Phylogenetic Relationships of the Mangalitsa Swine Breed Inferred from Mitochondrial DNA Variation


Abstract
:1. Introduction
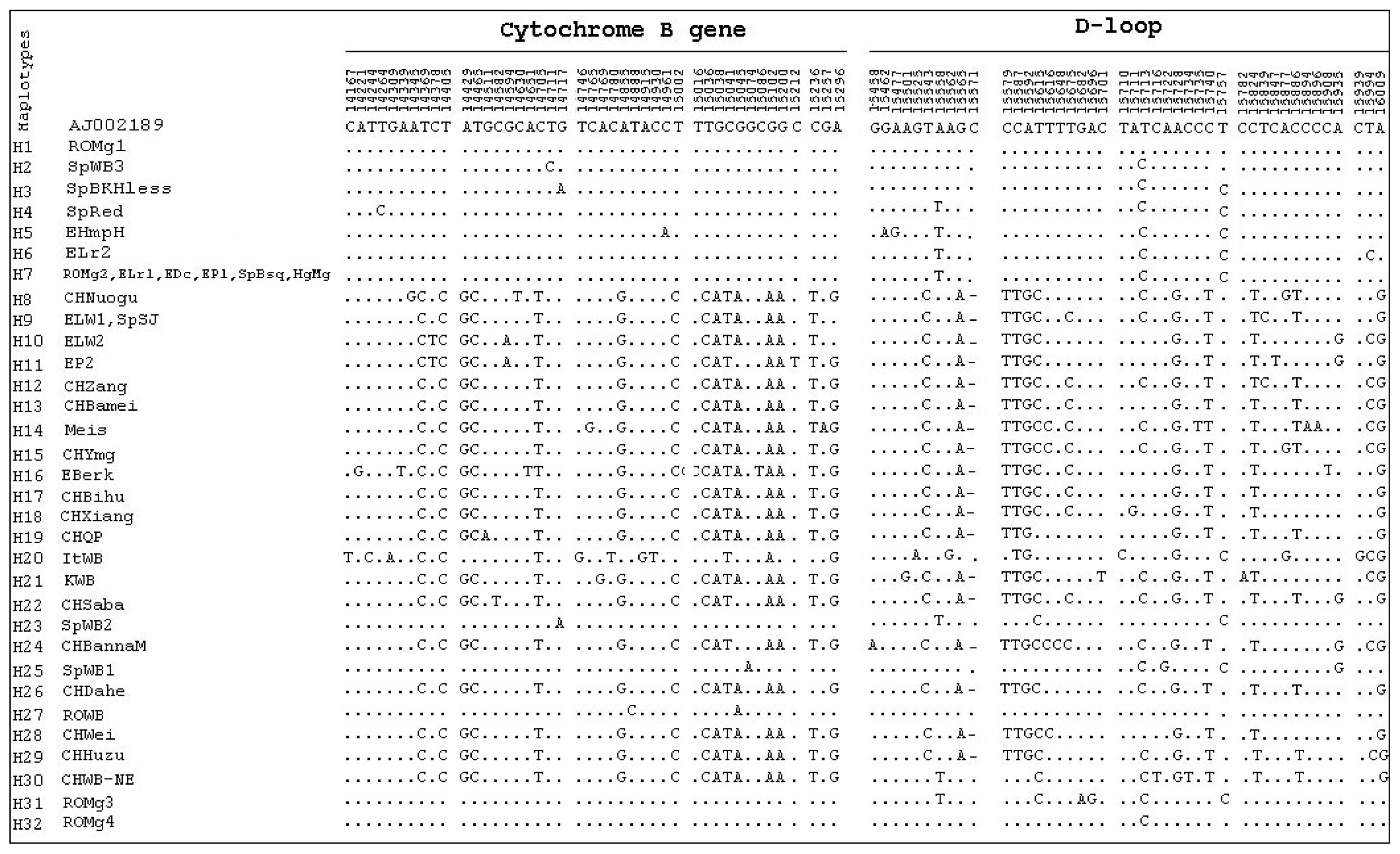
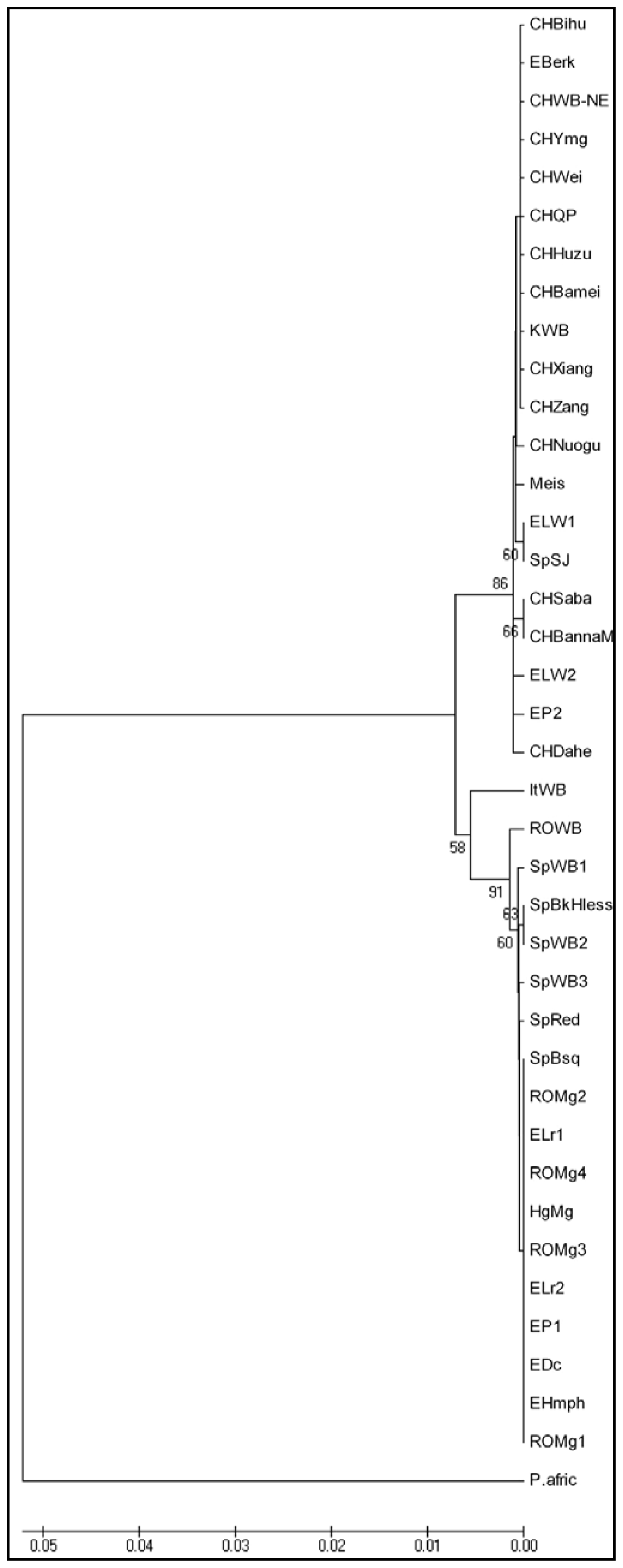
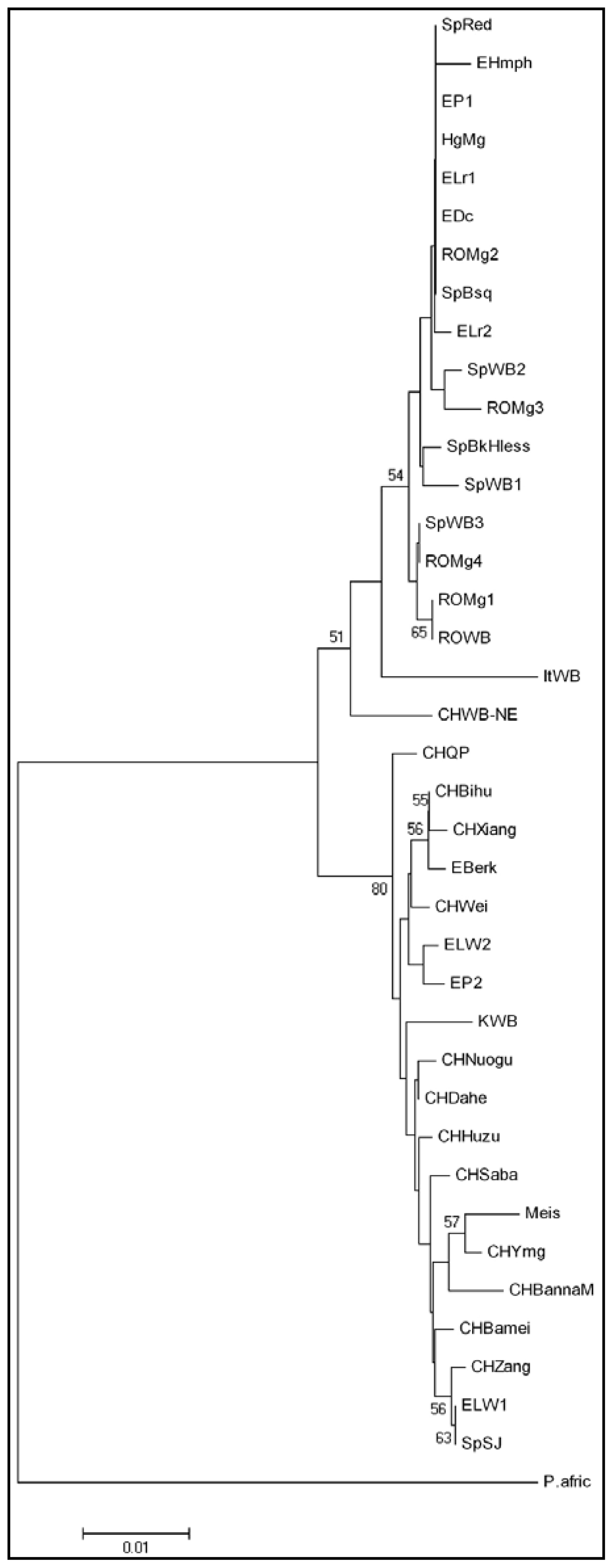
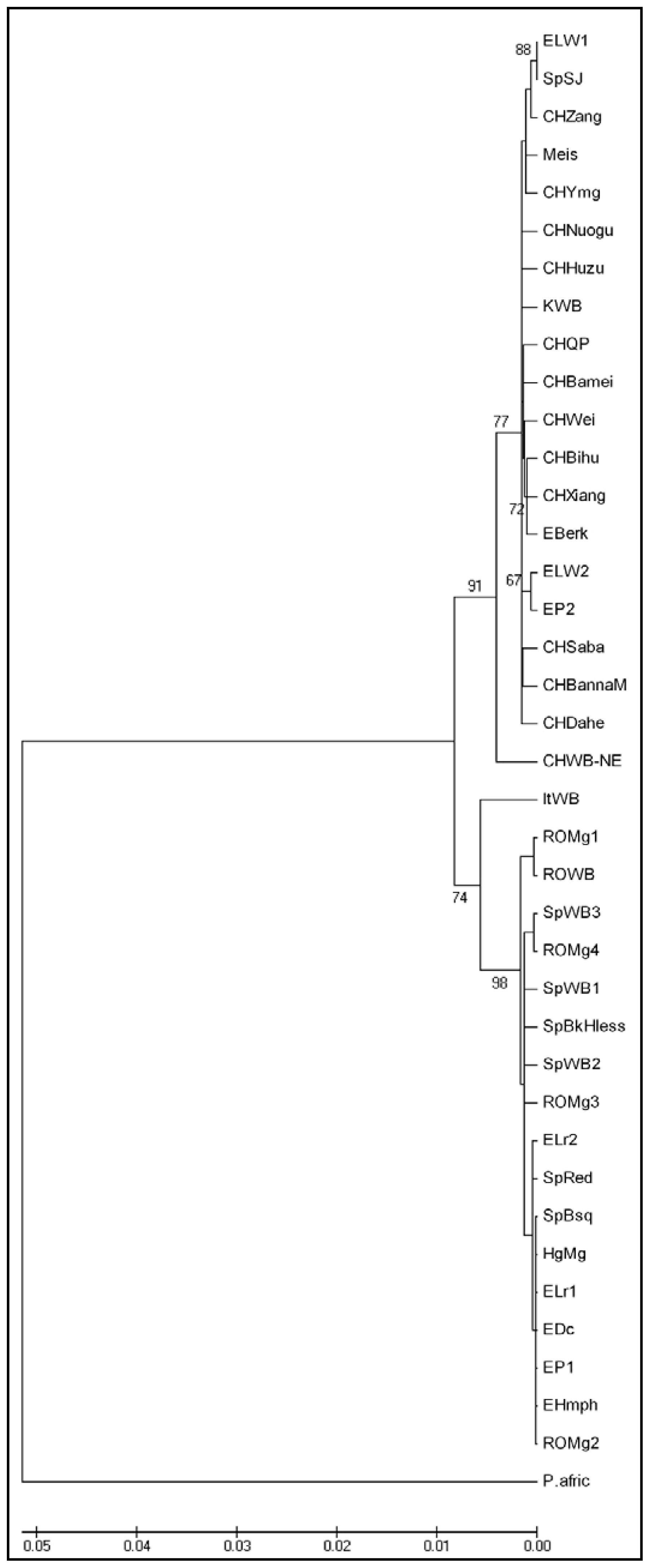
2. Results and Discussion
3. Experimental Section
3.1. Sampling and DNA Extraction
3.2. DNA Amplification and Sequencing
3.3. Sequence Alignment and Molecular Phylogenetic Analysis
4. Conclusions
References
- Luís, C.; Bastos-Silveira, C.; Cothran, E.G.; Om, M. Variation in the mitochondrial control region sequence between the two maternal lines of the Sorraia horse breed. Genet. Mol. Biol 2002, 25, 309–311. [Google Scholar]
- Aberle, K.; Hamann, H.; Drögemüller, C.; Distl, O. Genetic diversity in german draught horse breeds compared with a group of primitive, riding and wild horses by means of microsatellite DNA markers. Anim. Genet 2004, 35, 270–277. [Google Scholar]
- Georgescu, S.E.; Manea, M.A.; Dudu, A.; Costache, M. Phylogenetic relationships of the Hucul horse from Romania inferred from mitochondrial D-loop variation. Genet. Mol. Res 2011, 10, 4104–4113. [Google Scholar]
- Kim, K.; Yeo, J.; Choi, C. Genetic diversity of north-east Asian cattle based on microsatellite data. Anim. Genet 2002, 33, 201–204. [Google Scholar]
- Lai, S.; Liu, Y.; Li, X.; Yao, Y. Genetic diversity and origin of Chinese cattle revealed by mtDNA D-loop sequence variation. Mol. Phylogenet. Evol 2006, 38, 146–154. [Google Scholar]
- Li, K.; Chen, Y.; Moran, C.; Fan, B.; Zhao, S.; Peng, Z. Analysis of diversity and genetic relationships between four Chinese indigenous pig breeds and one Australian commercial pig breed. Anim. Genet 2000, 31, 322–325. [Google Scholar]
