Genome-Wide Gene Expression Profiles in Antioxidant Pathways and Their Potential Sex Differences and Connections to Vitamin C in Mice



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Expression Levels of Oxidative Genes between Female and Male in sfx Mice in Comparison to Its Wild-Type Control
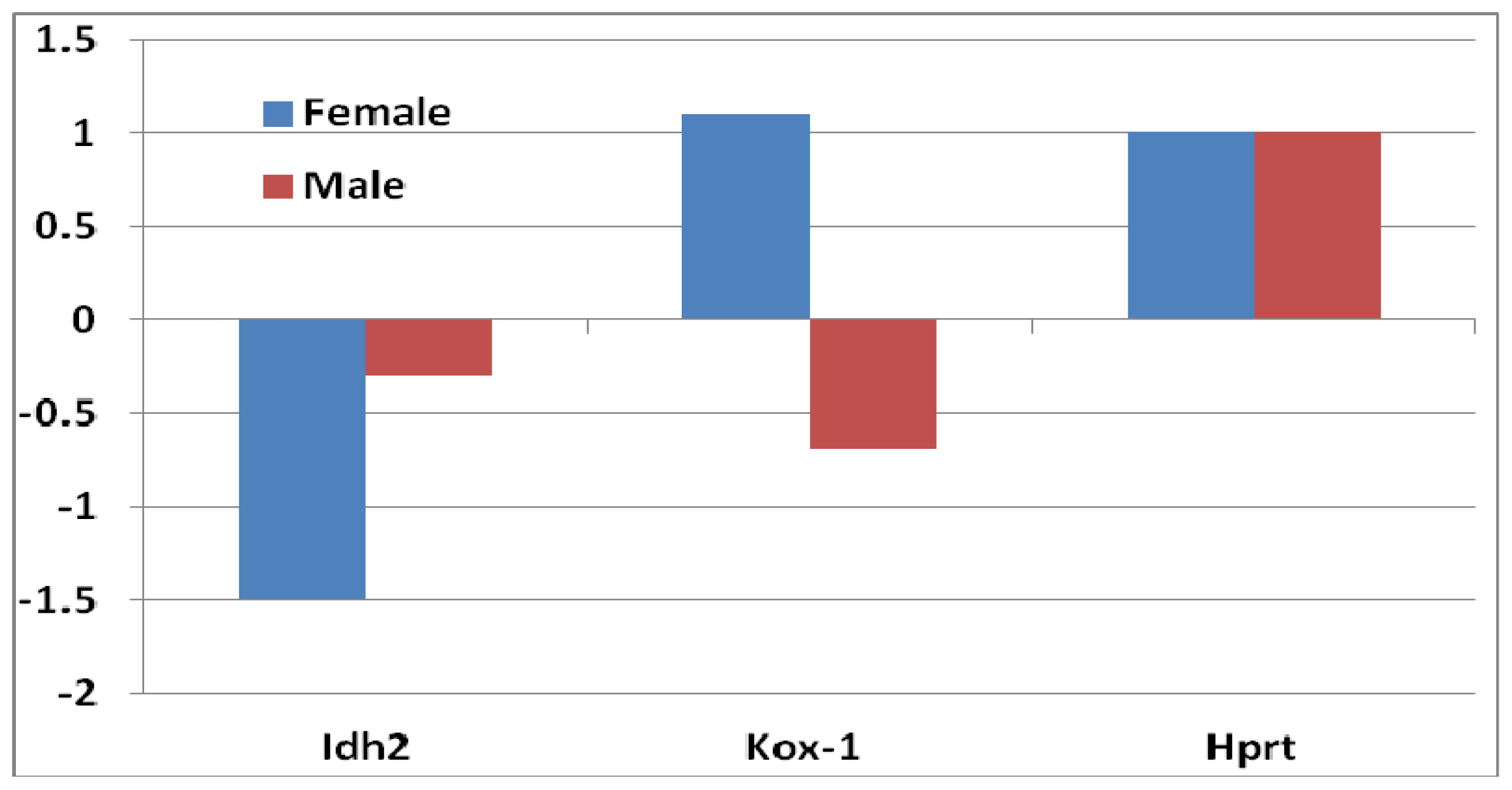
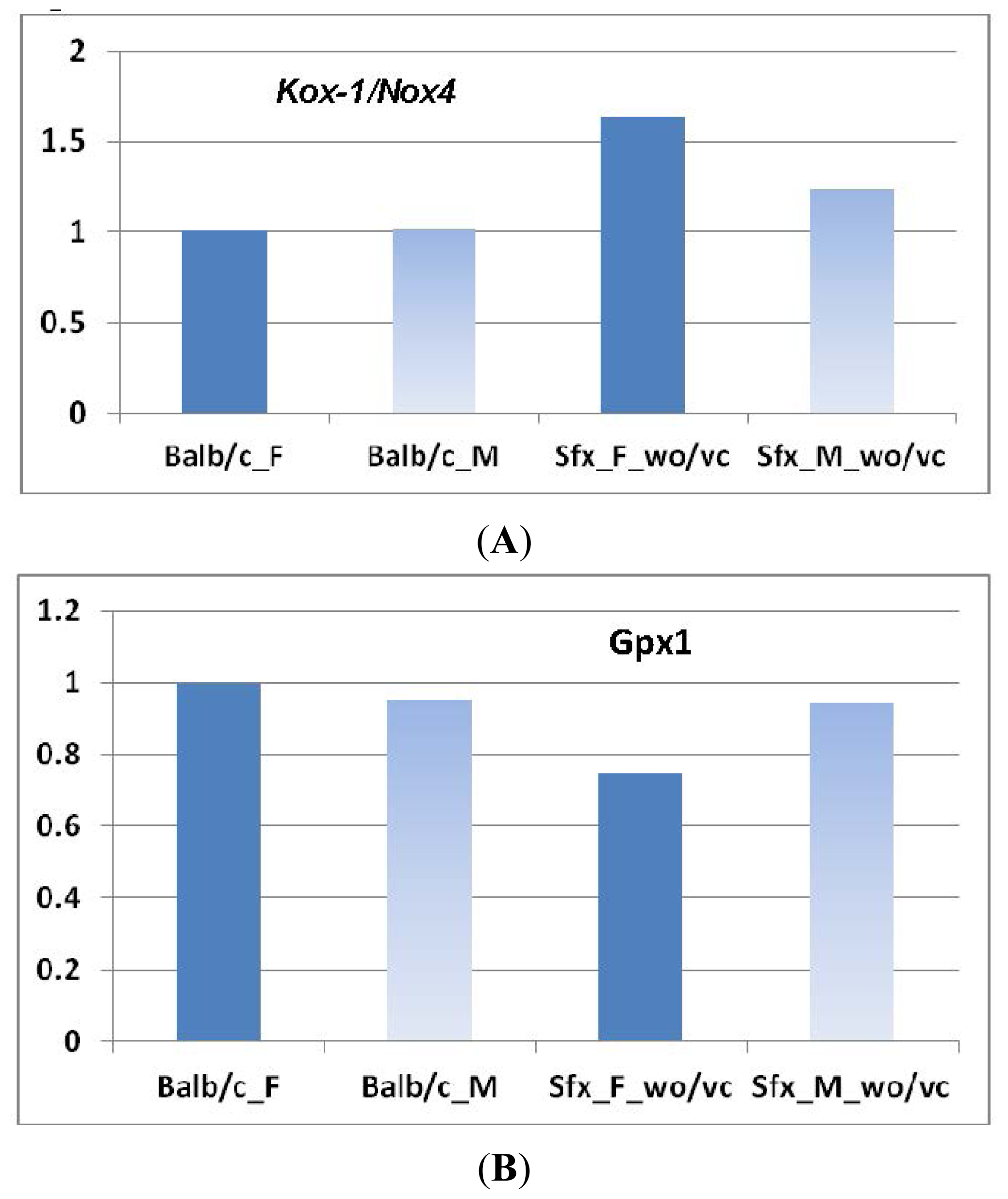
2.1.1. Significant Changes in Pro-Oxidative Enzymes
2.1.2. Significant Changes in Antioxidative Enzymes
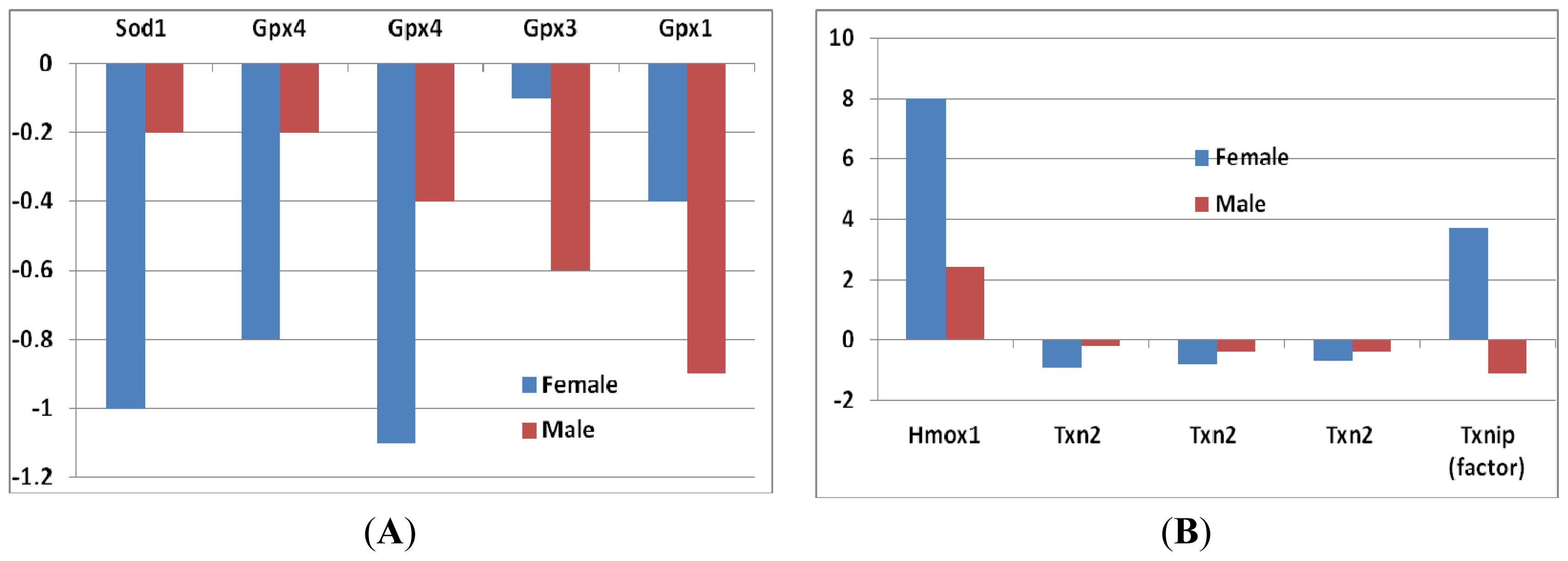
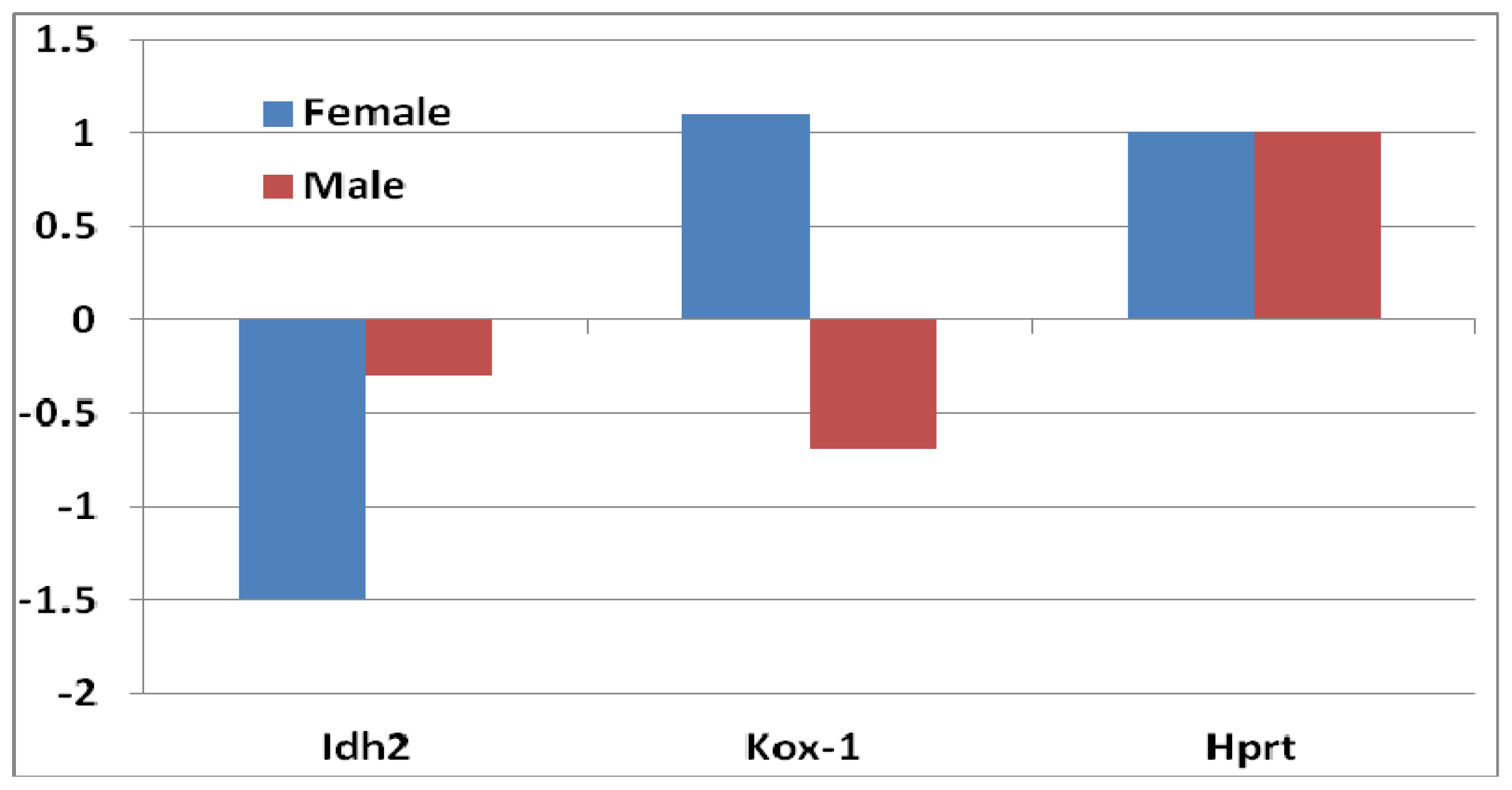
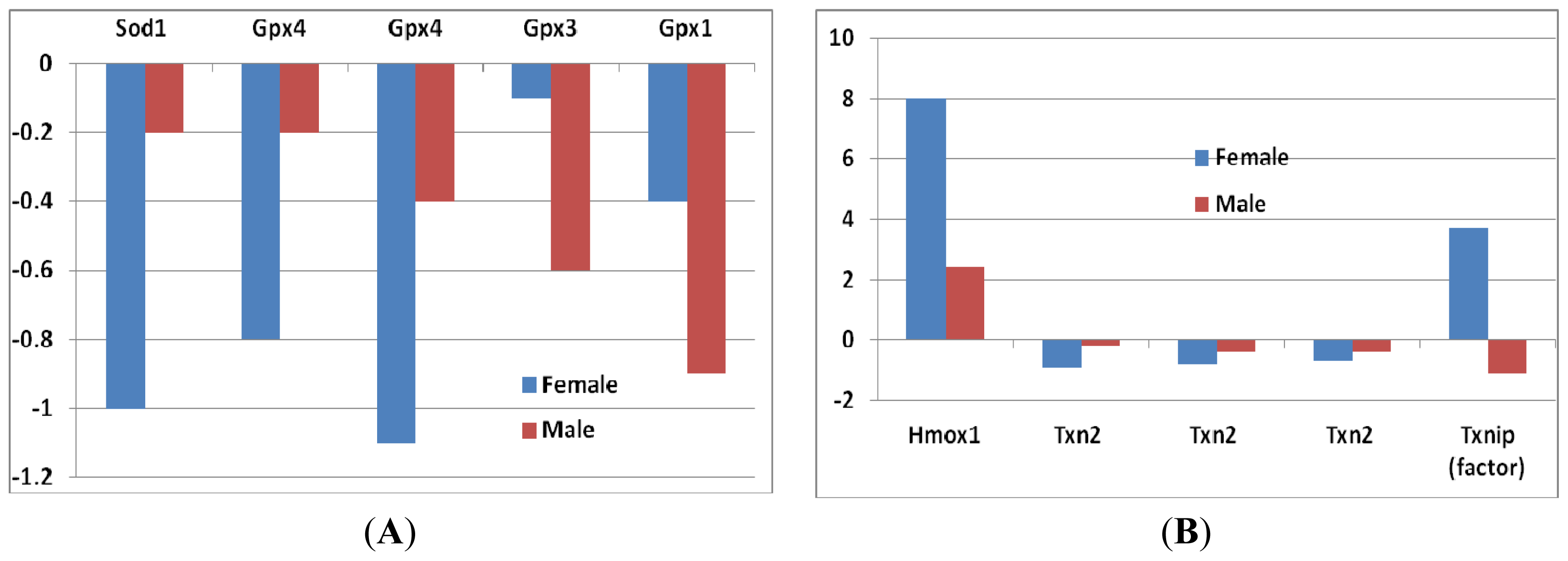
- Superoxide dismutase 1 (Sod1): In female mice, the decrease of Sod1 expression was greater compared to that of male mice (Figure 2A), although it did not reach to a significant level.
- Glutathione peroxidase (Gpx): Four probes of Gpxs suggested a decrease of Gpx genes in sfx mice. The decrease of Gpx4 in female mice was greater than that in male mice, while the decrease of Gpx1 and Gpx3 in female was smaller than that in male mice (Figure 2A). None of those differences reaches to the significance level.
- Heme oxygenase: The expression level of heme oxygenase (decycling) 1 (Hmox1) increased in both sexes, while the increase was much greater in females than that in males (Figure 2B). High levels of heme oxygenase-1 expression of cells can provide an antioxidant effect on skin, as well as anti-inflammatory properties, in mammals and rodents.
