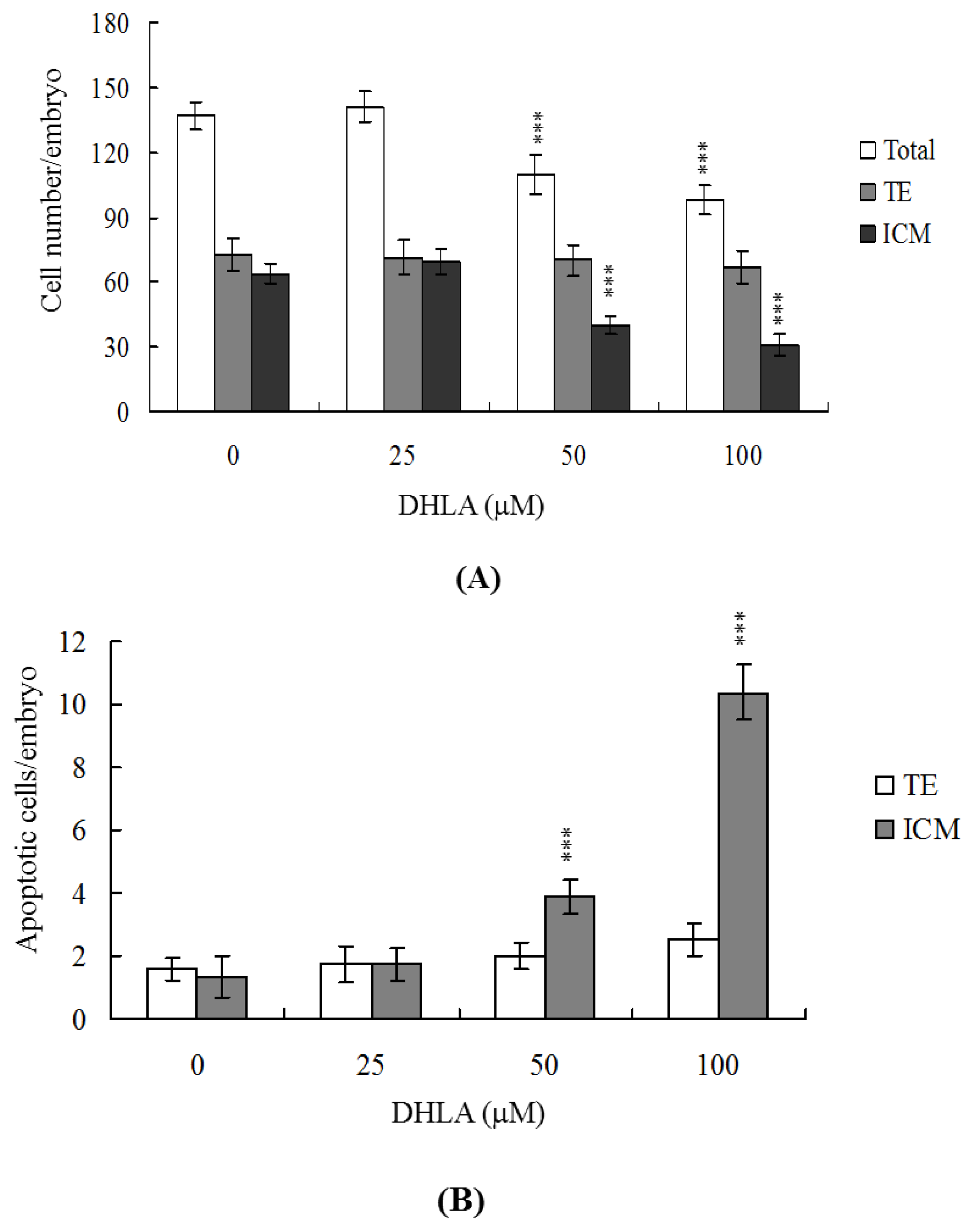
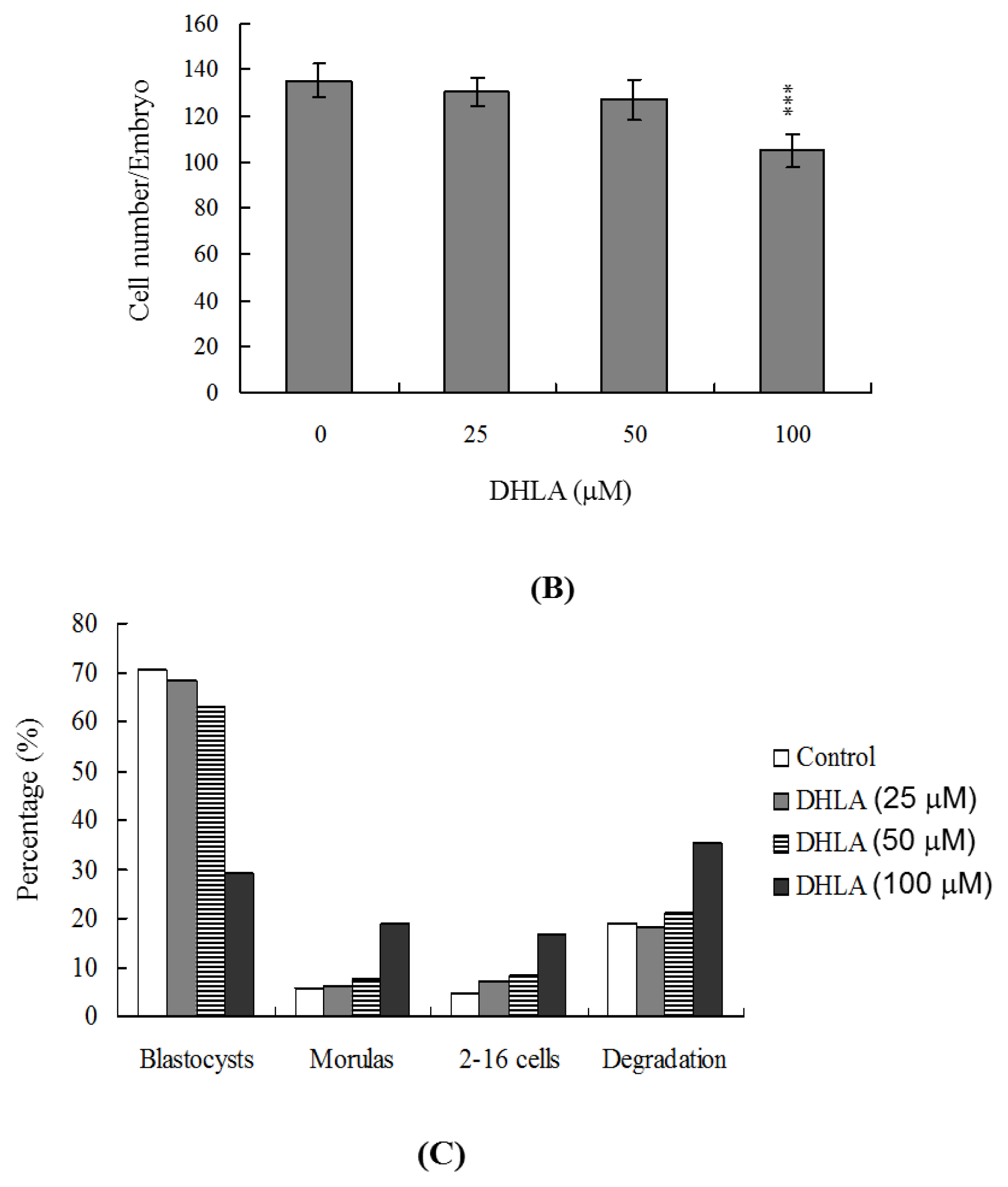
2.1. Effects of DHLA on Mouse Blastocysts
2.2. Effects of DHLA on Cell Proliferation
2.3. Effects of DHLA on Mouse Embryonic Developmental Potential in Vitro