- Kim, K.I.; Lee, J.H.; Li, K.; Zhang, Y.P.; Lee, S.S.; Gongora, J.; Moran, C. Phylogenetic relationships of Asian and European pig breeds determined by mitochondrial DNA D-loop sequence polymorphism. Anim. Genet 2002, 33, 19–25. [Google Scholar]
- Fabuel, E.; Barragan, C.; Silio, L.; Rodriguez, M.C.; Toro, M.A. Analysis of genetic diversity and conservation priorities in Iberian pigs based on microsatellite markers. Heredity 2004, 93, 104–113. [Google Scholar]
- Fang, M.; Hu, X.; Jiang, T.; Braunschweig, M.; Hu, L.; Du, Z.; Feng, J.; Zhang, Q.; Wu, C.; Li, N. The phylogeny of Chinese indigenous pig breeds inferred from microsatellite markers. Anim. Genet 2005, 36, 7–13. [Google Scholar]
- Fang, M.; Andersson, L. Mitochondrial diversity in European and Chinese pigs is consistent with population expansions that occurred prior to domestication. Proc. Biol. Sci 2006, 273, 1803–1810. [Google Scholar]
- Pedrosa, S.; Uzun, M.; Arranz, J.; Gutierrez-Gil, B.; Primitivo, F.; Bayon, Y. Evidence of three maternal lineages in near eastern sheep supporting multiple domestication events. Proc. R. Soc. Lond 2005, B272, 2211–2217. [Google Scholar]
- Peter, C.; Brufford, M.; Perez, T.; Dalamitra, S.; Hewitt, G.; Erhardt, G. Econogene consortium, genetic diversity and subdivision of 57 European and Middle-Eastern sheep breeds. Anim. Genet 2007, 38, 37–44. [Google Scholar]
- Radnóczi, L. The hungarian mangalica; Agroservice AgriBusiness Club, 2003. Web site. Available online: http://www.agroservice.hu/mangainfo1.htm accessed on 30 May 2012.
- Ciobanu, D.C.; Day, A.E.; Nagy, A.; Wales, R.; Rothschild, M.F.; Plastow, G.S. Genetic variation in two conserved local Romanian pig breeds using type 1 DNA markers. Genet. Sel. Evol 2001, 33, 417–432. [Google Scholar]