- Thioredoxin reductase (TrxR) and thioredoxin (Trx): These enzymes are major regulators of intracellular protein thiol redox balance [18]. Their prolonged inhibition can disrupt a number of redox-sensitive functions in cells. Thioredoxin 2 (Txn2) was decreased in both sexes, but the decrease in females was greater than that in males. The expression level of thioredoxin interacting factor (Txnip) increased in females, while it decreased in males (Figure 2B).
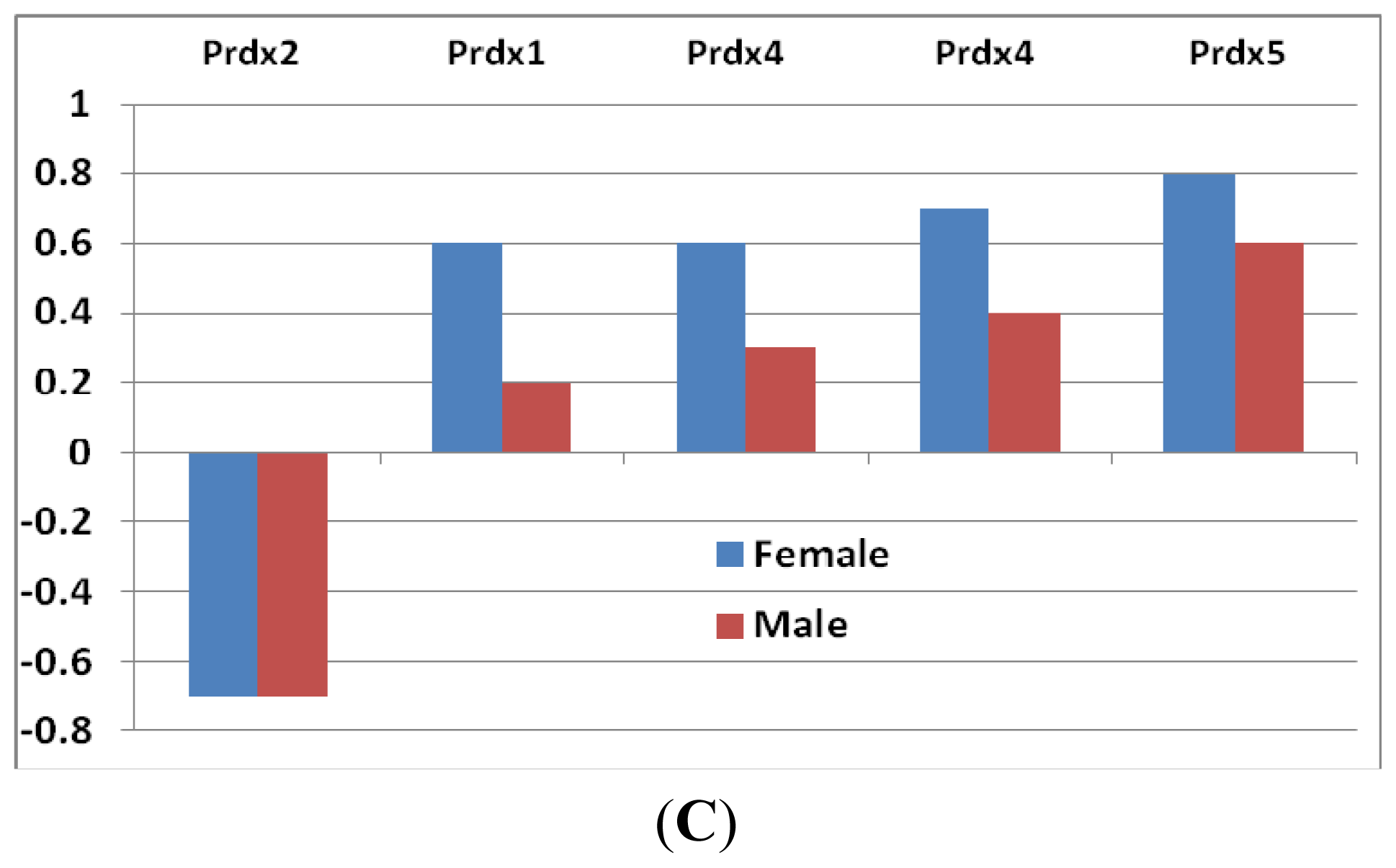
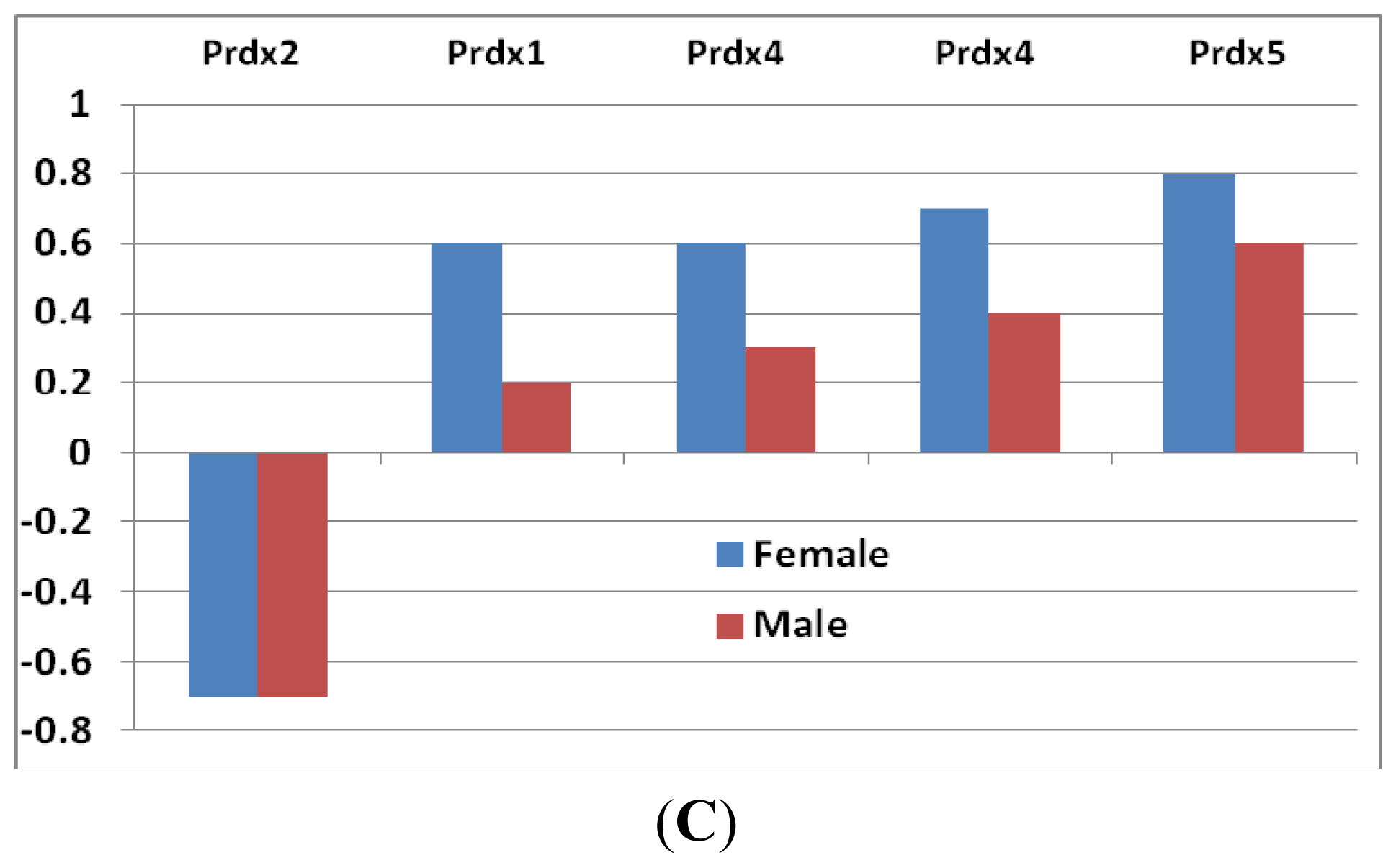
- Peroxiredoxins: These are important hydroperoxide detoxification enzymes, yet have only come to the fore in recent years relative to other major players in peroxide detoxification, heme-containing catalases and peroxidases and glutathione peroxidases [19]. Five family members of peroxiredoxin (Prdx) genes showed changes in expression levels (Figure 2C). Prx2 showed a decrease in both sexes, while the other three (Prdx1, Prdx4 and Prdx5) increased. The level of decrease in Prdx2 was similar in both sexes. The increases of the other three Prdxs, however, were greater in females than that in males (Figure 2C).