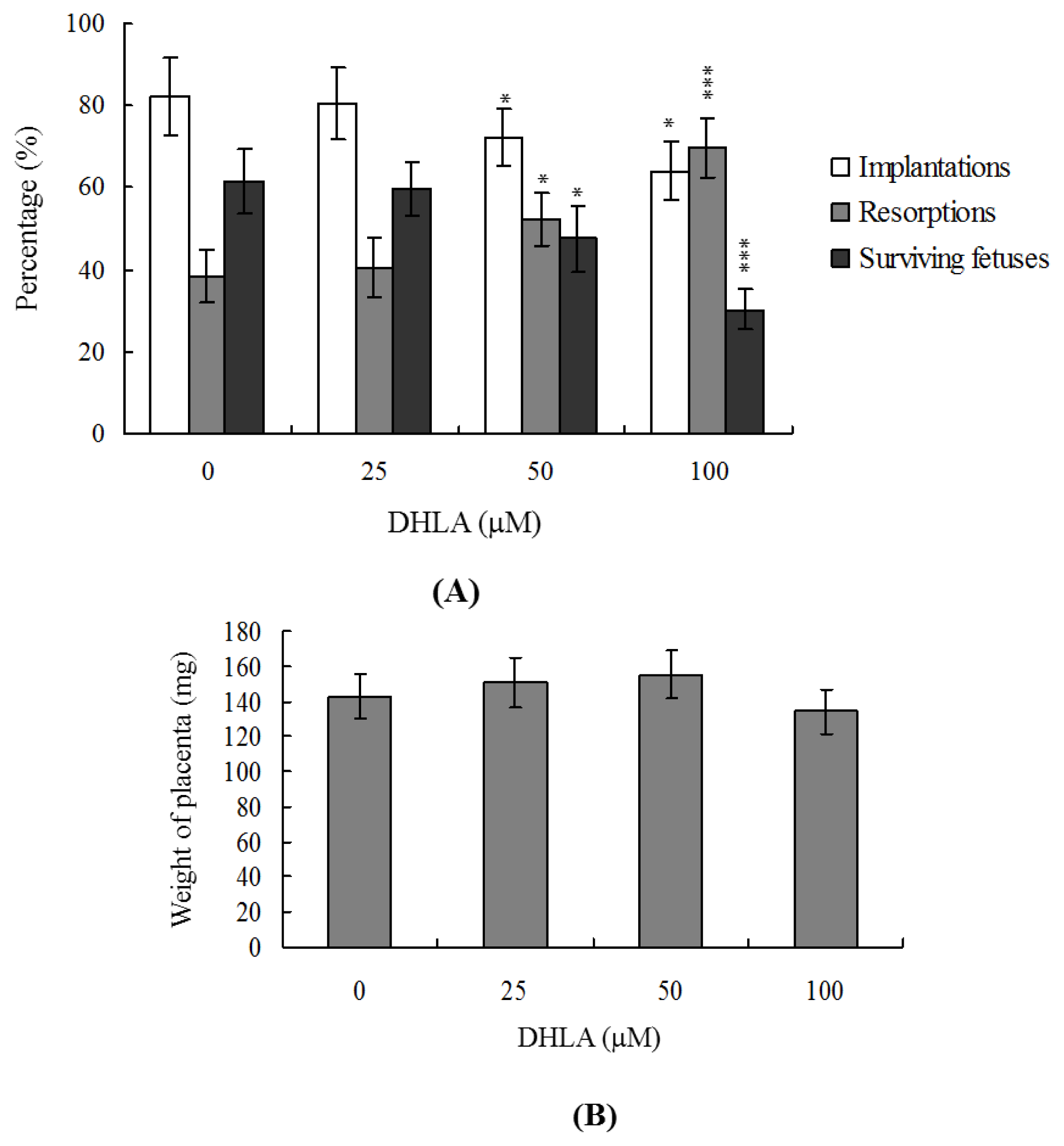
2.4. Effects of DHLA on the Developmental Potential of Blastocysts in Vivo
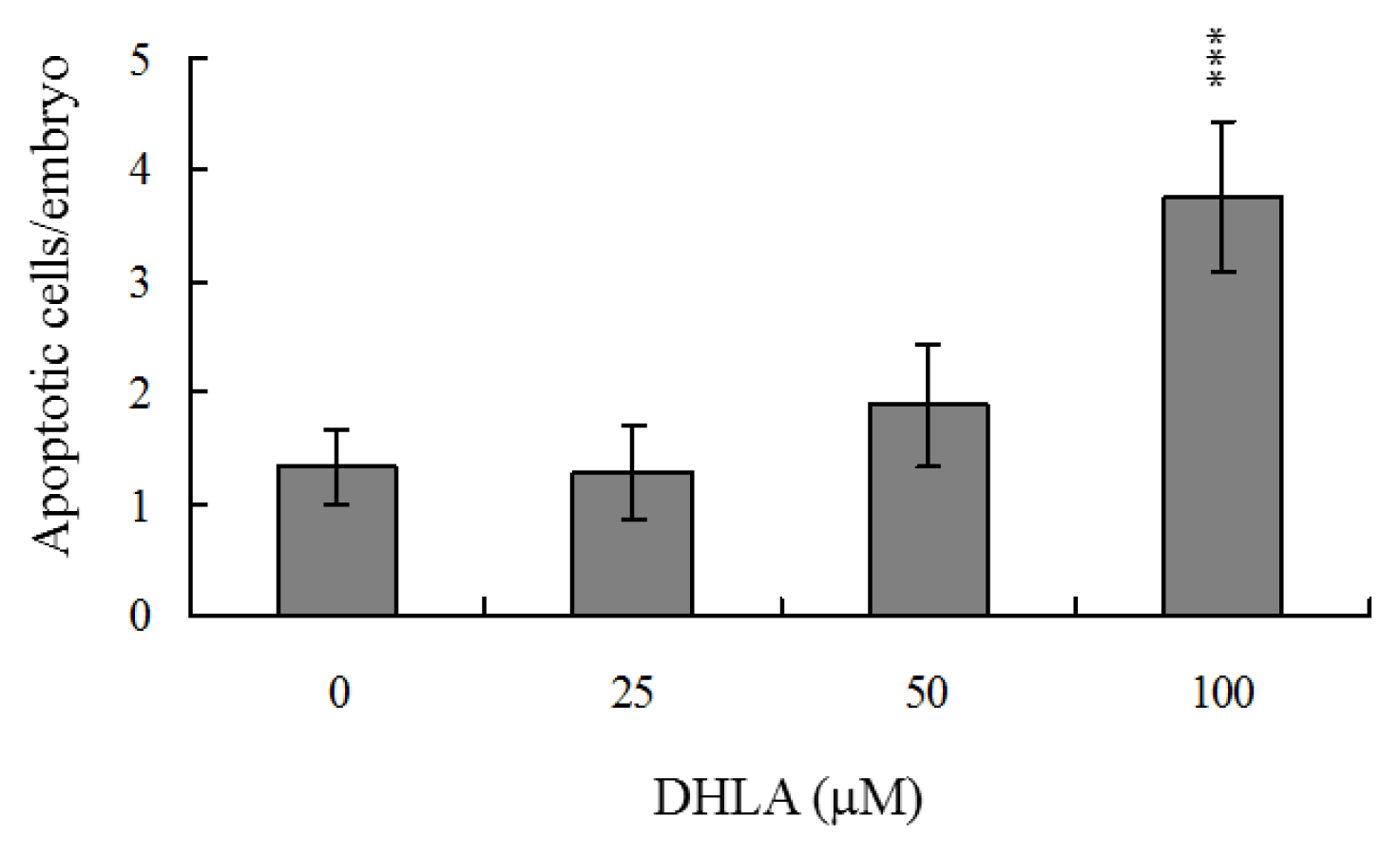
2.5. Disruption of Blastocyst Development by DHLA in Vivo
3. Discussion
4. Experimental Section
4.1. Materials
4.2. Collection of Mouse Morulas and Blastocysts
4.3. DHLA Treatment and TUNEL Assay
4.4. DHLA Treatment and Cell Proliferation
4.5. Annexin V Staining
4.6. Morphological Analysis of Embryonic Development
4.7. Blastocyst Development Following Embryo Transfer
4.8. Statistics
5. Conclusions
Acknowledgements
- Conflicts of interestThe authors declare no conflicts of interest.