- Ursing, B.M.; Arnason, U. The complete mitochondrial DNA sequence of the pig (Sus scrofa). J. Mol. Evol 1998, 47, 302–306. [Google Scholar]
- Kimura, M. A simple method for estimating evolutionary rate of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol 1980, 16, 111–120. [Google Scholar]
- Giuffra, E.; Kijas, J.M.; Amarger, V.; Carlborg, O.; Jeon, J.T.; Andersson, L. The origin of the domestic pig: Independent domestication and subsequent introgression. Genetics 2000, 154, 1785–1791. [Google Scholar]
- Alves, E.; Ovilo, C.; Rodriguez, M.C.; Silio, L. Mitochondrial DNA sequence variation and phylogenetic relationships among Iberian pigs and other domestic and wild pig populations. Anim. Genet 2003, 34, 319–324. [Google Scholar]
- Jones, G.F. Genetic Aspects of Domestication, Common Breeds and Their Origins. In The Genetics of the Pig; Rothschild, M.F., Ruvinsky, A., Eds.; CAB International: Wallingford, Oxon, UK, 1998; pp. 17–50. [Google Scholar]
- Epstein, J.; Bichard, M. Pig. In Evolution of Domesticated Animals; Mason, I.L., Ed.; John Wiley & Sons: New York, NY, USA, 1986; pp. 145–162. [Google Scholar]
- Watanobe, T.; Ishiguro, N.; Nakano, M.; Matsui, A.; Hongo, H.; Yamazaki, K.; Takahashi, O. Prehistoric Sado Island populations of Sus scrofa distinguished from contemporany Japanese Wild boar by ancient mitochondrial DNA. Zoolog. Sci 2004, 21, 219–228. [Google Scholar]
- Kijas, J.M.; Andersson, L. A phylogenetic study of the origin of the domestic pig estimated from the near-complete mtDNA genome. J. Mol. Evol 2001, 52, 302–308. [Google Scholar]
- Li, W.H. Molecular Evolution; Sinauer Associates: Sunderland, MA, USA, 1997; p. 432. [Google Scholar]