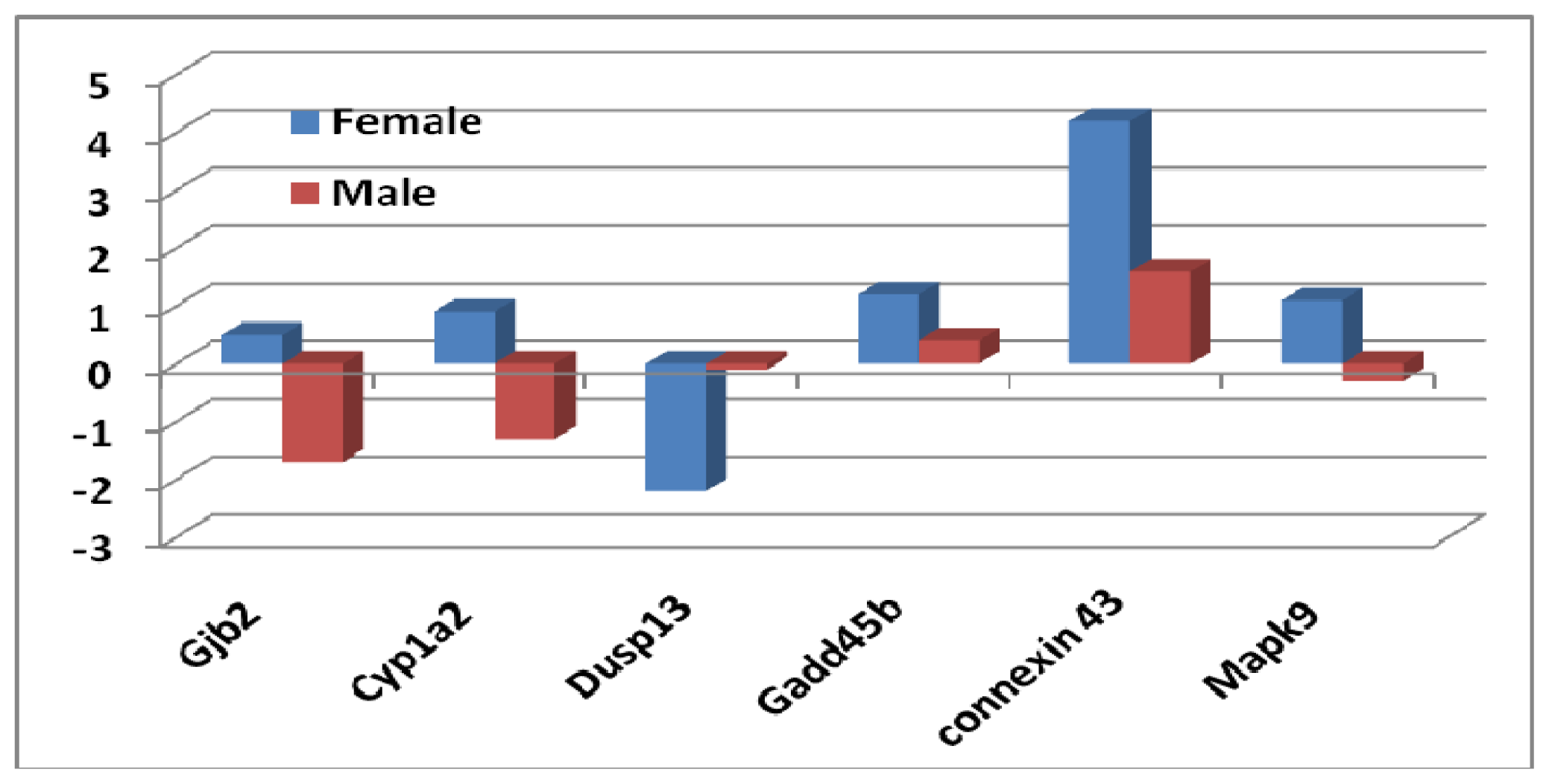
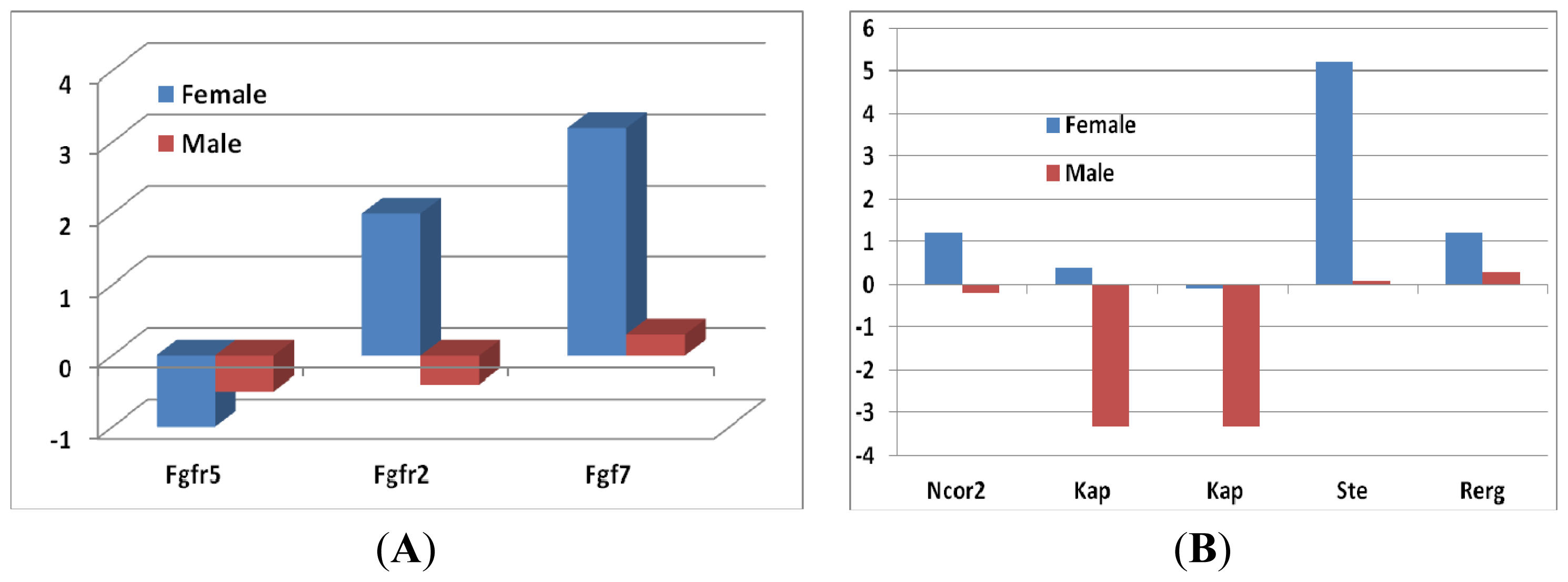
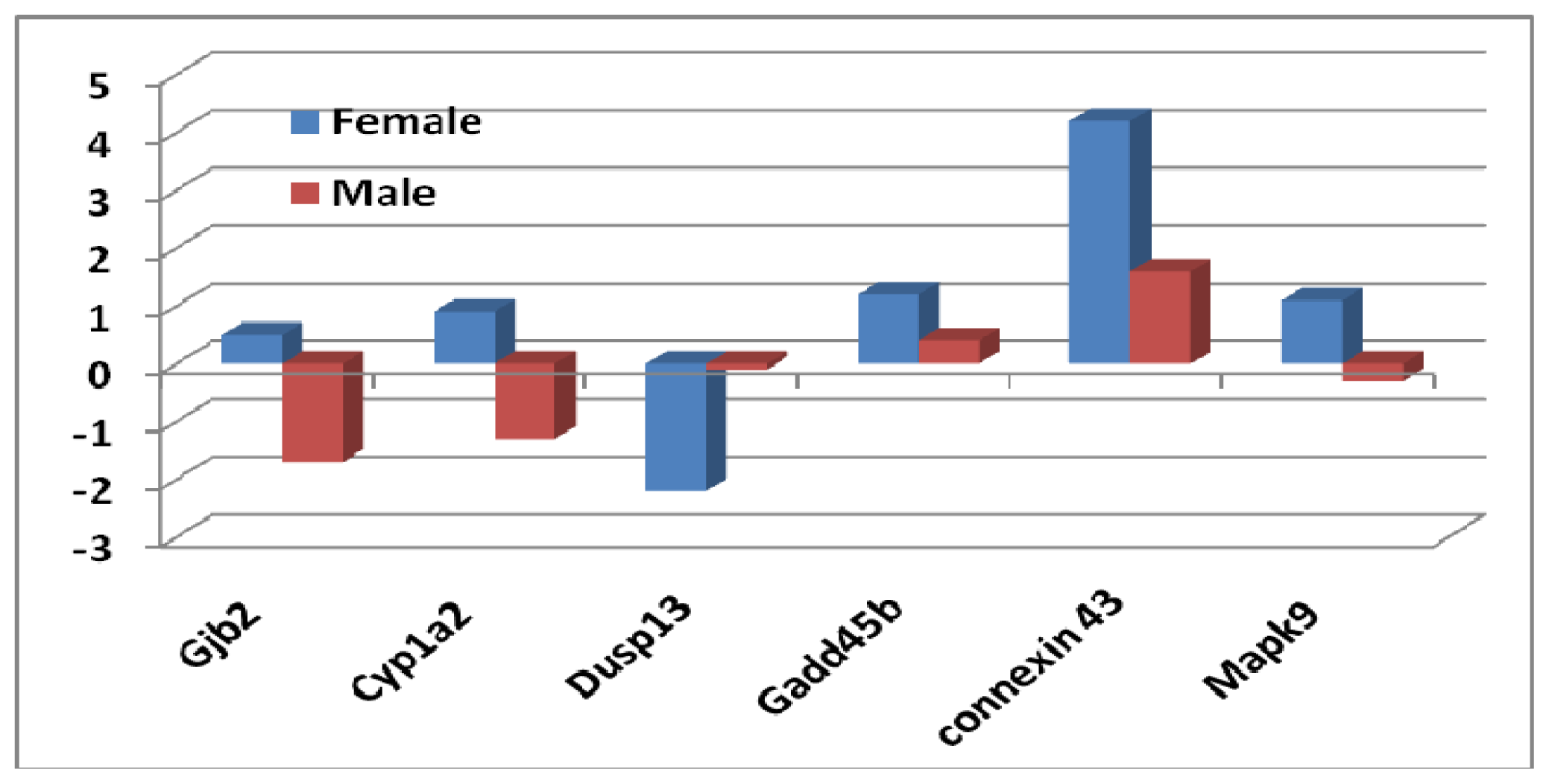
2.1.3. Sex Differential Expression of Genes Involved in Regulating Mapk Signaling
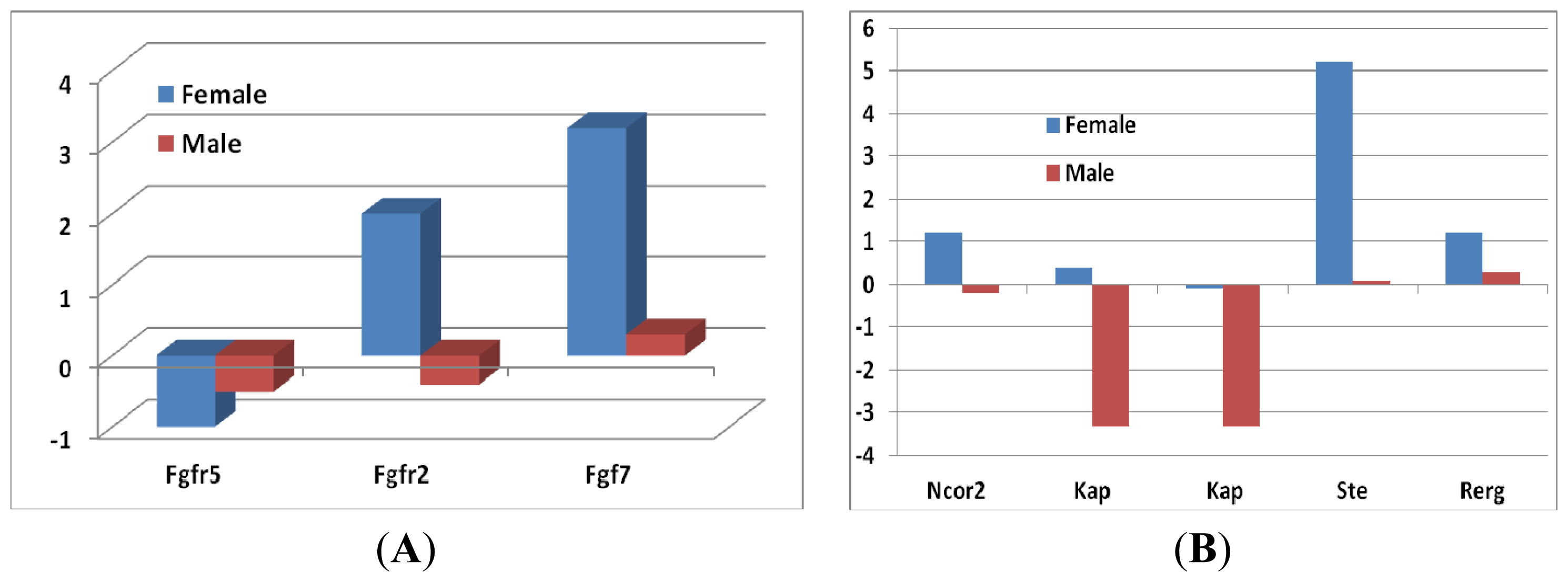
2.1.4. Gene Expression Levels of Growth Hormones in Female and in Male Mice between Sfx and WT Mice
2.1.5. Validation of Microarray Data Using Real-Time qPCR
2.2. Pathway Analysis Using Gene Expression Profiles of BXD Mice
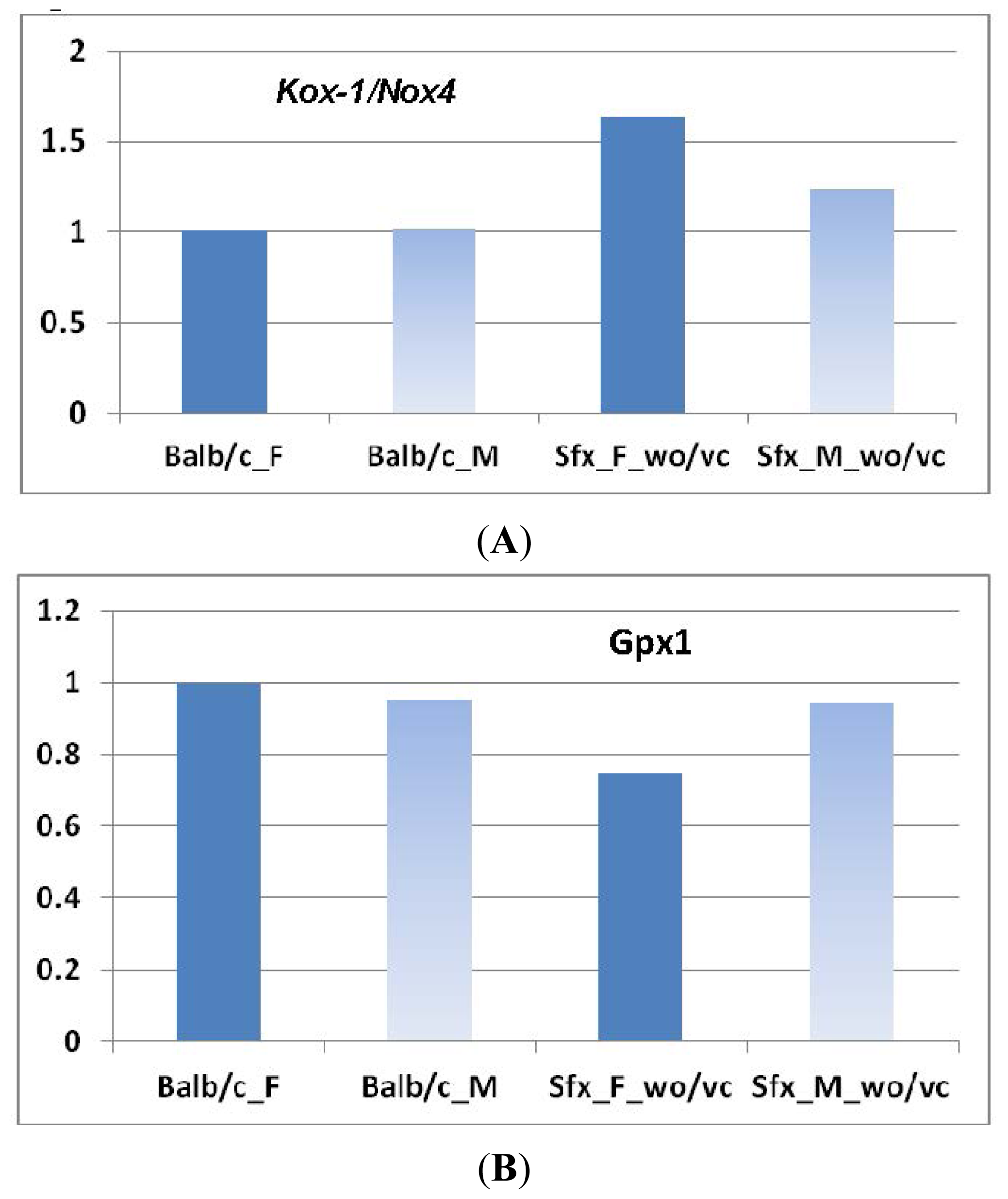
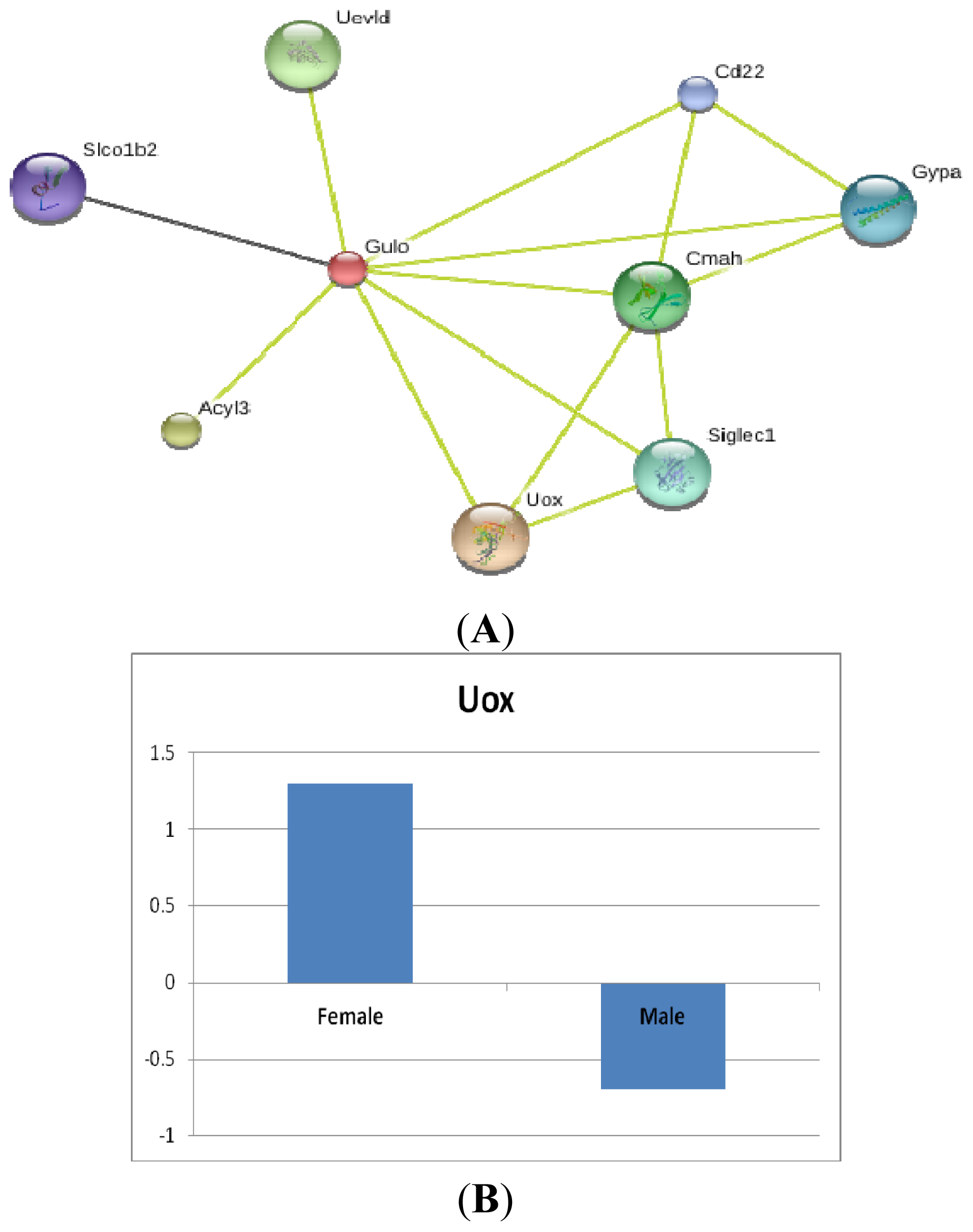
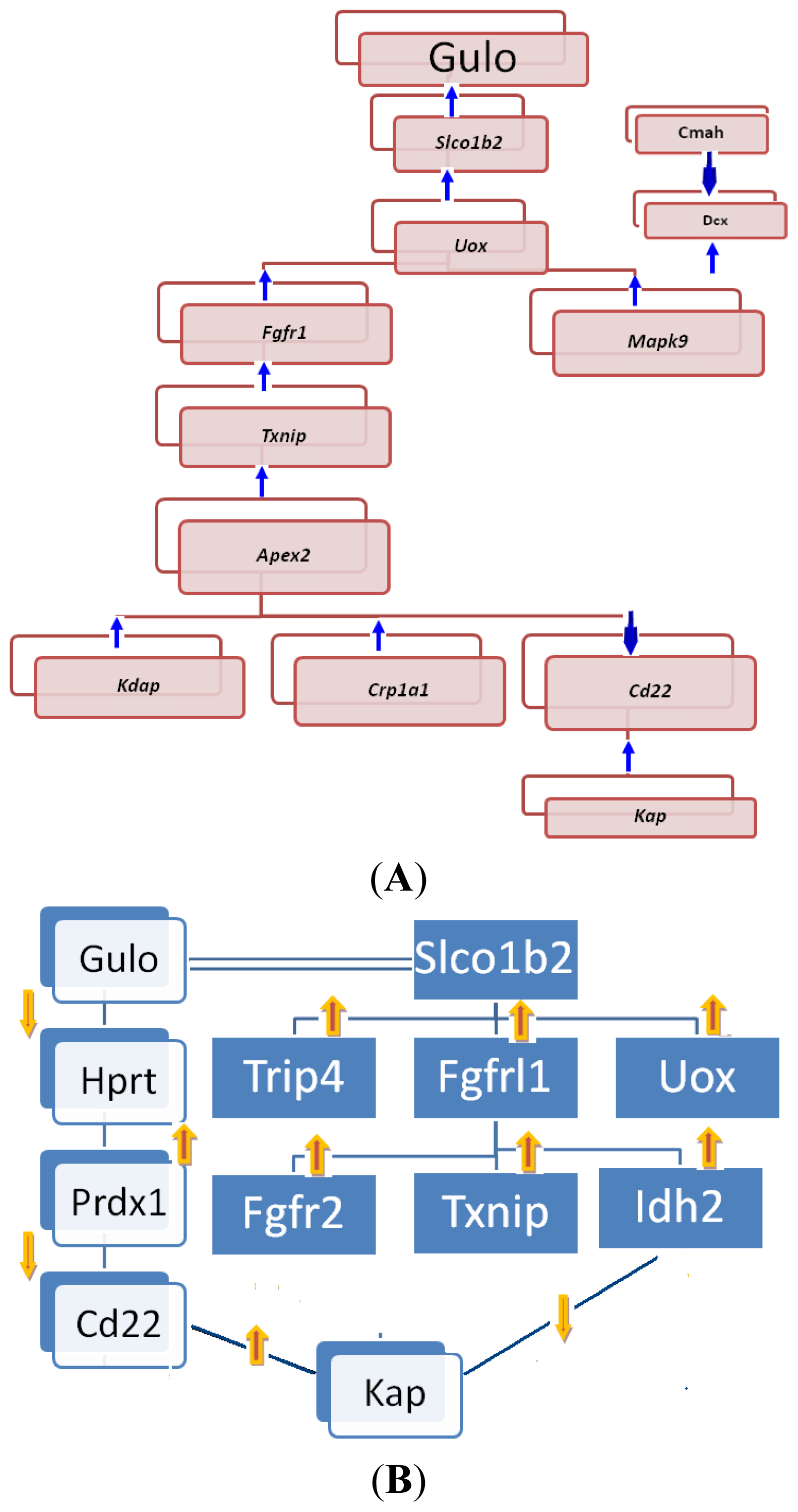
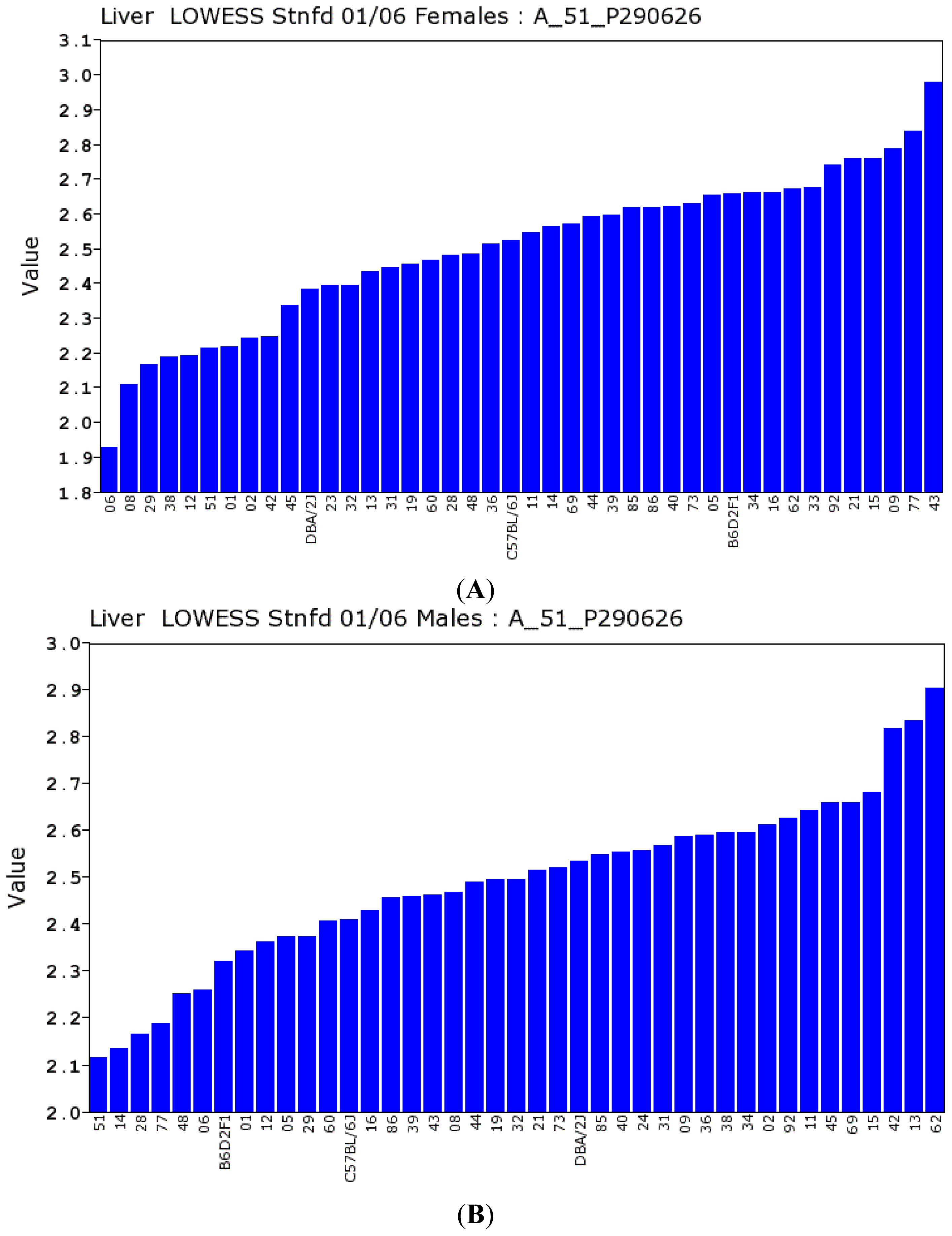
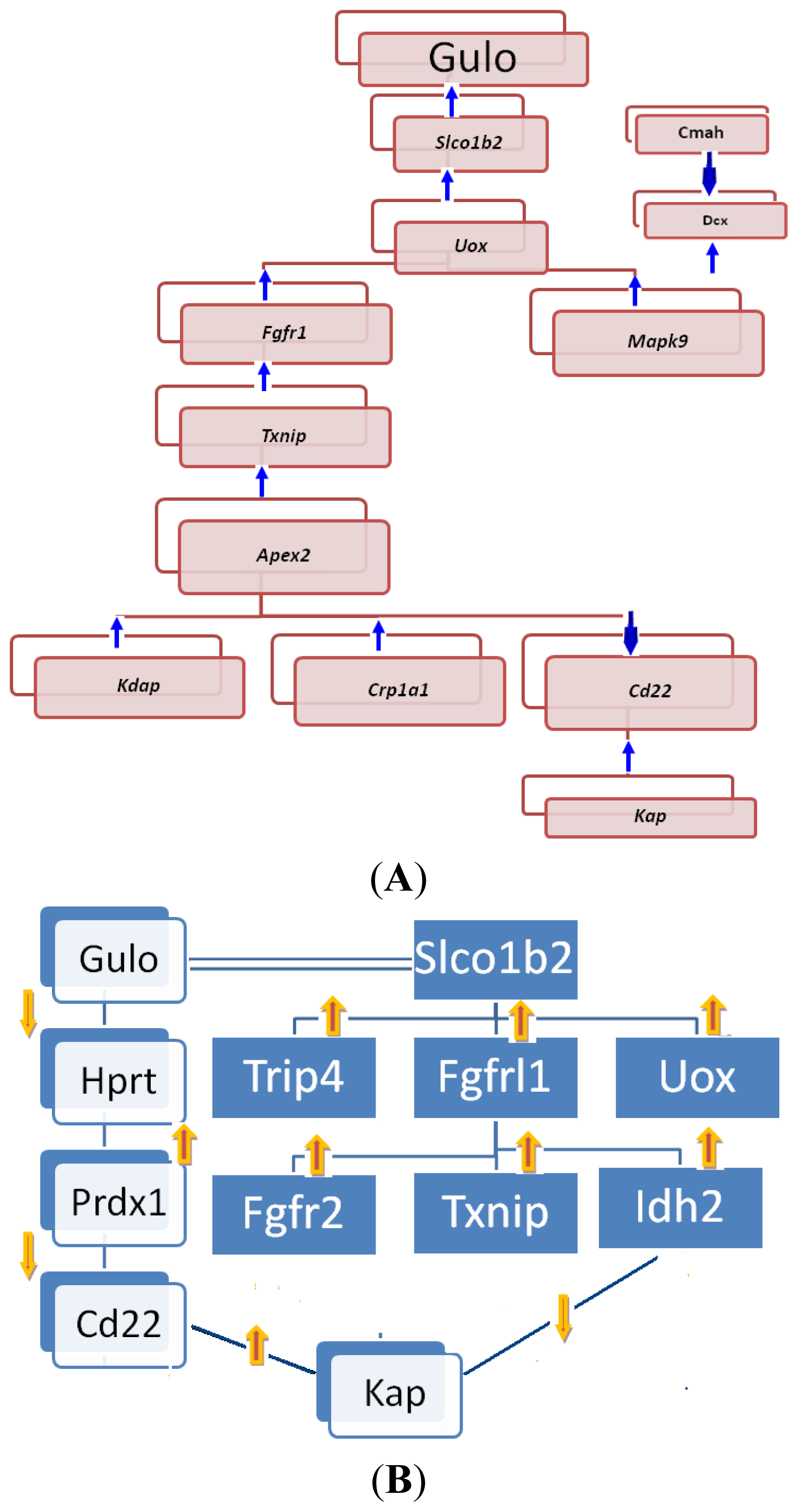
2.2.1. Gulo Gene and Its Partners
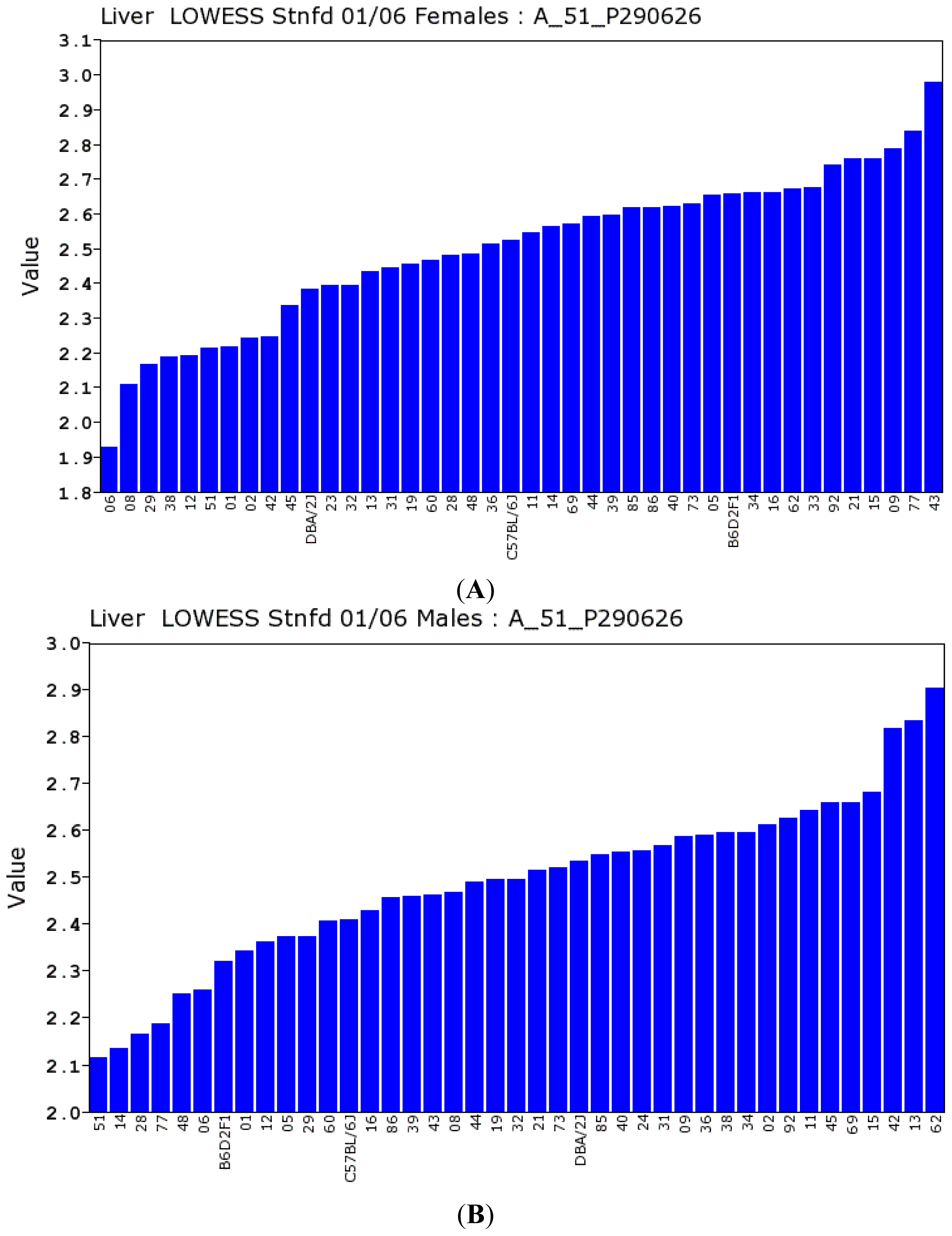
2.2.2. Transcriptomic Loci that Regulate Gulo in Female and Male Mice
2.2.3. Potential Gene Network Eluted from Whole-Genome Expression Profiles of Livers of BXD Strains
3. Materials and Methods
3.1. Animals
3.2. Procedure of Analysis of sfx Mice
3.3. Whole-Genome Expression Data of RI Strains of BXD Mice
3.4. Transcriptome Mapping
3.5. Association of Expression Levels among Genes
4. Conclusions
Acknowledgments
Conflict of Interest
References
- Levine, M.; Wang, Y.; Padayatty, S.J.; Morrow, J. A new recommended dietary allowance of vitamin C for healthy young women. Proc. Natl. Acad. Sci. USA 2001, 98, 9842–9846. [Google Scholar]
- Fain, O.; Pariés, J.; Jacquart, B.; Le Moël, G.; Kettaneh, A.; Stirnemann, J.; Héron, C.; Sitbon, M.; Taleb, C.; Letellier, E.; et al. Hypovitaminosis C in hospitalized patients. Eur. J. Intern. Med 2003, 14, 419–425. [Google Scholar]
- Maruyama, C.; Araki, R.; Takeuchi, M.; Kuniyoshi, E.; Iwasawa, A.; Maruyama, T.; Nakano, S.; Motohashi, Y.; Nakanishi, M.; Kyotani, S.; et al. Relationships of nutrient intake and lifestyle-related factors to serum folate and plasma homocysteine concentrations in 30–69 year-old Japanese. J. Nutr. Sci. Vitaminol. (Tokyo) 2004, 50, 1–8. [Google Scholar]
- Beitz, R.; Mensink, G.B.; Fischer, B. Blood pressure and vitamin C and fruit and vegetable intake. Ann. Nutr. Metab 2003, 47, 214–220. [Google Scholar]
- Al-Rejaie, S.S.; Abuohashish, H.M.; Alkhamees, O.A.; Aleisa, A.M.; Alroujayee, A.S. Gender difference following high cholesterol diet induced renal injury and the protective role of rutin and ascorbic acid combination in Wistar albino rats. Lipids Health Dis 2012, 11, 41. [Google Scholar]
- Alpsoy, L.; Yalvac, M.E. Key roles of vitamins A, C, and E in aflatoxin B1-induced oxidative stress. Vitam. Horm 2011, 86, 287–305. [Google Scholar]
