The COX-2 Selective Blocker Etodolac Inhibits TNFα-Induced Apoptosis in Isolated Rabbit Articular Chondrocytes

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
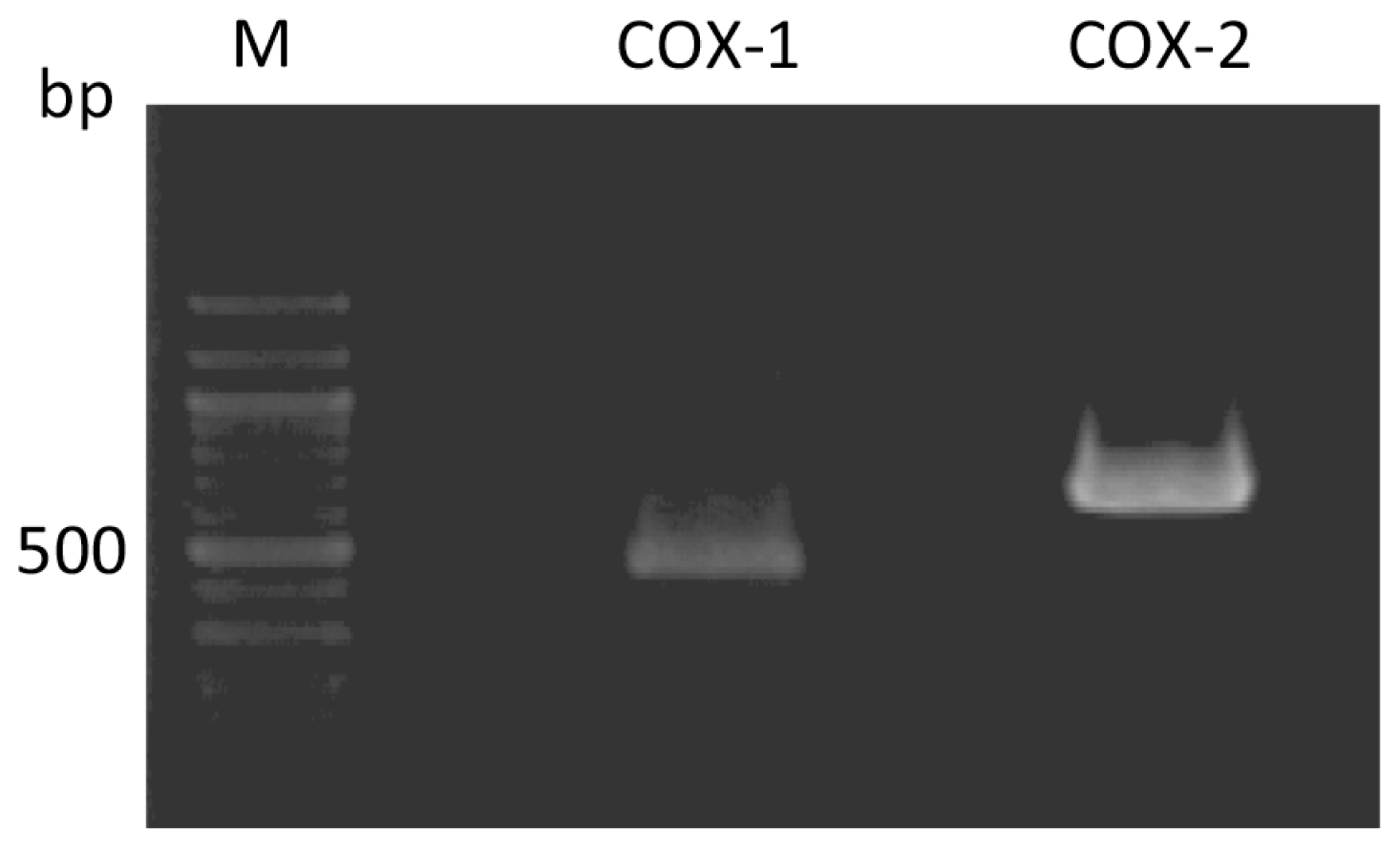
2.1. Expression of COX-1 and COX-2 in Rabbit Articular Chondrocytes
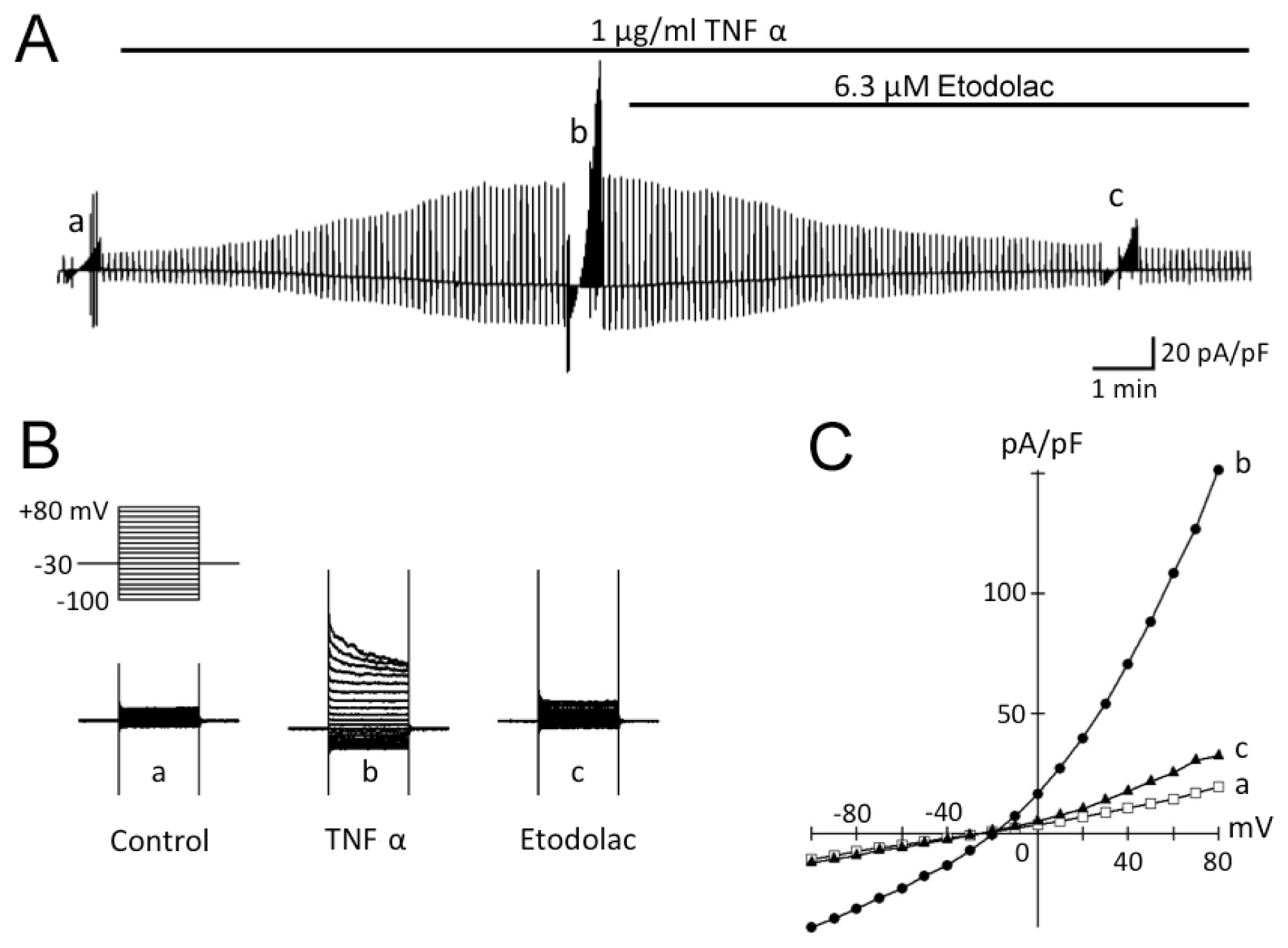
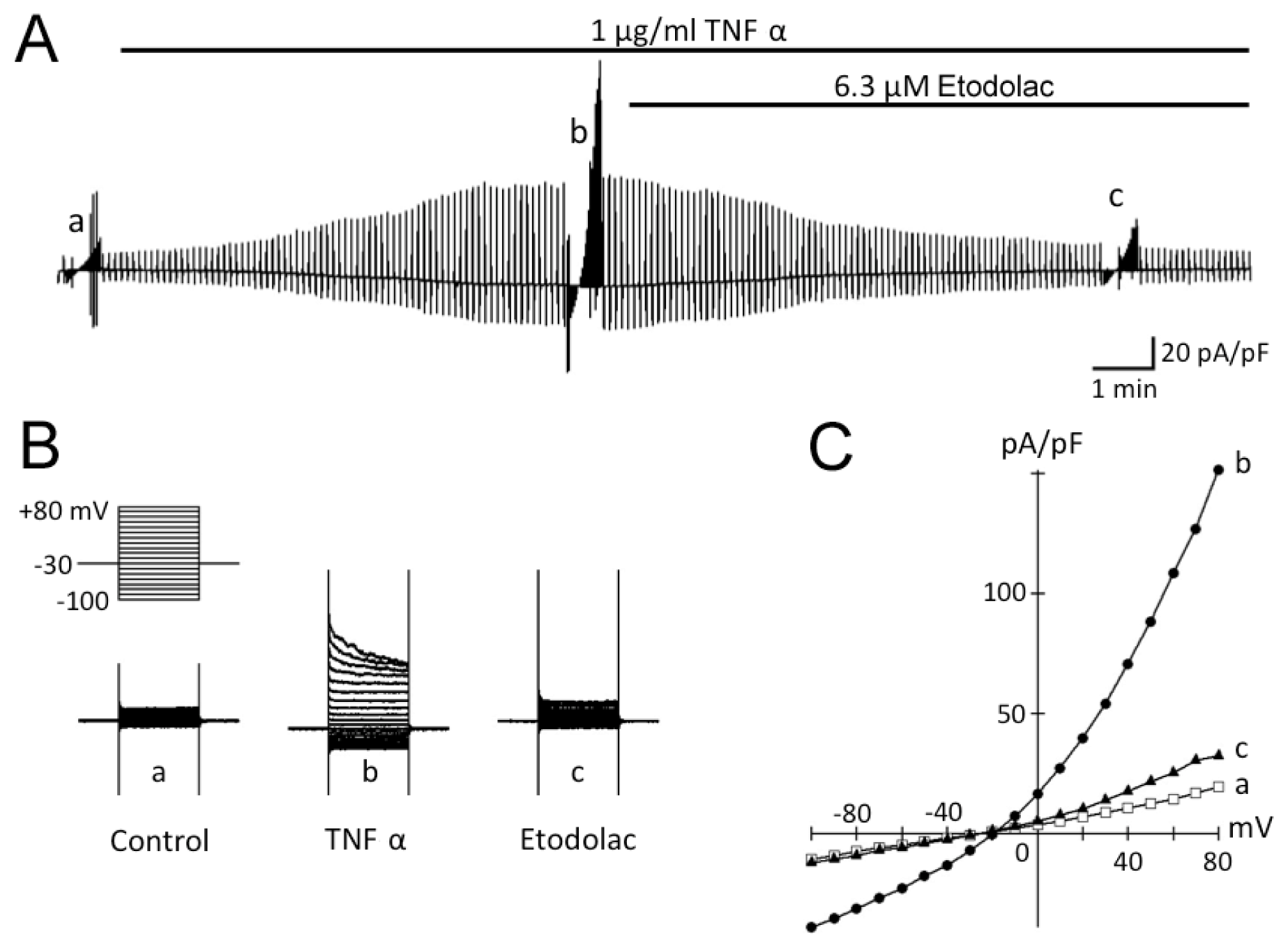
2.2. TNFα-Induced Activation of Cl− Current in Rabbit Articular Chondrocytes
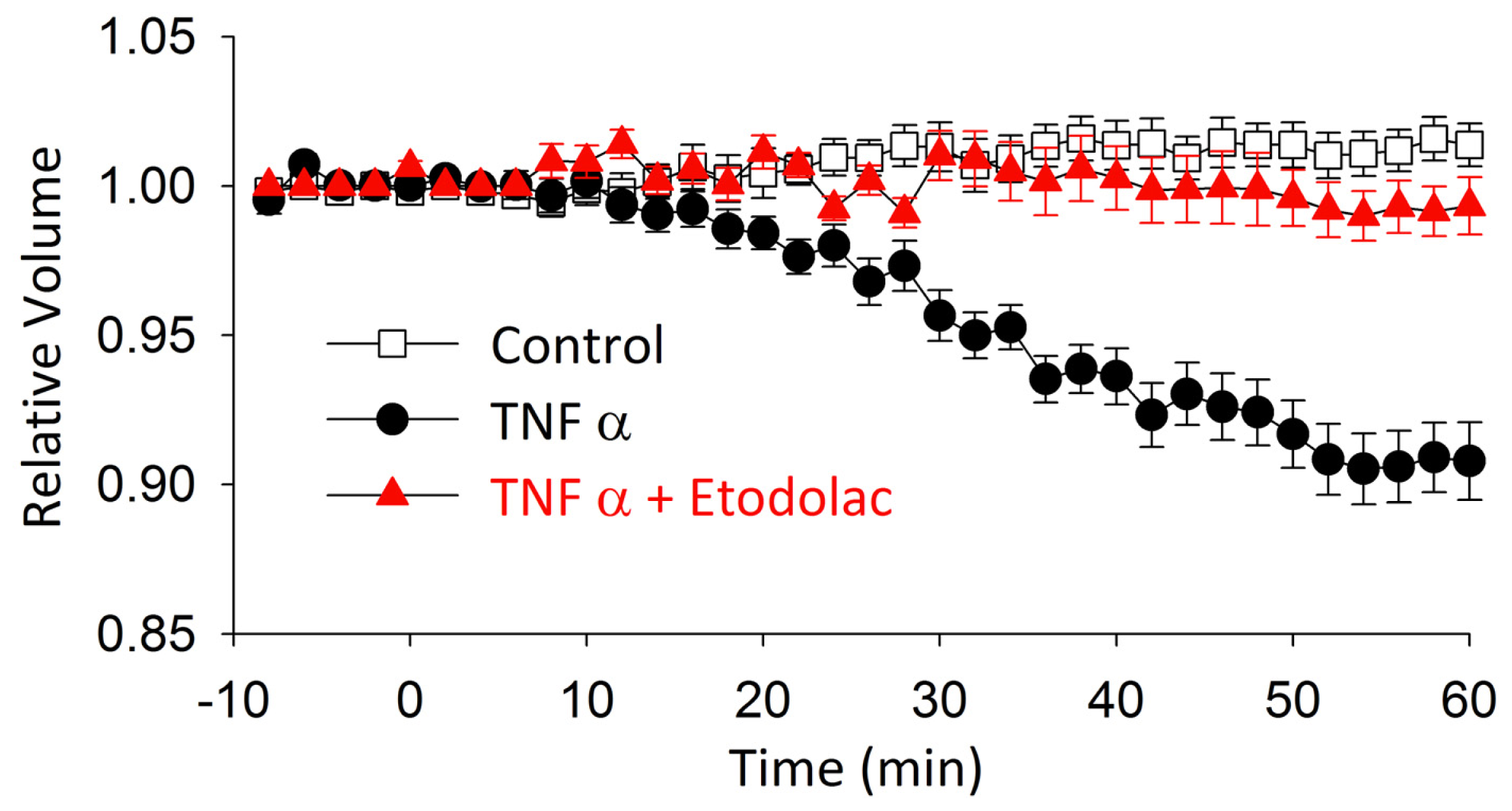
2.3. TNFα-Induced AVD and Its Inhibition by Etodolac
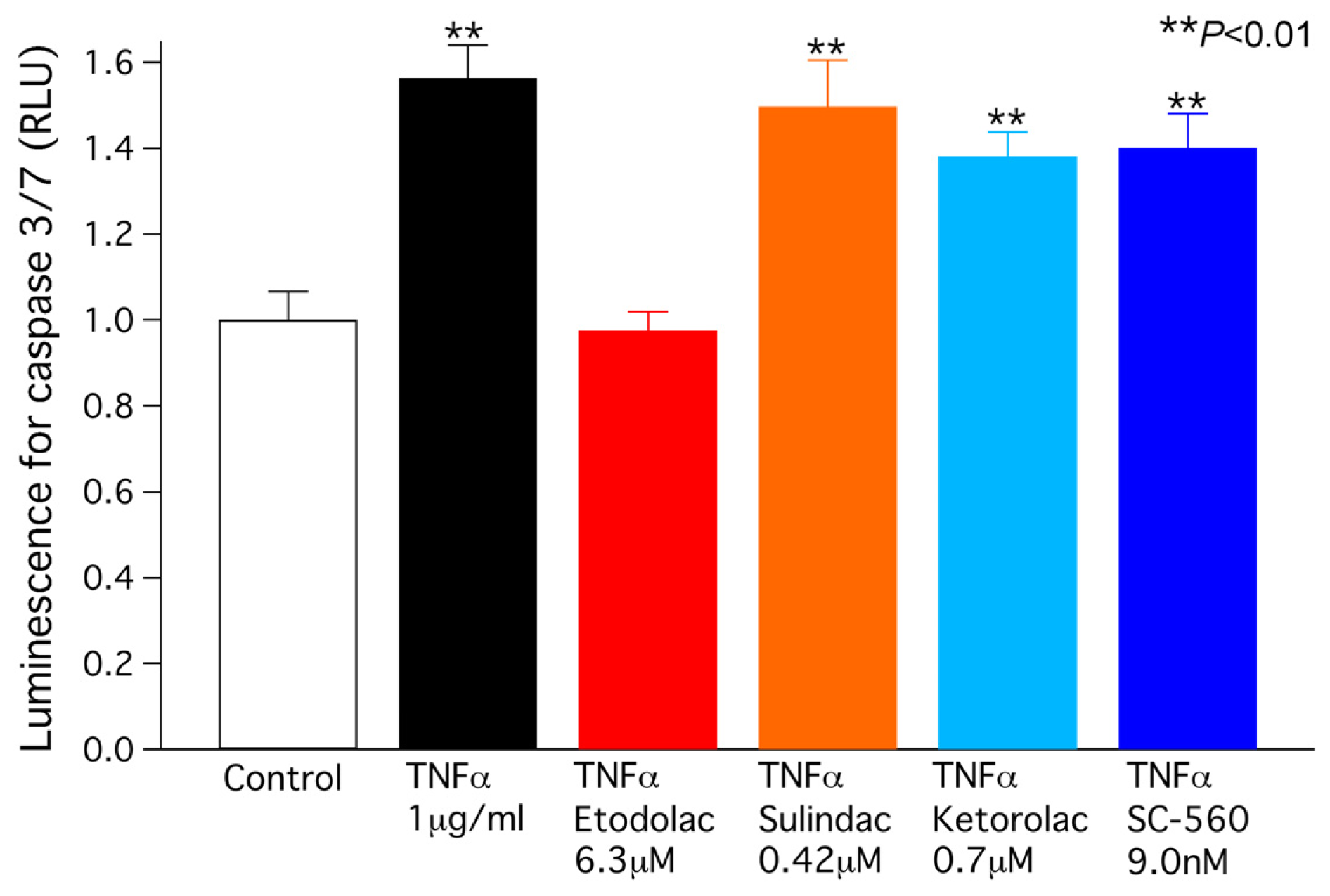
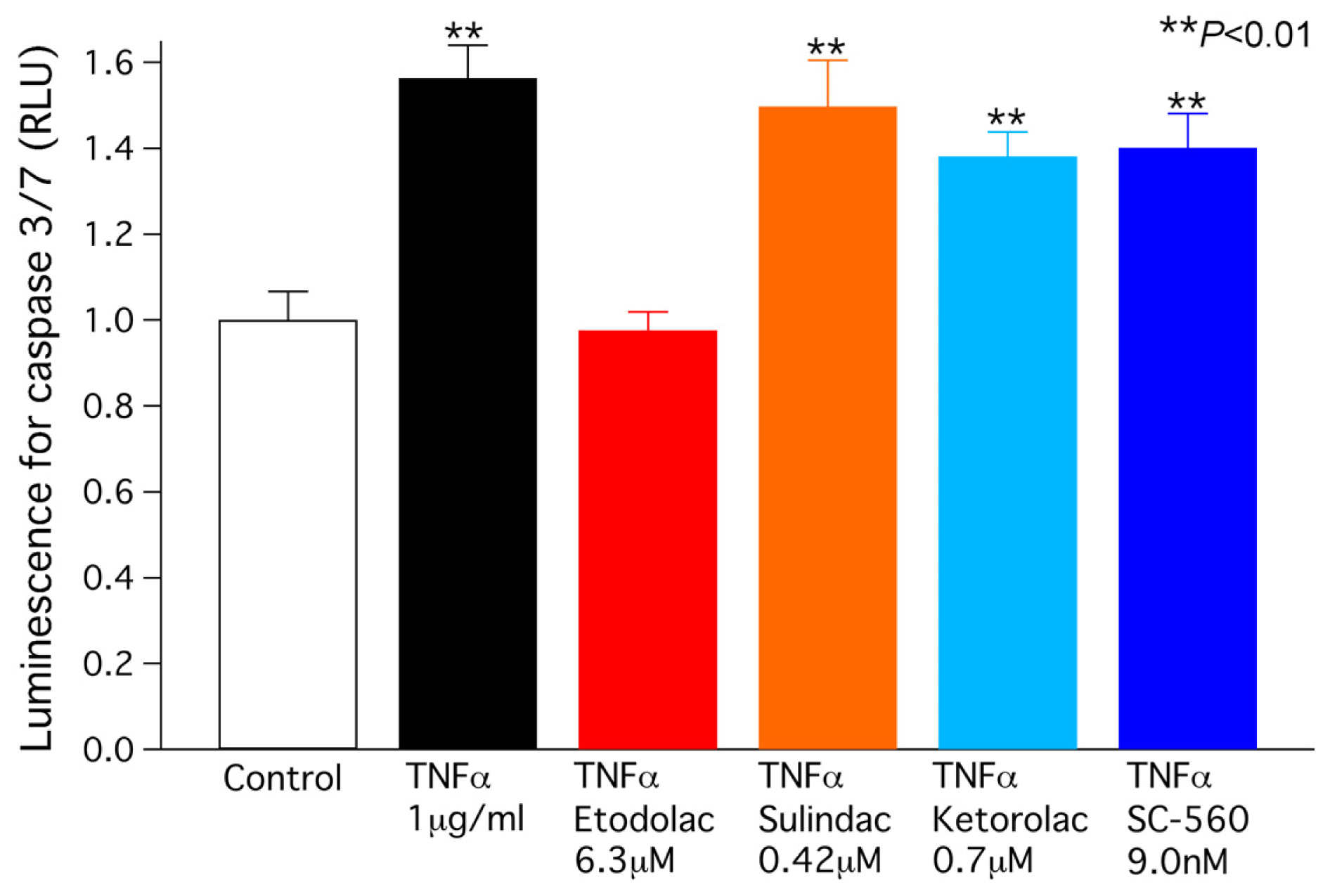
2.4. TNFα-Induced Caspase 3/7 Activity
3. Experimental Section
3.1. Chondrocyte Isolation
3.2. Solutions and Chemicals
3.3. Whole-Cell Patch-Clamp Technique and Data Analysis
3.4. Microscopy and Image Analysis
3.5. Caspase-3/7 Activity Measurement
3.6. Statistical Analysis
4. Conclusions

Acknowledgments
Conflicts of Interest
References
- Adams, C.S.; Shapiro, I.M. The fate of the terminally differentiated chondrocyte: Evidence for microenvironmental regulation of chondrocyte apoptosis. Crit. Rev. Oral Biol. Med 2002, 13, 465–473. [Google Scholar]
