Effects of Brown Rice and White Rice on Expression of Xenobiotic Metabolism Genes in Type 2 Diabetic Rats

Abstract
:1. Introduction
2. Results and Discussion
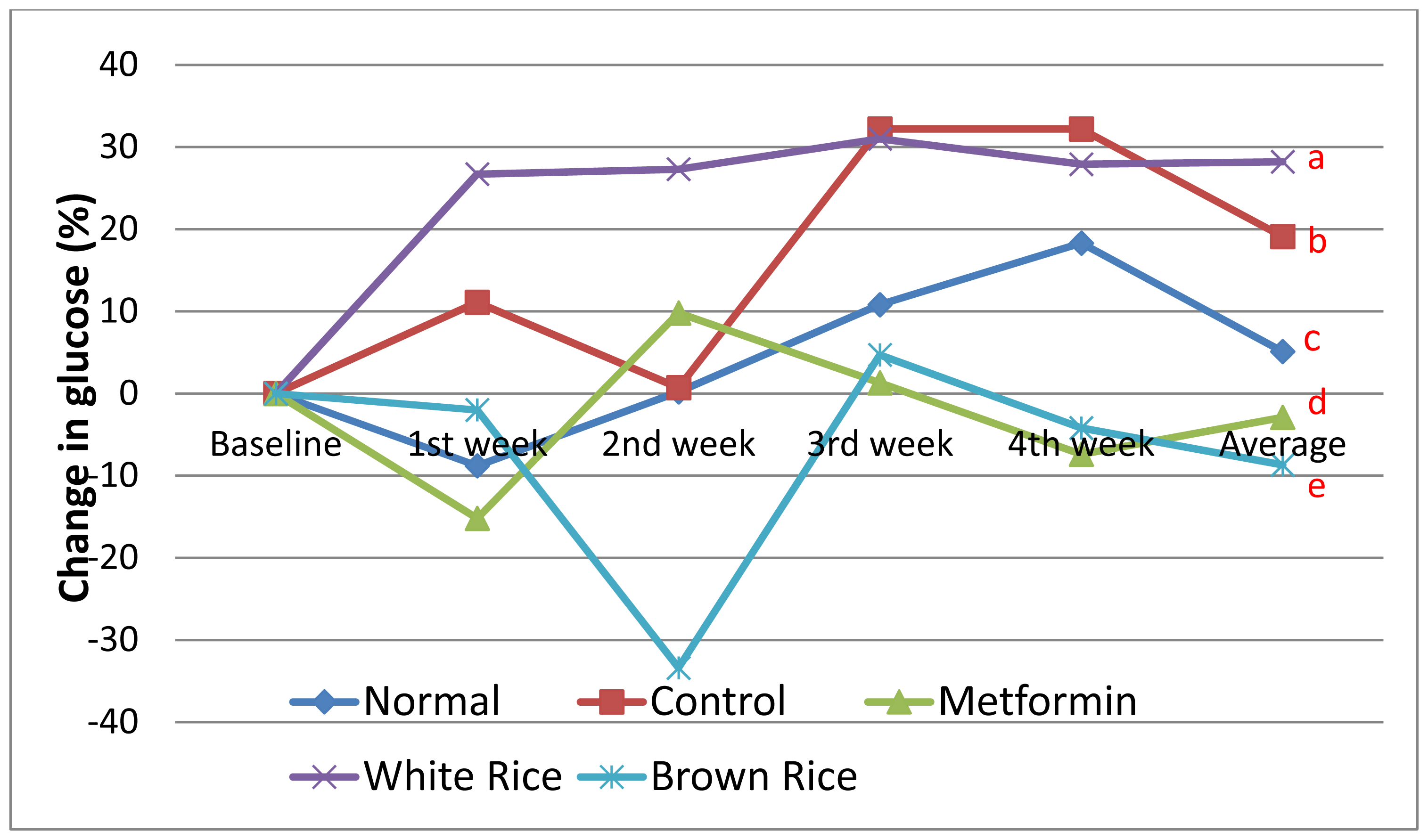
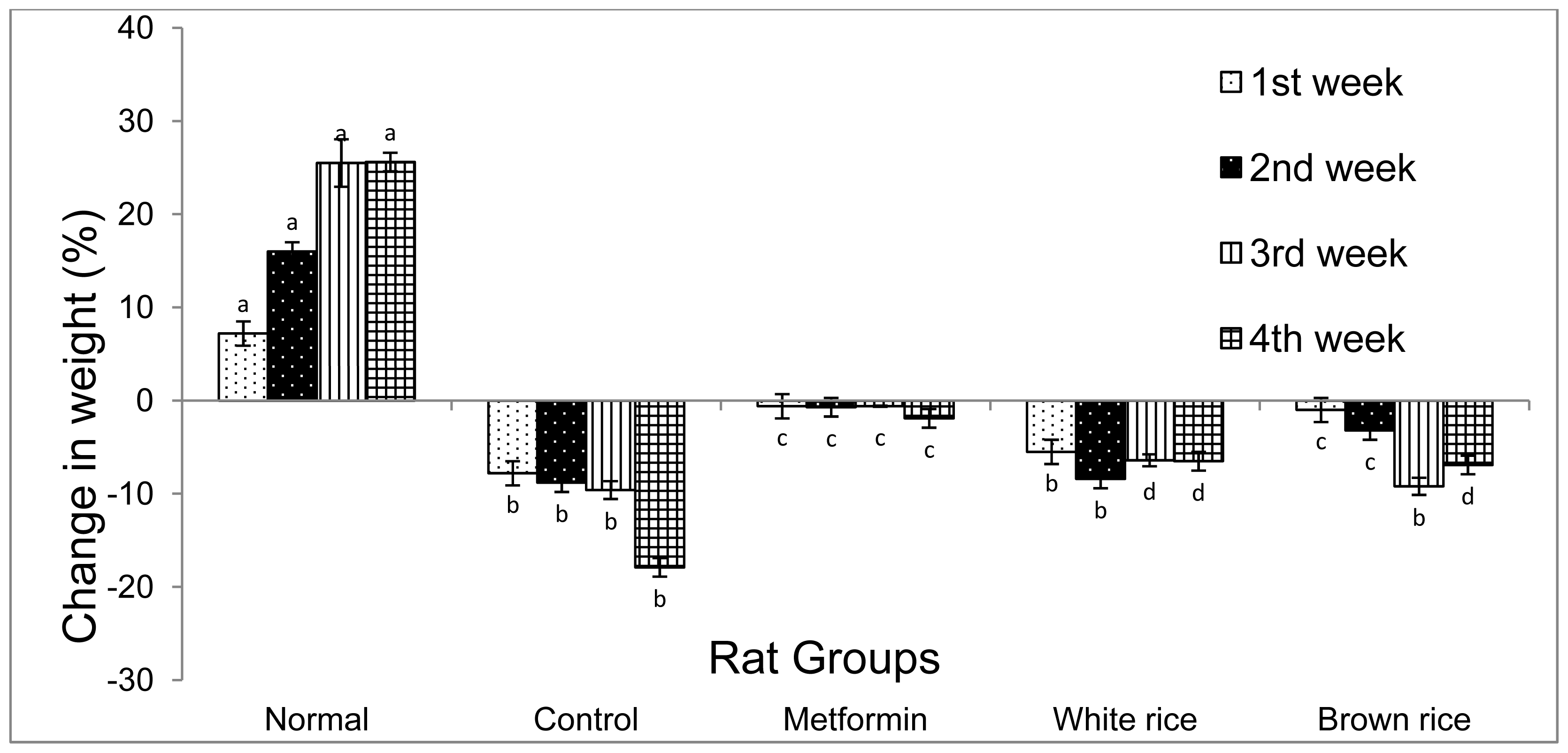
2.1. Blood Glucose and Weight
2.2. Effect of Brown Rice on Expression of Hepatic Xenobiotic Metabolism Genes
3. Experimental Section
3.1. Chemicals
3.2. Rice Grain
3.3. Animal Handling, Feeding and Induction of Diabetes
3.4. Glucose Analysis
3.5. RNA Isolation
3.6. cDNA Synthesis and PCR Amplification
3.7. GeXP Multiplex Data Analysis
3.8. Fragment Analysis and Gene Expression Signature Analysis
4. Conclusions
Acknowledgments
References
- Xu, C.; Li, C.Y.T.; Kong, A.N.T. Induction of phase I, II and III drug metabolism/transport by xenobiotics. Arch. Pharm. Res 2005, 28, 249–268. [Google Scholar]
- World Health Organization. Diabetes fact sheet. Available online: http://www.who.int/mediacentre/factsheets/fs312/en/index.html accessed on 2 June 2012.