References
- Reed, L.J. A trail of research from lipoic acid to alpha-keto acid dehydrogenase complexes. J. Biol. Chem 2001, 276, 38329–38336. [Google Scholar]
- Perham, R.N.; Jones, D.D.; Chauhan, H.J.; Howard, M.J. Substrate channelling in 2-oxo acid dehydrogenase multienzyme complexes. Biochem. Soc. Trans 2002, 30, 47–51. [Google Scholar]
- Bilska, A.; Wlodek, L. Lipoic acid—the drug of the future? Pharmacol. Rep 2005, 57, 570–577. [Google Scholar]
- Packer, L.; Tritschler, H.J.; Wessel, K. Neuroprotection by the metabolic antioxidant alpha-lipoic acid. Free Radic. Biol. Med 1997, 22, 359–378. [Google Scholar]
- Morkunaite-Haimi, S.; Kruglov, A.G.; Teplova, V.V.; Stolze, K.; Gille, L.; Nohl, H.; Saris, N.E. Reactive oxygen species are involved in the stimulation of the mitochondrial permeability transition by dihydrolipoate. Biochem. Pharmacol 2003, 65, 43–49. [Google Scholar]
- Scott, B.C.; Aruoma, O.I.; Evans, P.J.; O’Neill, C.; Van der Vliet, A.; Cross, C.E.; Tritschler, H.; Halliwell, B. Lipoic and dihydrolipoic acids as antioxidants. A critical evaluation. Free Radic. Res 1994, 20, 119–133. [Google Scholar]
- Simbula, G.; Columbano, A.; Ledda-Columbano, G.M.; Sanna, L.; Deidda, M.; Diana, A.; Pibiri, M. Increased ROS generation and p53 activation in alpha-lipoic acid-induced apoptosis of hepatoma cells. Apoptosis 2007, 12, 113–123. [Google Scholar]
- Moungjaroen, J.; Nimmannit, U.; Callery, P.S.; Wang, L.; Azad, N.; Lipipun, V.; Chanvorachote, P.; Rojanasakul, Y. Reactive oxygen species mediate caspase activation and apoptosis induced by lipoic acid in human lung epithelial cancer cells through Bcl-2 down-regulation. J. Pharmacol. Exp. Ther 2006, 319, 1062–1069. [Google Scholar]
- Zheng, Y.; Yamaguchi, H.; Tian, C.; Lee, M.W.; Tang, H.; Wang, H.G.; Chen, Q. Arsenic trioxide (As(2)O(3)) induces apoptosis through activation of Bax in hematopoietic cells. Oncogene 2005, 24, 3339–3347. [Google Scholar]
- Singh, L.P.; Cheng, D.W.; Kowluru, R.; Levi, E.; Jiang, Y. Hexosamine induction of oxidative stress, hypertrophy and laminin expression in renal mesangial cells: Effect of the anti-oxidant alpha-lipoic acid. Cell. Biochem. Funct 2007, 25, 537–550. [Google Scholar]
- Abdul, H.M.; Butterfield, D.A. Involvement of PI3K/PKG/ERK1/2 signaling pathways in cortical neurons to trigger protection by cotreatment of acetyl-l-carnitine and alpha-lipoic acid against HNE-mediated oxidative stress and neurotoxicity: implications for Alzheimer’s disease. Free Radic. Biol. Med 2007, 42, 371–384. [Google Scholar]
- Choi, S.Y.; Yu, J.H.; Kim, H. Mechanism of alpha-lipoic acid-induced apoptosis of lung cancer cells. Ann. NY Acad. Sci 2009, 1171, 149–155. [Google Scholar]
- Chan, W.H.; Houng, W.L.; Lin, C.A.; Lee, C.H.; Li, P.W.; Hsieh, J.T.; Shen, J.L.; Yeh, H.I.; Chang, W.H. Impact of dihydrolipoic acid on mouse embryonic stem cells and related regulatory mechanisms. Environ. Toxicol 2011. [Google Scholar] [CrossRef]
- Liu, W.; Howarth, M.; Greytak, A.B.; Zheng, Y.; Nocera, D.G.; Ting, A.Y.; Bawendi, M.G. Compact biocompatible quantum dots functionalized for cellular imaging. J. Am. Chem. Soc 2008, 130, 1274–1284. [Google Scholar]
- Lin, C.A.; Yang, T.Y.; Lee, C.H.; Huang, S.H.; Sperling, R.A.; Zanella, M.; Li, J.K.; Shen, J.L.; Wang, H.H.; Yeh, H.I.; et al. Synthesis, characterization, and bioconjugation of fluorescent gold nanoclusters toward biological labeling applications. ACS Nano 2009, 3, 395–401. [Google Scholar]