- Mobasheri, A. Role of chondrocyte death and hypocellularity in ageing human articular cartilage and the pathogenesis of osteoarthritis. Med. Hypotheses 2002, 58, 193–197. [Google Scholar]
- Blanco, F.J.; Guitian, R.; Vazquez-Martul, E.; de Toro, F.J.; Galdo, F. Osteoarthritis chondrocytes die by apoptosis. A possible pathway for osteoarthritis pathology. Arthr. Rheum 1998, 41, 284–289. [Google Scholar]
- Hashimoto, S.; Ochs, R.L.; Komiya, S.; Lotz, M. Linkage of chondrocyte apoptosis and cartilage degradation in human osteoarthritis. Arthr. Rheum 1998, 41, 1632–1638. [Google Scholar]
- Maroudas, A.; Venn, M. Chemical composition and swelling of normal and osteoarthrotic femoral head cartilage. II. Swelling. Ann. Rheum. Dis 1977, 36, 399–406. [Google Scholar]
- Lewis, R.; Feetham, C.H.; Barrett-Jolley, R. Cell volume regulation in chondrocytes. Cell. Physiol. Biochem 2011, 28, 1111–1122. [Google Scholar]
- Isoya, E.; Toyoda, F.; Imai, S.; Okumura, N.; Kumagai, K.; Omatsu-Kanbe, M.; Kubo, M.; Matsuura, H.; Matsusue, Y. Swelling-activated Cl− current in isolated rabbit articular chondrocytes: Inhibition by arachidonic acid. J. Pharmacol. Sci 2009, 109, 293–304. [Google Scholar]