- Pesole, G.; Gissi, C.; de Chirico, A.; Saccone, C. Nucleotide substitution rate of mammalian mitochondrial genomes. J. Mol. Evol 1999, 48, 427–434. [Google Scholar]
- Sbisà, E.; Tanzariello, F.; Reyes, A.; Pesole, G.; Saccone, C. Mammalian mitochondrial D-loop region structural analisis: Identification of new conserved sequences and their functional and evolutionary implications. Genetics 1997, 205, 125–140. [Google Scholar]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl. Acids Symp. Ser 1999, 41, 95–98. [Google Scholar]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar]
- Posada, D.; Crandall, K.A. Modeltest: Testing the model of DNA substitution. Bioinformatics 1998, 14, 817–818. [Google Scholar]
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA4: Molecular evolutionary genetics analysis (MEGA) software version 4.0. Mol. Biol. Evol 2007, 24, 1596–1599. [Google Scholar]
- Wang, J.F.; Li, S.; Ran, X.Q. Department of Animal Science, Guizhou University, Xueshi Road, Huaxi, Guiyang, Guizhou Province 550025, ChinaSus scrofa breed Nuogu mitochondrion, complete genome. In Unpublished work; 2006. [Google Scholar]
- Su, X.X.; Cheng, W.M.; Zeng, Y.Z. Key Laboratory of Banna Mini-Pig Inbred Line of Yunnan Province, Yunnan Agricultural University: Longquan Load, Kunming, Yunnan 650201, China; Complete mitochondrial genome of Banan mini-pig inbred line. In Unpublished work; 2009. [Google Scholar]
- Su, X.X.; Cheng, W.M.; Zeng, Y.Z. Key Laboratory of Banna Mini-Pig Inbred Line of Yunnan Province, Yunnan Agricultural University: Longquan Load, Kunming, Yunnan 650201, China; Complete sequence and gene organization of the Dahe pig mitochondrial genome. In Unpublished work; 2009. [Google Scholar]