- Lin, C.-A.J.; Lee, C.-H.; Hsieh, J.-T.; Wang, H.-H.; Li, J.K.; Shen, J.-L.; Chan, W.-H.; Yeh, H.-I.; Chang, W.H. Synthesis of fluorescent metallic nanoclusters toward biomedical application: Recent progress and present challenges. J. Med. Biol. Eng 2009, 29, 276–283. [Google Scholar]
- Thompson, C.B. Apoptosis in the pathogenesis and treatment of disease. Science 1995, 267, 1456–1462. [Google Scholar]
- Brill, A.; Torchinsky, A.; Carp, H.; Toder, V. The role of apoptosis in normal and abnormal embryonic development. J. Assist. Reprod. Genet 1999, 16, 512–519. [Google Scholar]
- Lotz, K.; Proff, P.; Bienengraeber, V.; Fanghaenel, J.; Gedrange, T.; Weingaertner, J. Apoptosis as a creative agent of embryonic development of bucca, mentum and nasolacrimal duct. An in vivo study in rats. J. Craniomaxillofac. Surg 2006, 34, 8–13. [Google Scholar]
- Weingaertner, J.; Proff, P.; Bienengraeber, V.; Gedrange, T.; Fanghaenel, J.; Lotz, K. In vivo study of apoptosis as a creative agent of embryonic development of the primary nasal duct in rats. J. Craniomaxillofac. Surg 2006, 34, 3–7. [Google Scholar]
- Huang, F.J.; Shen, C.C.; Chang, S.Y.; Wu, T.C.; Hsuuw, Y.D. Retinoic acid decreases the viability of mouse blastocysts in vitro. Hum. Reprod 2003, 18, 130–136. [Google Scholar]
- Hsuuw, Y.D.; Chang, C.K.; Chan, W.H.; Yu, J.S. Curcumin prevents methylglyoxal-induced oxidative stress and apoptosis in mouse embryonic stem cells and blastocysts. J. Cell. Physiol 2005, 205, 379–386. [Google Scholar]
- Chan, W.H. Ginkgolide B induces apoptosis and developmental injury in mouse embryonic stem cells and blastocysts. Hum. Reprod 2006, 21, 2985–2995. [Google Scholar]
- Shang, E.H.; Wu, R.S. Aquatic hypoxia is a teratogen and affects fish embryonic development. Environ. Sci. Technol 2004, 38, 4763–4767. [Google Scholar]
- Detmar, J.; Rabaglino, T.; Taniuchi, Y.; Oh, J.; Acton, B.M.; Benito, A.; Nunez, G.; Jurisicova, A. Embryonic loss due to exposure to polycyclic aromatic hydrocarbons is mediated by Bax. Apoptosis 2006, 11, 1413–1425. [Google Scholar]
- Selvakumar, E.; Hsieh, T.C. Regulation of cell cycle transition and induction of apoptosis in HL-60 leukemia cells by lipoic acid: role in cancer prevention and therapy. J. Hematol. Oncol 2008, 1, 4. [Google Scholar]
- Huang, F.J.; Hsuuw, Y.D.; Lan, K.C.; Kang, H.Y.; Chang, S.Y.; Hsu, Y.C.; Huang, K.E. Adverse effects of retinoic acid on embryo development and the selective expression of retinoic acid receptors in mouse blastocysts. Hum. Reprod 2006, 21, 202–209. [Google Scholar]
- Chan, W.H. Impact of genistein on maturation of mouse oocytes, fertilization, and fetal development. Reprod. Toxicol 2009, 28, 52–58. [Google Scholar]
- Chan, W.H.; Shiao, N.H. Cytotoxic effect of CdSe quantum dots on mouse embryonic development. Acta Pharmacol. Sin 2008, 29, 259–266. [Google Scholar]
- Chan, W.H.; Shiao, N.H. Effect of citrinin on mouse embryonic development in vitro and in vivo. Reprod. Toxicol 2007, 24, 120–125. [Google Scholar]
- Hardy, K. Cell death in the mammalian blastocyst. Mol. Hum. Reprod 1997, 3, 919–925. [Google Scholar]
- Hardy, K.; Stark, J.; Winston, R.M. Maintenance of the inner cell mass in human blastocysts from fragmented embryos. Biol. Reprod 2003, 68, 1165–1169. [Google Scholar]