- Okumura, N.; Imai, S.; Toyoda, F.; Isoya, E.; Kumagai, K.; Matsuura, H.; Matsusue, Y. Regulatory role of tyrosine phosphorylation in the swelling-activated chloride current in isolated rabbit articular chondrocytes. J. Physiol 2009, 587, 3761–3776. [Google Scholar]
- Kumagai, K.; Imai, S.; Toyoda, F.; Okumura, N.; Isoya, E.; Matsuura, H.; Matsusue, Y. 17beta-Oestradiol inhibits doxorubicin-induced apoptosis via block of the volume-sensitive Cl(−) current in rabbit articular chondrocytes. Br. J. Pharmacol 2012, 166, 702–720. [Google Scholar]
- Okada, Y.; Sato, K.; Numata, T. Pathophysiology and puzzles of the volume-sensitive outwardly rectifying anion channel. J. Physiol 2009, 587, 2141–2149. [Google Scholar]
- Feyen, J.H.; van der Wilt, G.; Moonen, P.; di Bon, A.; Nijweide, P.J. Stimulation of arachidonic acid metabolism in primary cultures of osteoblast-like cells by hormones and drugs. Prostaglandins 1984, 28, 769–781. [Google Scholar]
- Harrison, J.R.; Lorenzo, J.A.; Kawaguchi, H.; Raisz, L.G.; Pilbeam, C. Stimulation of prostaglandin E2 production by interleukin-1 alpha and transforming growth factor alpha in osteoblastic MC3T3-E1 cells. J. Bone Miner. Res 1994, 9, 817–823. [Google Scholar]