- Stumvoll, M.; Goldstein, B.J.; van Haeften, T.W. Type 2 diabetes: Principles of pathogenesis and therapy. Lancet 2005, 365, 1333–1346. [Google Scholar]
- Lewis, D.F. 57 varieties: The human cytochromes P450. Pharmacogenomics 2004, 5, 305–318. [Google Scholar]
- Barnett, C.R.; Flatt, P.R.; Ioannides, C. Modulation of the rat hepatic cytochrome P450 composition by long-term streptozotocin-induced insulin-dependent diabetes. J. Biochem. Toxicol 1994, 2, 63–69. [Google Scholar]
- Nobuo, S. Cytochrome P450 changes in rats with streptozocin-induced diabetes. Int. J. Biochem 1994, 26, 1261–1268. [Google Scholar]
- Sakuma, T.; Honma, R.; Maguchi, S.; Tamaki, H.; Nemoto, N. Different expression of hepatic and renal cytochrome P450s between the streptozotocininduced diabetic mouse and rat. Xenobiotica 2001, 4, 223–237. [Google Scholar]
- Shimojo, N.; Ishizaki, T.; Imaoka, S.; Funae, Y.; Fuji, S.; Okuda, K. Changes in amounts of cytochrome P450 isozymes and levels of catalytic activities in hepatic and renal microsomes of rats with streptozocin-induced diabetes. Biochem. Pharmacol 1993, 46, 621–627. [Google Scholar]
- Wang, Z.; Hall, S.D.; Maya, J.F.; Li, L.; Asghar, A.; Gorski, J.C. Diabetes mellitus increases the in vivo activity of cytochrome P450 2E1 in humans. Br. J. Clin. Pharmacol 2003, 55, 77–85. [Google Scholar]
- Khush, G. Productivity improvements in rice. Nutr. Rev 2003, 61, S114–S116. [Google Scholar]
- Hu, E.A.; Pan, A.; Malik, V.; Sun, Q. White rice consumption and risk of type 2 diabetes: Meta-analysis and systematic review. Br. Med. J 2012, 344. [Google Scholar] [CrossRef]
- Miller, J.B.; Pang, E.; Bramall, L. Rice: A high or low glycemic index food? Am. J. Clin. Nutr 1992, 56, 1034–1036. [Google Scholar]
- Panlasigui, L.N.; Thompson, L.U. Blood glucose lowering effects of brown rice in normal and diabetic subjects. Int. J. Food Sci. Nutr 2006, 57, 151–158. [Google Scholar]
- Sun, Q.; Donna, S.; Rob, M.V.D.; Michelle, D.H.; Vasanti, S.M.; Walter, C.W.; Frank, B.H. White rice, brown rice, and risk of type 2 diabetes in US men and women. Arch. Intern. Med 2010, 170, 961–969. [Google Scholar]
- Zhang, G.; Malik, V.S.; Pan, A.; Kumar, S.; Holmes, M.D.; Spiegelman, D.; Lin, X.; Hu, F.B. Substituting brown rice for white rice to lower diabetes risk: A focus-group study in Chinese adults. J. Am. Diet. Assoc 2010, 110, 1216–1221. [Google Scholar]
- Jenkins, D.J.A.; Kendall, C.W.C.; Augustin, L.S.A.; Franceschi, S.; Hamidi, M.; Marchie, A.; Jenkins, A.L.; Axelsen, M. Glycemic index: Overview of implications in health and disease. Am. J. Clin. Nutr 2002, 76, 266S–273S. [Google Scholar]
- Miller, A.; Engel, K.H. Content of γ-oryzanol and composition of steryl ferulates in brown rice (Oryza sativa L.) of European origin. J. Agric. Food Chem 2006, 54, 8127–8133. [Google Scholar]
- Lu, T.J.; Chen, H.N.; Wang, H.J. Chemical constituents, dietary fiber, and γ-oryzanol in six commercial varieties of brown rice from Taiwan. Cereal Chem 2011, 88, 463–466. [Google Scholar]