- Byrne, A.T.; Southgate, J.; Brison, D.R.; Leese, H.J. Analysis of apoptosis in the preimplantation bovine embryo using TUNEL. J. Reprod. Fertil 1999, 117, 97–105. [Google Scholar]
- Chan, W.H. Effects of citrinin on maturation of mouse oocytes, fertilization, and fetal development in vitro and in vivo. Toxicol. Lett 2008, 180, 28–32. [Google Scholar]
- Chan, W.H. Ginkgolides induce apoptosis and decrease cell numbers in mouse blastocysts. Biochem. Biophys. Res. Commun 2005, 338, 1263–1267. [Google Scholar]
- Cross, J.C.; Werb, Z.; Fisher, S.J. Implantation and the placenta: Key pieces of the development puzzle. Science 1994, 266, 1508–1518. [Google Scholar]
- Pampfer, S.; de Hertogh, R.; Vanderheyden, I.; Michiels, B.; Vercheval, M. Decreased inner cell mass proportion in blastocysts from diabetic rats. Diabetes 1990, 39, 471–476. [Google Scholar]
- Kelly, S.M.; Robaire, B.; Hales, B.F. Paternal cyclophosphamide treatment causes postimplantation loss via inner cell mass-specific cell death. Teratology 1992, 45, 313–318. [Google Scholar]
- Tam, P.P. Postimplantation development of mitomycin C-treated mouse blastocysts. Teratology 1988, 37, 205–212. [Google Scholar]
- Hardy, K.; Handyside, A.H.; Winston, R.M. The human blastocyst: Cell number, death and allocation during late preimplantation development in vitro. Development 1989, 107, 597–604. [Google Scholar]
- Gardner, R.L.; Davies, T.J. Lack of coupling between onset of giant transformation and genome endoreduplication in the mural trophectoderm of the mouse blastocyst. J. Exp. Zool 1993, 265, 54–60. [Google Scholar]
- Huang, F.J.; Wu, T.C.; Tsai, M.Y. Effect of retinoic acid on implantation and post-implantation development of mouse embryos in vitro. Hum. Reprod 2001, 16, 2171–2176. [Google Scholar]
- Witschi, E. Characterization of Developmental Stages. Part II. Rat. In Biology Data Book, 2nd ed; Federation of American Societies of Experimental Biologies: Washington DC, USA, 1972; pp. 178–180. [Google Scholar]
- Armant, D.R.; Kaplan, H.A.; Lennarz, W.J. Fibronectin and laminin promote in vitro attachment and outgrowth of mouse blastocysts. Dev. Biol 1986, 116, 519–523. [Google Scholar]
- Pampfer, S.; Wuu, Y.D.; Vanderheyden, I.; de Hertogh, R. In vitro study of the carry-over effect associated with early diabetic embryopathy in the rat. Diabetologia 1994, 37, 855–862. [Google Scholar]







© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Houng, W.-L.; Lin, C.-A.J.; Shen, J.-L.; Yeh, H.-I.; Wang, H.-H.; Chang, W.H.; Chan, W.-H. Dihydrolipoic Acid Induces Cytotoxicity in Mouse Blastocysts through Apoptosis Processes. Int. J. Mol. Sci. 2012, 13, 3988-4002. https://doi.org/10.3390/ijms13033988
Houng W-L, Lin C-AJ, Shen J-L, Yeh H-I, Wang H-H, Chang WH, Chan W-H. Dihydrolipoic Acid Induces Cytotoxicity in Mouse Blastocysts through Apoptosis Processes. International Journal of Molecular Sciences. 2012; 13(3):3988-4002. https://doi.org/10.3390/ijms13033988
Chicago/Turabian StyleHoung, Wei-Li, Cheng-An J. Lin, Ji-Lin Shen, Hung-I Yeh, Hsueh-Hsiao Wang, Walter H. Chang, and Wen-Hsiung Chan. 2012. "Dihydrolipoic Acid Induces Cytotoxicity in Mouse Blastocysts through Apoptosis Processes" International Journal of Molecular Sciences 13, no. 3: 3988-4002. https://doi.org/10.3390/ijms13033988