- Pohanka, M.; Pejchal, J.; Snopkova, S.; Havlickova, K.; Karasova, J.Z.; Bostik, P.; Pikula, J. Ascorbic acid: An old player with a broad impact on body physiology including oxidative stress suppression and immunomodulation: A review. Mini Rev. Med. Chem 2012, 12, 35–43. [Google Scholar]
- Förstermann, U. Nitric oxide and oxidative stress in vascular disease. Pflugers. Arch 2010, 459, 923–939. [Google Scholar]
- Minghetti, L.; Greco, A.; Zanardo, V.; Suppiej, A. Early-life sex-dependent vulnerability to oxidative stress: The natural twining model. J. Matern. Fetal Neonatal Med 2012, 26, 259–262. [Google Scholar]
- Giergiel, M.; Lopucki, M.; Stachowicz, N.; Kankofer, M. The influence of age and gender on antioxidant enzyme activities in humans and laboratory animals. Aging Clin. Exp. Res 2012, 24, 256–259. [Google Scholar]
- Beamer, W.G.; Rosen, C.J.; Bronson, R.T.; Gu, W.; Donahue, L.R.; Baylink, D.J.; Richardson, C.C.; Crawford, G.C.; Barker, J.E. Spontaneous fracture (sfx): A mouse genetic model of defective peripubertal bone formation. Bone 2000, 27, 619–626. [Google Scholar]
- Jiao, Y.; Li, X.; Beamer, W.G.; Yan, J.; Tong, Y.; Goldowitz, D.; Roe, B.; Gu, W. A deletion causing spontaneous fracture identified from a candidate region of mouse Chromosome 14. Mamm. Genome 2005, 16, 20–31. [Google Scholar]
- Yan, J.; Jiao, Y.; Li, X.; Jiao, F.; Beamer, W.G.; Rosen, C.J.; Gu, W. Evaluation of gene expression profiling in a mouse model of l-Gulonolactone oxidase gene deficiency. Genet. Mol. Biol 2007, 30, 322–329. [Google Scholar]
- Jiao, Y.; Zhang, J.; Yan, J.; Stuart, J.; Gibson, G.; Lu, L.; Williams, R.; Wang, Y.J.; Gu, W. Differential gene expression between wild-type and Gulo-deficient mice supplied with vitamin C. Genet. Mol. Biol 2011, 34, 386–395. [Google Scholar]
- Hayes, K.R.; Vollrath, A.L.; Zastrow, G.M.; McMillan, B.J.; Craven, M.; Jovanovich, S.; Rank, D.R.; Penn, S.; Walisser, J.A.; Reddy, J.K.; et al. EDGE: A centralized resource for the comparison, analysis, and distribution of toxicogenomic information. Mol. Pharmacol 2005, 67, 1360–1368. [Google Scholar]
- Quinn, M.T.; Gauss, K.A. Structure and regulation of the neutrophil respiratory burst oxidase: Comparison with nonphagocyte oxidases. J. Leukocyte Biol 2004, 76, 760–781. [Google Scholar]
- Groemping, Y.; Rittinger, K. Activation and assembly of the NADPH oxidase: A structural perspective. Biochem. J 2005, 386, 401–416. [Google Scholar]
- Myers, C.R.; Myers, J.M.; Kufahl, T.D.; Forbes, R.; Szadkowski, A. The effects of acrolein on the thioredoxin system: Implications for redox-sensitive signaling. Mol. Nutr. Food Res 2011, 55, 1361–1374. [Google Scholar]
- Poole, L.B.; Hall, A.; Nelson, K.J. Overview of peroxiredoxins in oxidant defense and redox regulation. Curr. Protoc. Toxicol 2011. [Google Scholar] [CrossRef]