- Wu, G.S.; Yao, Y.G.; Qu, K.X.; Ding, Z.L.; Li, H.; Palanichamy, M.G.; Duan, Z.Y.; Li, N.; Chen, Y.S.; Zhang, Y.P. Population phylogenomic analysis of mitochondrial DNA in wild boars and domestic pigs revealed multiple domestication events in East Asia. Genome Biol 2007, 8, R245. [Google Scholar]
- Cho, I.C.; Han, S.H.; Lee, S.S.; Ko, M.S.; Lee, J.G.; Jeon, J.T. Division of Livestock, National Institute of Subtropical Agriculture: R.D.A., 175-6, O-deung dong, Jeju, 690-150, South Korea; The complete mitochondrial DNA sequence of Korean wild boar. In Unpublished work; 2007. [Google Scholar]
- Yu, H.; Li, L.; Liu, D. Animal Science and Technology College, Northeast Agricultural University: 59 Mucai Street, Harbin, HeiLongjiang 150030, China; Phylogeography of Boar based on mtDNA evolution. In Unpublished work; 2007. [Google Scholar]
- Cho, I.C.; Park, J.J.; Jeon, J.T. National Institute of Subtropical Agriculture: R.D.A., 1696, Odeung dong, Jeju 690-150, South Korea; The complete sequence of mitochondrial DNA of Berkshire (Sus scrofa). In Unpublished work; 2004. [Google Scholar]
- Cho, I.C.; Park, J.J.; Jeon, J.T. National Institute of Subtropical Agriculture: R.D.A., 1696, Odeung dong, Jeju 690-150, South Korea; The complete sequence of mitochondrial DNA of Hampshire (Sus scrofa). In Unpublished work; 2004. [Google Scholar]
- Larson, G.; Dobney, K.; Albarella, U.; Fang, M.; Matisoo-Smith, E.; Robins, J.; Lowden, S.; Finlayson, H.; Brand, T.; Willerslev, E.; et al. Worldwide phylogeography of wild boar reveals multiple centers of pig domestication. Science 2005, 307, 1618–1621. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Breed name | Polymorphic sites | Haplotypes | Transitions | Transversions | Deletions |