- Jayadeep, A.; Malleshi, N.G. Nutrients, composition of tocotrienols, tocopherols, and γ-oryzanol, and antioxidant activity in brown rice before and after biotransformation. CyTA J. Food 2011, 9, 82–87. [Google Scholar]
- Roohinejad, S.; Mirhosseini, H.; Saari, N.; Mustafa, S.; Alias, I.; Shobirin, M.H.A.; Hamid, A.; Manap, M.Y. Evaluation of GABA, crude protein and amino acid composition from different varieties of Malaysian’s brown rice. Aust. J. Crop Sci 2009, 3, 184–190. [Google Scholar]
- Hu, F.B.; van Dam, R.M.; Liu, S. Diet and risk of Type II diabetes: The role of types of fat and carbohydrate. Diabetologia 2001, 44, 805–817. [Google Scholar]
- Ludwig, D.S.; Majzoub, J.A.; Al-Zahrani, A.; Dallal, G.E.; Blanco, I.; Roberts, S.B. High glycemic index foods, overeating, and obesity. Pediatrics 1999, 103. [Google Scholar] [CrossRef]
- Brand-Miller, J.; Hayne, S.; Petocz, P.; Colagiuri, S. Low–glycemic index diets in the management of diabetes: A meta-analysis of randomized controlled trials. Diabetes Care 2003, 26, 2261–2267. [Google Scholar]
- Bailey, C.J.; Turner, R.C. Metformin. New Eng. J. Med 1996, 334, 574–579. [Google Scholar]
- Edenberg, H.J. The genetics of alcohol metabolism: Role of alcohol dehydrogenase and aldehyde. Alcohol. Res. Health 2007, 30, 5–13. [Google Scholar]
- Ingelman-Sundberg, M. Genetic polymorphisms of cytochrome P450 2D6 (CYP2D6): Clinical consequences, evolutionary aspects and functional diversity. Pharmacogenomics J 2005, 5, 6–13. [Google Scholar]
- Pelkonen, O.; Turpeinen, M.; Hakkola, J.; Honkakoski, P.; Hukkanen, J.; Raunio, H. Inhibition and induction of human cytochrome P450 enzymes: Current status. Arch. Toxicol 2008, 82, 667–715. [Google Scholar]
- Mitschke, D.; Reichel, A.; Fricker, G.; Moenning, U. Characterization of cytochrome P450 protein expression along the entire length of the intestine of male and female rats. Drug Metab. Dispos 2008, 36, 1039–1045. [Google Scholar]
- Schmider, J.; Greenblatt, D.J.; von Moltke, L.L.; Karsov, D.; Shader, R.I. Inhibition of CYP2C9 by selective serotonin reuptake inhibitors in vitro: Studies of phenytoin p-hydroxylation. Br. J. Clin. Pharmacol 1997, 44, 495–498. [Google Scholar]
- Caraco, Y.; Muszkat, M.; Wood, A.J. Phenytoin metabolic ratio: A putative marker of CYP2C9 activity in vivo. Pharmacogenetics 2001, 11, 587–596. [Google Scholar]
- Lee, C.R.; Pieper, J.A.; Frye, R.F.; Hinderliter, A.L.; Blaisdell, J.A.; Goldstein, J.A. Tolbutamide, flurbiprofen, and losartan as probes of CYP2C9 activity in humans. J. Clin. Pharmacol 2003, 43, 84–91. [Google Scholar]
- Owens, I.S.; Ritter, J.K.; Yeatman, M.T.; Chen, F. The novelUGT1 gene complex links bilirubin, xenobiotics, and therapeutic drug metabolism by encoding UDP-glucuronosyltransferase isozymes with a common carboxyl terminus. J. Pharmacokinet Biopharm 1996, 24, 491–508. [Google Scholar]
- Levin, B.E.; Hogan, S.; Sullivan, A.C. Initiation and perpetuation of obesity and obesity resistance in rats. Am. J. Physiol 1989, 256, R766–R771. [Google Scholar]