- Chesler, E.J.; Lu, L.; Wang, J.; Williams, R.W.; Manly, K.F. WebQTL: Rapid exploratory analysis of gene expression and genetic networks for brain and behavior. Nat. Neurosci 2004, 7, 485–486. [Google Scholar]
- Wu, X.; Iguchi, T.; Itoh, N.; Okamoto, K.; Takagi, T.; Tanaka, K.; Nakanishi, T. Ascorbic acid transported by sodium-dependent vitamin C transporter 2 stimulates steroidogenesis in human choriocarcinoma cells. Endocrinology 2008, 149, 73–83. [Google Scholar]
- Zhu, J.; Sanborn, J.Z.; Diekhans, M.; Lowe, C.B.; Pringle, T.H.; Haussler, D. Comparative genomics search for losses of long-established genes on the human lineage. PLoS Comput. Biol 2007, 3, e247. [Google Scholar]
- Kloor, M.; Bork, P.; Duwe, A.; Klaes, R.; von Knebel Doeberitz, M.; Ridder, R. Identification and characterization of UEV3, a human cDNA with similarities to inactive E2 ubiquitin-conjugating enzymes. Biochim. Biophys. Acta 2002, 1579, 219–224. [Google Scholar]
- Kitamura, T.; Ido Kitamura, Y. Role of FoxO proteins in pancreatic beta cells. Endocr. J 2007, 54, 507–515. [Google Scholar]
- Liu, L.; Lu, Y.; Kong, H.; Li, L.; Marshall, C.; Xiao, M.; Ding, J.; Gao, J.; Hu, G. Aquaporin-4 deficiency exacerbates brain oxidative damage and memory deficits induced by long-term ovarian hormone deprivation and d-galactose injection. Int. J. Neuropsychopharmacol 2011, 1, 1–14. [Google Scholar]
- Yamamoto, S.; Takahashi, N.; Mori, Y. Chemical physiology of oxidative stress-activated TRPM2 and TRPC5 channels. Prog. Biophys. Mol. Biol 2010, 103, 18–27. [Google Scholar]
- Burkovics, P.; Hajdú, I.; Szukacsov, V.; Unk, I.; Haracska, L. Role of PCNA-dependent stimulation of 3′-phosphodiesterase and 3′–5′ exonuclease activities of human Ape2 in repair of oxidative DNA damage. Nucleic Acids Res 2009, 37, 4247–4255. [Google Scholar]
- Nakabeppu, Y.; Tsuchimoto, D.; Ichinoe, A.; Ohno, M.; Ide, Y.; Hirano, S.; Yoshimura, D.; Tominaga, Y.; Furuichi, M.; Sakumi, K. Biological significance of the defense mechanisms against oxidative damage in nucleic acids caused by reactive oxygen species: From mitochondria to nuclei. Ann. N. Y. Acad. Sci 2004, 1011, 101–111. [Google Scholar]
- Sanadgol, H.; Bayani, M.; Mohammadi, M.; Bayani, B.; Mashhadi, M.A. Effect of vitamin C on parathyroid hormone in hemodialysis patients with mild to moderate secondary hyperparathyroidism. Iran. J. Kidney Dis 2011, 5, 410–415. [Google Scholar]
- Ohta, Y.; Yashiro, K.; Kaida, S.; Imai, Y.; Ohashi, K.; Kitagawa, A. Water-immersion restraint stress disrupts nonenzymatic antioxidant defense systems through rapid and continuous ascorbic acid depletion in the adrenal gland of rats. Cell Biochem. Funct. 2012. [Google Scholar] [CrossRef]