- Rodan, S.B.; Rodan, G.A.; Simmons, H.A.; Walenga, R.W.; Feinstein, M.B.; Raisz, L.G. Bone resorptive factor produced by osteosarcoma cells with osteoblastic features is PGE2. Biochem. Biophys. Res. Commun 1981, 102, 1358–1365. [Google Scholar]
- Fletcher, B.S.; Kujubu, D.A.; Perrin, D.M.; Herschman, H.R. Structure of the mitogen-inducible TIS10 gene and demonstration that the TIS10-encoded protein is a functional prostaglandin G/H synthase. J. Biol. Chem 1992, 267, 4338–4344. [Google Scholar]
- Kujubu, D.A.; Fletcher, B.S.; Varnum, B.C.; Lim, R.W.; Herschman, H.R. TIS10, a phorbol ester tumor promoter-inducible mRNA from Swiss 3T3 cells, encodes a novel prostaglandin synthase/cyclooxygenase homologue. J. Biol. Chem 1991, 266, 12866–12872. [Google Scholar]
- Xie, W.L.; Chipman, J.G.; Robertson, D.L.; Erikson, R.L.; Simmons, D.L. Expression of a mitogen-responsive gene encoding prostaglandin synthase is regulated by mRNA splicing. Proc. Natl. Acad. Sci. USA 1991, 88, 2692–2696. [Google Scholar]
- Khanna, I.K.; Weier, R.M.; Yu, Y.; Collins, P.W.; Miyashiro, J.M.; Koboldt, C.M.; Veenhuizen, A.W.; Currie, J.L.; Seibert, K.; Isakson, P.C. 1,2-Diarylpyrroles as potent and selective inhibitors of cyclooxygenase-2. J. Med. Chem 1997, 40, 1619–1633. [Google Scholar]