|---|---|---|---|---|---|
| European breeds | |||||
| Mangalitsa (Romania) | 9 | 4 | 8 | 1 | - |
| Mangalitsa (Hungary) | 3 | 1 | 2 | 1 | - |
| Black Hairless | 3 | 1 | 3 | - | - |
| Red | 4 | 1 | 3 | 1 | - |
| Duroc | 3 | 1 | 2 | 1 | - |
| Large White | 33 | 2 | 32 | - | 1 |
| Landrace | 4 | 2 | 4 | - | - |
| Pietran | 32 | 2 | 31 | - | 1 |
| Hampshire | 6 | 1 | 4 | 2 | - |
| Spotted Black Jabugo | 29 | 1 | 28 | - | 1 |
| Basque | 3 | 1 | 2 | 1 | - |
| Berkshire | 33 | 1 | 30 | 2 | 1 |
| Asiatic breeds | |||||
| Nuogu | 31 | 1 | 30 | - | 1 |
| Banna Mini | 32 | 1 | 31 | - | 1 |
| Dahe | 26 | 1 | 25 | - | 1 |
| Saba | 30 | 1 | 29 | - | 1 |
| Zang | 31 | 1 | 30 | - | 1 |
| Wei | 27 | 1 | 26 | - | 1 |
| Qing Ping | 27 | 1 | 26 | - | 1 |
| Bamei | 29 | 1 | 28 | - | 1 |
| Huzu | 29 | 1 | 28 | - | 1 |
| Yimenghei | 32 | 1 | 31 | - | 1 |
| Bihu | 28 | 1 | 27 | - | 1 |
| Xiang | 28 | 1 | 27 | - | 1 |
| Meishan | 36 | 1 | 32 | 3 | 1 |
| Asiatic and European Wild Boars | |||||
| Wild Boar (N-E China) | 26 | 1 | 25 | - | 1 |
| Wild Boar (Koreea) | 32 | 1 | 30 | 1 | 1 |
| Wild Boar (Italy) | 24 | 1 | 22 | 2 | - |
| Wild Boar (Romania) | 2 | 1 | 2 | - | - |
| Wild Boar (Spain) | 11 | 3 | 10 | 1 | - |
| mtDNA region | Nucleotide substitution rate (%) [24] | K ± SD (%) | T (× 103 years) |
|---|---|---|---|
| Cytochrome b gene | |||
| Synonymous positions | 27.4 ± 3.3 | 3.875 ± 0.011 | 707 |
| Non-synonymous positions | 1.8 ± 0.3 | 0.154 ± 0.001 | 428 |
| D-loop region | |||
| ETAS Domain | 19.4 ± 7.8 | 2.273 ± 0.006 | 586 |
| Central Domain | 3.8 ± 1.9 | 0.843 ± 0.004 | 1109 |
| Average (including Central Domain) | 707 | ||
| Average (without Central Domain) | 574 | ||
| Abbreviation | Accession number | Breed/Location | References |