- Ambali, S.F.; Orieji, C.; Abubakar, W.O.; Shittu, M.; Kawu, M.U. Ameliorative effect of vitamin C on alterations in thyroid hormones concentrations induced by subchronic coadministration of chlorpyrifos and lead in wistar rats. J. Thyroid Res 2011, 2011, 214924. [Google Scholar]
- Lima-Silva, V.; Rosado, A.; Amorin-Silva, V.; Muñoz-Mérida, A.; Pons, C.; Bombarely, A.; Trelles, O.; Fernández-Muñoz, R.; Granell, A.; Valpuesta, V.; et al. Genetic and genome-wide transcriptomic analyses identify co-regulation of oxidative response and hormone transcript abundance with vitamin c content in tomato fruit. BMC Genomics 2012, 13, 187. [Google Scholar]
- Yu, W.; Dittenhafer-Reed, K.E.; Denu, J.M. SIRT3 protein deacetylates isocitrate dehydrogenase 2 (IDH2) and regulates mitochondrial redox status. J. Biol. Chem 2012, 287, 14078–14086. [Google Scholar]
- Kim, H.K.; Chung, Y.W.; Chock, P.B.; Yim, M.B. Effect of CCS on the accumulation of FALS SOD1 mutant-containing aggregates and on mitochondrial translocation of SOD1 mutants: Implication of a free radical hypothesis. Arch. Biochem. Biophys 2011, 509, 177–185. [Google Scholar]
- Brigelius-Flohé, R.; Maiorino, M. Glutathione peroxidases. Biochim. Biophys. Acta 2012, 1830, 3289–3303. [Google Scholar]
- Tornavaca, O.; Pascual, G.; Barreiro, M.L.; Grande, M.T.; Carretero, A.; Riera, M.; Garcia-Arumi, E.; Bardaji, B.; González-Núñez, M.; Montero, M.A.; et al. Kidney androgen-regulated protein transgenic mice show hypertension and renal alterations mediated by oxidative stress. Circulation 2009, 119, 1908–1917. [Google Scholar]
- Bošković, M.; Vovk, T.; Saje, M.; Goričar, K.; Dolžan, V.; Kores Plesničar, B.; Grabnar, I. Association of SOD2, GPX1, CAT, and TNF genetic polymorphisms with oxidative stress, neurochemistry, psychopathology, and extrapyramidal symptoms in schizophrenia. Neurochem. Res 2012, 38, 433–442. [Google Scholar]
- Dokic, I.; Hartmann, C.; Herold-Mende, C.; Régnier-Vigouroux, A. Glutathione peroxidase 1 activity dictates the sensitivity of glioblastoma cells to oxidative stress. Glia 2012, 60, 1785–1800. [Google Scholar]
- Kaneko, H.; Anzai, T.; Morisawa, M.; Kohno, T.; Nagai, T.; Anzai, A.; Takahashi, T.; Shimoda, M.; Sasaki, A.; Maekawa, Y.; et al. Resveratrol prevents the development of abdominal aortic aneurysm through attenuation of inflammation, oxidative stress, and neovascularization. Atherosclerosis 2011, 217, 350–357. [Google Scholar]

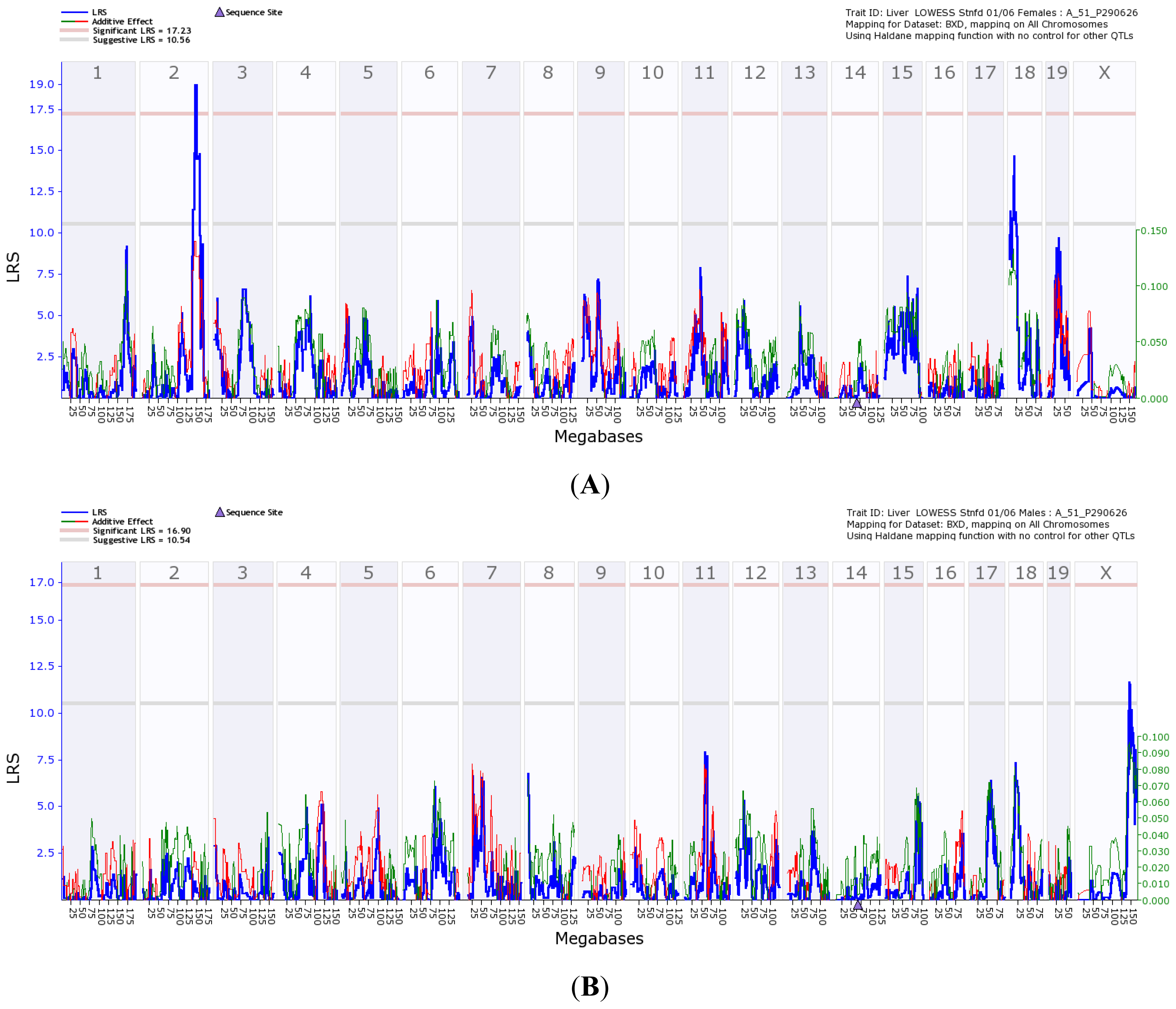
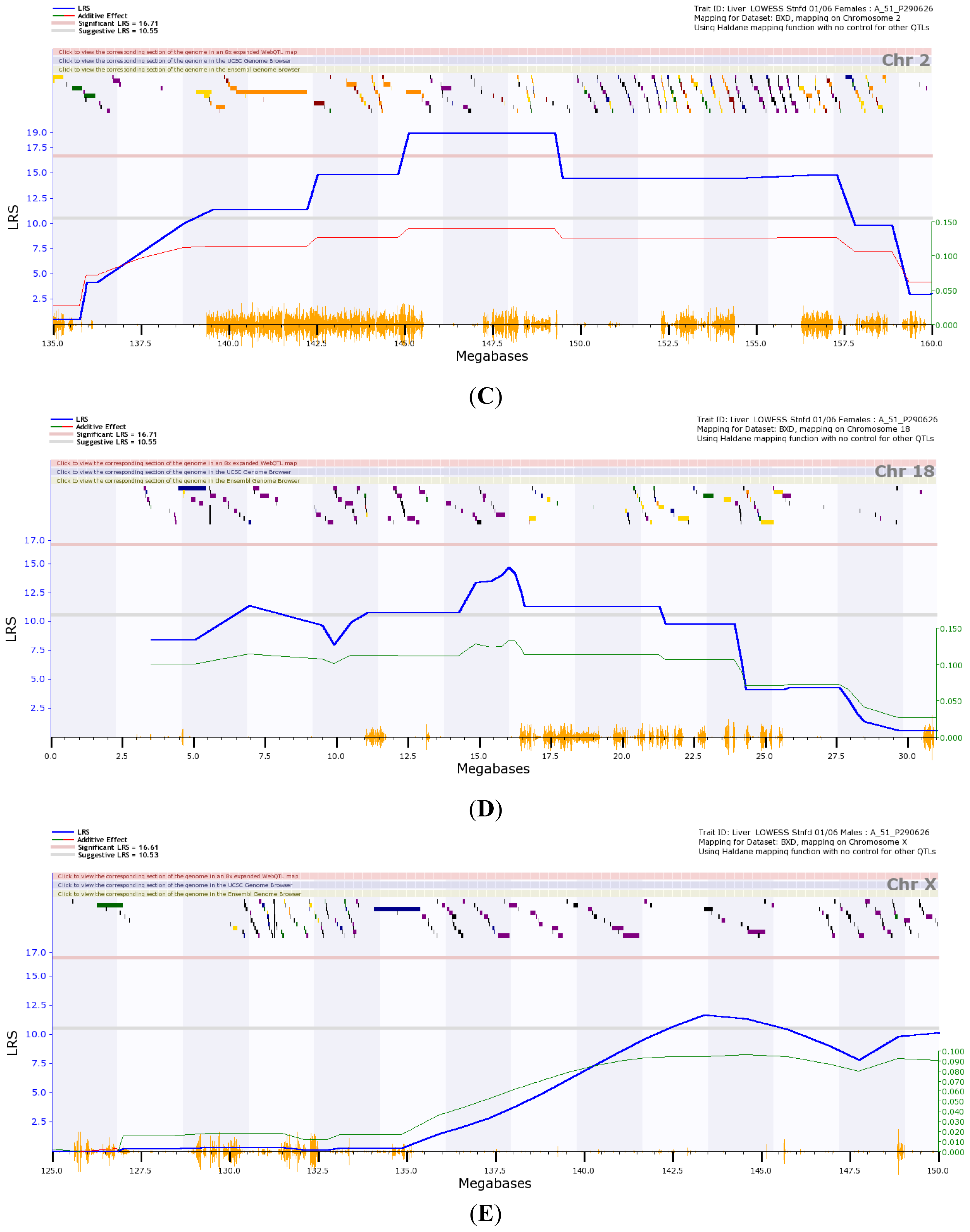
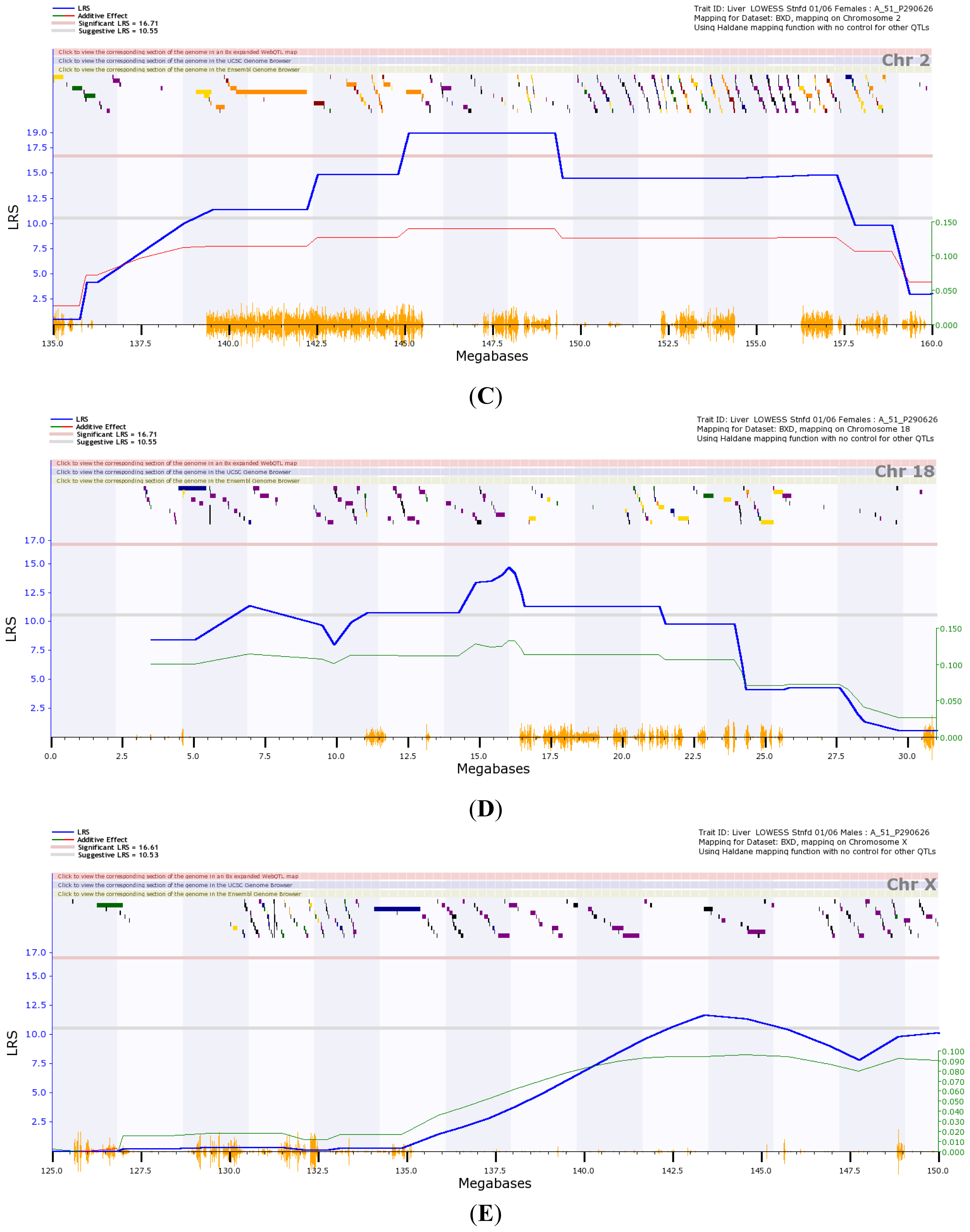