- Warner, T.D.; Giuliano, F.; Vojnovic, I.; Bukasa, A.; Mitchell, J.A.; Vane, J.R. Nonsteroid drug selectivities for cyclo-oxygenase-1 rather than cyclo-oxygenase-2 are associated with human gastrointestinal toxicity: A full in vitro analysis. Proc. Natl. Acad. Sci. USA 1999, 96, 7563–7568. [Google Scholar]
- Jones, R.A. Etodolac: An overview of a selective COX-2 inhibitor. Inflammopharmacology 1999, 7, 269–275. [Google Scholar]
- Yanaoka, K.; Oka, M.; Yoshimura, N.; Deguchi, H.; Mukoubayashi, C.; Enomoto, S.; Maekita, T.; Inoue, I.; Ueda, K.; Utsunomiya, H.; et al. Preventive effects of etodolac, a selective cyclooxygenase-2 inhibitor, on cancer development in extensive metaplastic gastritis, a Helicobacter pylori-negative precancerous lesion. Int. J. Cancer 2010, 126, 1467–1473. [Google Scholar]
- Steinbach, G.; Lynch, P.M.; Phillips, R.K.; Wallace, M.H.; Hawk, E.; Gordon, G.B.; Wakabayashi, N.; Saunders, B.; Shen, Y.; Fujimura, T.; et al. The effect of celecoxib, a cyclooxygenase-2 inhibitor, in familial adenomatous polyposis. N. Engl. J. Med 2000, 342, 1946–1952. [Google Scholar]