|---|---|---|---|
| SpBkHless | AY237494/cytb AY232852/D-loop | Black Hairless/Spain/Europe | Alves et al., 2003 [18] |
| SpRed | AY237498/cytb AY232856/D-loop | Red/Spain/Europe | Alves et al., 2003 [18] |
| SpWB1 | AY237510/cytb AY232868/D-loop | Wild Boar 1/Spain/Europe | Alves et al., 2003 [18] |
| SpWB2 | AY237513/cytb AY232871/D-loop | Wild Boar 2/Spain/Europe | Alves et al., 2003 [18] |
| SpWB3 | AY237515/cytb AY232873/D-loop | Wild Boar 3/Spain/Europe | Alves et al., 2003 [18] |
| EDc | AY237519/cytb AY232877/D-loop | Duroc/Europe | Alves et al., 2003 [18] |
| ELW1 | AY237524/cytb AY232882/D-loop | Large White 1/Europe | Alves et al., 2003 [18] |
| ELW2 | AY237525/cytb AY232883/D-loop | Large White 2/Europe | Alves et al., 2003 [18] |
| ELr1 | AY237526/cytb AY232884/D-loop | Landrace 1/Europe | Alves et al., 2003 [18] |
| ELr2 | AY237527/cytb AY232885/D-loop | Landrace 2/Europe | Alves et al., 2003 [18] |
| EP1 | AY237528/citb AY232886/D-loop | Pietran 1/Europe | Alves et al., 2003 [18] |
| EP2 | AY237529/cytb AY232887/D-loop | Pietran 2/Europe | Alves et al., 2003 [18] |
| Meis | AY237530/cytb AY232888/D-loop | Meishan/China | Alves et al., 2003 [18] |
| SpSJ | AY237532/cytb AY232890/D-loop | Spotted Black Jabugo/Spain/Europe | Alves et al., 2003 [18] |
| SpBsq | AY237533/cytb AY232891/D-loop | Basque/Spain/Europe | Alves et al., 2003 [18] |
| HgMg | AY237534/cytb AY232892/D-loop | Mangalitsa/Hungary/urope | Alves et al., 2003 [18] |
| CHNuogu | DQ466081 | Nuogu/China | Wang et al., 2006 [31] |
| CHBannaM | GQ220328 | Banna Mini/China | Su et al., 2009 [32] |
| CHDahe | GQ220329 | Dahe/China | Su et al., 2009 [33] |
| CHSaba | EF545567 | Saba/China | Wu et al., 2007 [34] |