 ) is the likelihood ratio statistic (LRS) score. The sold pink line (
) is the likelihood ratio statistic (LRS) score. The sold pink line (
 ) indicates the level of significant LRS score, while the grey line (
) indicates the level of significant LRS score, while the grey line (
 ) indicates the suggestive LRS score. The green or red line (
) indicates the suggestive LRS score. The green or red line (
 ) is the additive score. (A) Transcriptome mapping of Gulo regulation QTL based on expression profiles generated from female strains, indicating that the two major loci located on Chr 2 and 18 regulate the expression of Gulo; (B) Transcriptome mapping of Gulo regulation QTL based on expression profiles generated from male strains, indicating that the major locus located on Chr X regulates the expression of Gulo; (C) The peak region of QTL on Chr 2 that regulates Gulo expression in female mice is located between 145 and 150 Mb; (D) The peak region of QTL on Chr 18 that regulates Gulo expression in female mice is located between 14 and 17 Mb; (E) The peak region of QTL on Chr X that regulates Gulo expression in male mice is located between 140 and 147 Mb.
) is the likelihood ratio statistic (LRS) score. The sold pink line (
) indicates the level of significant LRS score, while the grey line (
) indicates the suggestive LRS score. The green or red line (
) is the additive score. (A) Transcriptome mapping of Gulo regulation QTL based on expression profiles generated from female strains, indicating that the two major loci located on Chr 2 and 18 regulate the expression of Gulo; (B) Transcriptome mapping of Gulo regulation QTL based on expression profiles generated from male strains, indicating that the major locus located on Chr X regulates the expression of Gulo; (C) The peak region of QTL on Chr 2 that regulates Gulo expression in female mice is located between 145 and 150 Mb; (D) The peak region of QTL on Chr 18 that regulates Gulo expression in female mice is located between 14 and 17 Mb; (E) The peak region of QTL on Chr X that regulates Gulo expression in male mice is located between 140 and 147 Mb.
) is the additive score. (A) Transcriptome mapping of Gulo regulation QTL based on expression profiles generated from female strains, indicating that the two major loci located on Chr 2 and 18 regulate the expression of Gulo; (B) Transcriptome mapping of Gulo regulation QTL based on expression profiles generated from male strains, indicating that the major locus located on Chr X regulates the expression of Gulo; (C) The peak region of QTL on Chr 2 that regulates Gulo expression in female mice is located between 145 and 150 Mb; (D) The peak region of QTL on Chr 18 that regulates Gulo expression in female mice is located between 14 and 17 Mb; (E) The peak region of QTL on Chr X that regulates Gulo expression in male mice is located between 140 and 147 Mb.