- Srinivasan, K.; Viswanad, B.; Asrat, L.; Kaul, C.L.; Ramarao, P. Combination of high-fat diet-fed and low-dose streptozotocin-treated rat: A model for type 2 diabetes and pharmacological screening. Pharmacol. Res 2005, 52, 313–320. [Google Scholar]



{kind=link}
{kind=link}
| Rat groups | Body weight (g) #,* | Blood glucose (mmol/L)* | Dietary composition | Food consumption (kcal/100 g body weight/day)* | ||||
|---|---|---|---|---|---|---|---|---|
| Calories (Kcal/100 g pellet) | Total carbohydrate (%) | Protein (%) | Fat (%) | Vitamins and minerals mix (%) | ||||
| Normal non-diabetic | 278 ± 16 a | 4.6 ± 0.5 a | 335 | 59.4 | 20.0 | 4.8 | 5.1 | 30.5 ± 3.7 a |
| Diabetic untreated | 337 ± 23 b | 14.9 ± 2.2 b | 548 | 47.7 | 16.1 | 31.1 | 5.1 | 34.0 ± 6.0 a |
| Metformin | 356 ± 28 b | 14.7 ± 4.1 b | 548 | 47.7 | 16.1 | 31.1 | 5.1 | 30.7 ± 6.0 a |
| White rice | 329 ± 16 b | 19.1 ± 2 b | 554 | 47.7 | 16.1 | 31.1 | 5.1 | 33.2 ± 8.3 a |
| Brown rice | 364 ± 18 b | 18.4 ± 2.8 b | 554 | 47.7 | 16.1 | 31.1 | 5.1 | 30.5 ± 6.7 a |
| Gene (Accession number) | *Fold change in relation to rat groups | ||
|---|---|---|---|
| Normal non-diabetic | Diabetic untreated | Metformin | |
| Cyp2D4 | WR 3.6 | WR 1.6 | WR 5.2 |
| BR 1.4 | BR 0.6 | BR 2 | |
| Cyp3A1 | WR 6 | WR 3.5 | WR 5.5 |
| BR 3 | BR 1.7 | BR 2.6 | |
| Cyp4A1 | WR 15 | WR 1 | WR 4.5 |
| BR 10 | BR 0.7 | BR 3 | |
| Cyp2B1 | WR 20 | WR 6.5 | WR 13.7 |
| BR 4.7 | BR 1.6 | BR 3.3 | |
| Cyp2E1 | WR 10 | WR 5 | WR 5.7 |
| BR 3.7 | BR 1.7 | BR 2 | |
| Cyp2C11 | WR 1.8 | WR 1.2 | WR 2.7 |
| BR 1.2 | BR 0.8 | BR 1.8 | |
| UGT2B1 | WR 4.2 | WR 4.2 | WR 4.2 |
| BR 2.4 | BR 2.4 | BR 2.4 | |
| ALDH1A1 | WR 15 | WR 0.8 | WR 3.8 |
| BR 5 | BR 0.3 | BR 1.3 | |
| Cyp2C6 | WR 17 | WR 1 | WR 5 |
| BR 3.7 | BR 0.2 | BR 1 | |
| Accession number | Genes |
|---|---|
| NM_017101 | peptidylprolyl isomerase A (Ppia)* |
| NM_175837 | Cyp4a1 |
| J00719 | Cyp2b1 |
| L24207 | Cyp3A1 |
| NM_031543 | Cyp2e1 |
| J02657 | Cyp2c11 |
| NM_138515 | Cyp2d4 |
| NM_173295 | Ugt2B1 |
| AF001898 | Aldh1A1 |
| M13711 | Cyp2C6 |
| NM_031144 | actin, beta (Actb)* |
| NM_017008 | glyceraldehyde-3-phosphate dehydrogenase (Gapd)* |
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Imam, M.U.; Ismail, M. Effects of Brown Rice and White Rice on Expression of Xenobiotic Metabolism Genes in Type 2 Diabetic Rats. Int. J. Mol. Sci. 2012, 13, 8597-8608. https://doi.org/10.3390/ijms13078597
Imam MU, Ismail M. Effects of Brown Rice and White Rice on Expression of Xenobiotic Metabolism Genes in Type 2 Diabetic Rats. International Journal of Molecular Sciences. 2012; 13(7):8597-8608. https://doi.org/10.3390/ijms13078597
Chicago/Turabian StyleImam, Mustapha Umar, and Maznah Ismail. 2012. "Effects of Brown Rice and White Rice on Expression of Xenobiotic Metabolism Genes in Type 2 Diabetic Rats" International Journal of Molecular Sciences 13, no. 7: 8597-8608. https://doi.org/10.3390/ijms13078597