- Nishida, Y.; Tsukushi, S.; Shido, Y.; Wasa, J.; Ishiguro, N.; Yamada, Y. Successful treatment with meloxicam, a cyclooxygenase-2 inhibitor, of patients with extra-abdominal desmoid tumors: A pilot study. J. Clin. Oncol 2010, 28, e107–e109. [Google Scholar]
- Prins, J.B.; Niesler, C.U.; Winterford, C.M.; Bright, N.A.; Siddle, K.; O’Rahilly, S.; Walker, N.I.; Cameron, D.P. Tumor necrosis factor-alpha induces apoptosis of human adipose cells. Diabetes 1997, 46, 1939–1944. [Google Scholar]
- Maeno, E.; Ishizaki, Y.; Kanaseki, T.; Hazama, A.; Okada, Y. Normotonic cell shrinkage because of disordered volume regulation is an early prerequisite to apoptosis. Proc. Natl. Acad. Sci. USA 2000, 97, 9487–9492. [Google Scholar]
- L’Hoste, S.; Chargui, A.; Belfodil, R.; Corcelle, E.; Duranton, C.; Rubera, I.; Poujeol, C.; Mograbi, B.; Tauc, M.; Poujeol, P. CFTR mediates apoptotic volume decrease and cell death by controlling glutathione efflux and ROS production in cultured mice proximal tubules. Am. J. Physiol. Renal Physiol 2010, 298, F435–F453. [Google Scholar]
- Nicholson, D.W.; Thornberry, N.A. Caspases: Killer proteases. Trends Biochem. Sci 1997, 22, 299–306. [Google Scholar]