) is the likelihood ratio statistic (LRS) score. The sold pink line (
) indicates the level of significant LRS score, while the grey line (
) indicates the suggestive LRS score. The green or red line (
) is the additive score. (A) Transcriptome mapping of Gulo regulation QTL based on expression profiles generated from female strains, indicating that the two major loci located on Chr 2 and 18 regulate the expression of Gulo; (B) Transcriptome mapping of Gulo regulation QTL based on expression profiles generated from male strains, indicating that the major locus located on Chr X regulates the expression of Gulo; (C) The peak region of QTL on Chr 2 that regulates Gulo expression in female mice is located between 145 and 150 Mb; (D) The peak region of QTL on Chr 18 that regulates Gulo expression in female mice is located between 14 and 17 Mb; (E) The peak region of QTL on Chr X that regulates Gulo expression in male mice is located between 140 and 147 Mb.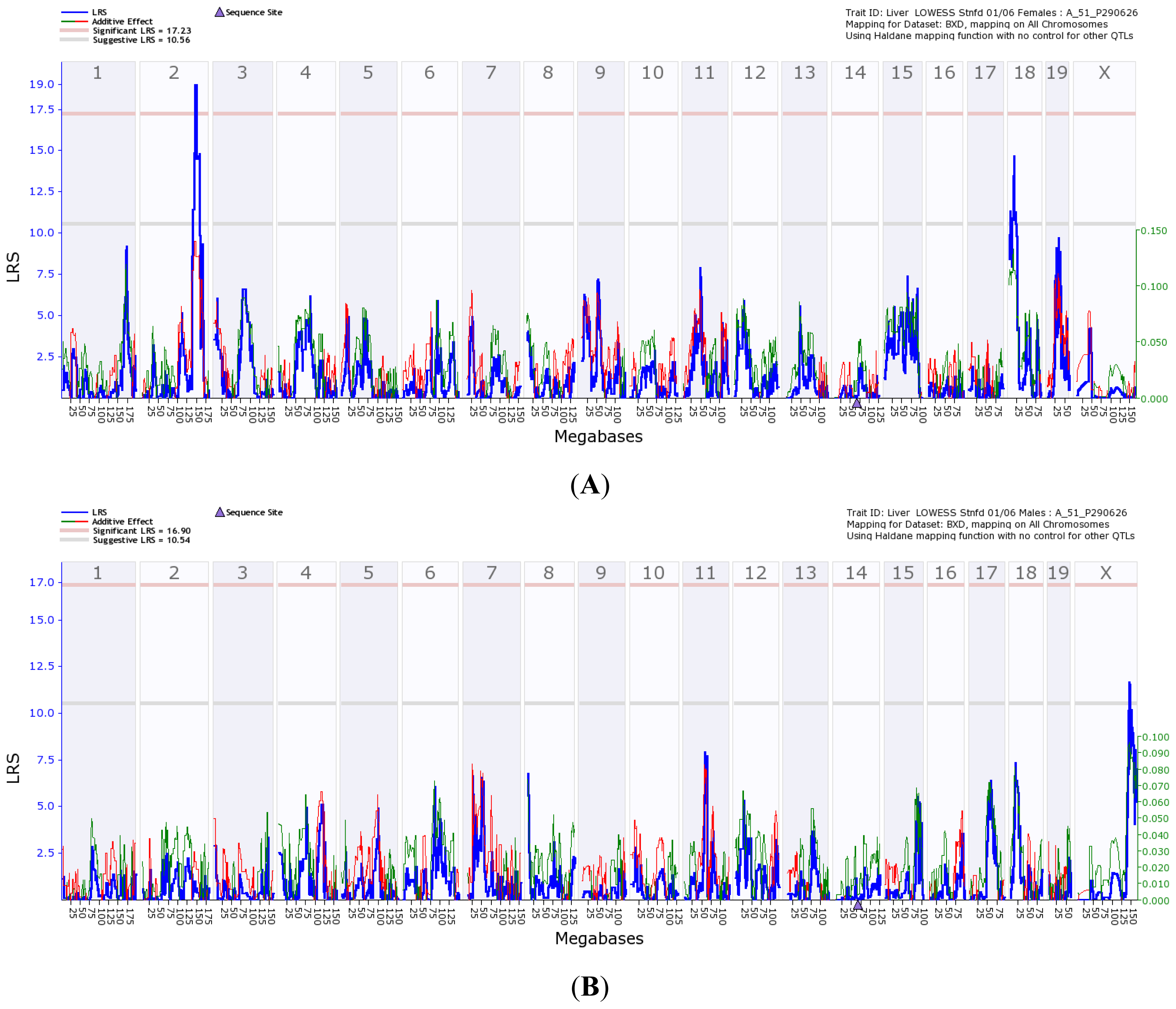
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Jiao, Y.; Chen, H.; Yan, J.; Wang, L.; Huang, Y.; Liu, X.; Williams, R.W.; Lu, L.; Wang, Y.; Gu, W. Genome-Wide Gene Expression Profiles in Antioxidant Pathways and Their Potential Sex Differences and Connections to Vitamin C in Mice. Int. J. Mol. Sci. 2013, 14, 10042-10062. https://doi.org/10.3390/ijms140510042
Jiao Y, Chen H, Yan J, Wang L, Huang Y, Liu X, Williams RW, Lu L, Wang Y, Gu W. Genome-Wide Gene Expression Profiles in Antioxidant Pathways and Their Potential Sex Differences and Connections to Vitamin C in Mice. International Journal of Molecular Sciences. 2013; 14(5):10042-10062. https://doi.org/10.3390/ijms140510042
Chicago/Turabian StyleJiao, Yan, Hong Chen, Jian Yan, Lishi Wang, Yue Huang, Xiaoyun Liu, Robert W. Williams, Lu Lu, Yongjun Wang, and Weikuan Gu. 2013. "Genome-Wide Gene Expression Profiles in Antioxidant Pathways and Their Potential Sex Differences and Connections to Vitamin C in Mice" International Journal of Molecular Sciences 14, no. 5: 10042-10062. https://doi.org/10.3390/ijms140510042
APA StyleJiao, Y., Chen, H., Yan, J., Wang, L., Huang, Y., Liu, X., Williams, R. W., Lu, L., Wang, Y., & Gu, W. (2013). Genome-Wide Gene Expression Profiles in Antioxidant Pathways and Their Potential Sex Differences and Connections to Vitamin C in Mice. International Journal of Molecular Sciences, 14(5), 10042-10062. https://doi.org/10.3390/ijms140510042