| CHZang | EF545576 | Zang/China | Wu et al., 2007 [34] |
| CHWei | EF545577 | Wei/China | Wu et al., 2007 [34] |
| CHQP | EF545582 | Qing Ping/China | Wu et al., 2007 [34] |
| CHBamei | EF545583 | Bamei/China | Wu et al., 2007 [34] |
| CHHuzu | EF545588 | Huzu/China | Wu et al., 2007 [34] |
| CHYmg | EF545589 | Yimenghei/China | Wu et al., 2007 [34] |
| CHBihu | EF545591 | Bihu/China | Wu et al., 2007 [34] |
| CHXiang | EF545593 | Xiang/China | Wu et al., 2007 [34] |
| KWB | EU090703 | Wild Boar/Korea | Cho et al., 2007 [35] |
| CHWB-NE | EU333163 | Wild Boar/Nord-East China | Yu et al., 2007 [36] |
| Eberk | AY574045 | Berkshire/Europe | Cho et al., 2004 [37] |
| EHmph | AY574046 | Hampshire/Europe | Cho et al., 2004 [38] |
| ItWB | AF304201 | Wild Boar/Italy/Europe | Kijas and Andersson, 2001 [22] |
| P. afric | NC_008830 | Phacochoerus africanus | Wu et al., 2007 [34] |
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Georgescu, S.E.; Manea, M.A.; Dudu, A.; Costache, M. Phylogenetic Relationships of the Mangalitsa Swine Breed Inferred from Mitochondrial DNA Variation. Int. J. Mol. Sci. 2012, 13, 8467-8481. https://doi.org/10.3390/ijms13078467
Georgescu SE, Manea MA, Dudu A, Costache M. Phylogenetic Relationships of the Mangalitsa Swine Breed Inferred from Mitochondrial DNA Variation. International Journal of Molecular Sciences. 2012; 13(7):8467-8481. https://doi.org/10.3390/ijms13078467
Chicago/Turabian StyleGeorgescu, Sergiu Emil, Maria Adina Manea, Andreea Dudu, and Marieta Costache. 2012. "Phylogenetic Relationships of the Mangalitsa Swine Breed Inferred from Mitochondrial DNA Variation" International Journal of Molecular Sciences 13, no. 7: 8467-8481. https://doi.org/10.3390/ijms13078467