- Garcia-Calvo, M.; Peterson, E.P.; Rasper, D.M.; Vaillancourt, J.P.; Zamboni, R.; Nicholson, D.W.; Thornberry, N.A. Purification and catalytic properties of human caspase family members. Cell Death Differ 1999, 6, 362–369. [Google Scholar]
- John, T.; Muller, R.D.; Oberholzer, A.; Zreiqat, H.; Kohl, B.; Ertel, W.; Hostmann, A.; Tschoeke, S.K.; Schulze-Tanzil, G. Interleukin-10 modulates pro-apoptotic effects of TNF-alpha in human articular chondrocytes in vitro. Cytokine 2007, 40, 226–234. [Google Scholar]
- Lopez-Armada, M.J.; Carames, B.; Lires-Dean, M.; Cillero-Pastor, B.; Ruiz-Romero, C.; Galdo, F.; Blanco, F.J. Cytokines, tumor necrosis factor-alpha and interleukin-1beta, differentially regulate apoptosis in osteoarthritis cultured human chondrocytes. Osteoarthr. Cartil 2006, 14, 660–669. [Google Scholar]
- Kim, J.; Xu, M.; Xo, R.; Mates, A.; Wilson, G.L.; Pearsall, A.W.T.; Grishko, V. Mitochondrial DNA damage is involved in apoptosis caused by pro-inflammatory cytokines in human OA chondrocytes. Osteoarthr. Cartil 2010, 18, 424–432. [Google Scholar]
- National Research Council, Guide for the Care and Use of Laboratory Animals, 8th ed; National Academies Press: Washington, DC, USA, 2011.
- Wilson, J.R.; Duncan, N.A.; Giles, W.R.; Clark, R.B. A voltage-dependent K+ current contributes to membrane potential of acutely isolated canine articular chondrocytes. J. Physiol 2004, 557, 93–104. [Google Scholar]
- Urban, J.P. The chondrocyte: A cell under pressure. Br. J. Rheumatol 1994, 33, 901–908. [Google Scholar]
- Hall, A.C. Volume-sensitive taurine transport in bovine articular chondrocytes. J. Physiol 1995, 484, 755–766. [Google Scholar]
- Fabiato, A.; Fabiato, F. Calculator programs for computing the composition of the solutions containing multiple metals and ligands used for experiments in skinned muscle cells. J. Physiol 1979, 75, 463–505. [Google Scholar]
- Tsien, R.Y.; Rink, T.J. Neutral carrier ion-selective microelectrodes for measurement of intracellular free calcium. Biochim. Biophys. Acta 1980, 599, 623–638. [Google Scholar]
- Hamill, O.P.; Marty, A.; Neher, E.; Sakmann, B.; Sigworth, F.J. Improved patch-clamp techniques for high-resolution current recording from cells and cell-free membrane patches. Pflug. Arch 1981, 391, 85–100. [Google Scholar]
- Lewis, R.S.; Ross, P.E.; Cahalan, M.D. Chloride channels activated by osmotic stress in T lymphocytes. J. Gen. Physiol 1993, 101, 801–826. [Google Scholar]
- Sakaguchi, M.; Matsuura, H.; Ehara, T. Swelling-induced Cl− current in guinea-pig atrial myocytes: Inhibition by glibenclamide. J. Physiol 1997, 505, 41–52. [Google Scholar]
- Benitah, J.P.; Gomez, A.M.; Bailly, P.; da Ponte, J.P.; Berson, G.; Delgado, C.; Lorente, P. Heterogeneity of the early outward current in ventricular cells isolated from normal and hypertrophied rat hearts. J. Physiol 1993, 469, 111–138. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Kumagai, K.; Kubo, M.; Imai, S.; Toyoda, F.; Maeda, T.; Okumura, N.; Matsuura, H.; Matsusue, Y. The COX-2 Selective Blocker Etodolac Inhibits TNFα-Induced Apoptosis in Isolated Rabbit Articular Chondrocytes. Int. J. Mol. Sci. 2013, 14, 19705-19715. https://doi.org/10.3390/ijms141019705
Kumagai K, Kubo M, Imai S, Toyoda F, Maeda T, Okumura N, Matsuura H, Matsusue Y. The COX-2 Selective Blocker Etodolac Inhibits TNFα-Induced Apoptosis in Isolated Rabbit Articular Chondrocytes. International Journal of Molecular Sciences. 2013; 14(10):19705-19715. https://doi.org/10.3390/ijms141019705
Chicago/Turabian StyleKumagai, Kousuke, Mitsuhiko Kubo, Shinji Imai, Futoshi Toyoda, Tsutomu Maeda, Noriaki Okumura, Hiroshi Matsuura, and Yoshitaka Matsusue. 2013. "The COX-2 Selective Blocker Etodolac Inhibits TNFα-Induced Apoptosis in Isolated Rabbit Articular Chondrocytes" International Journal of Molecular Sciences 14, no. 10: 19705-19715. https://doi.org/10.3390/ijms141019705