Combining Hexanoic Acid Plant Priming with Bacillus thuringiensis Insecticidal Activity against Colorado Potato Beetle

 , and
, and
Abstract
:
1. Introduction
2. Results and Discussion
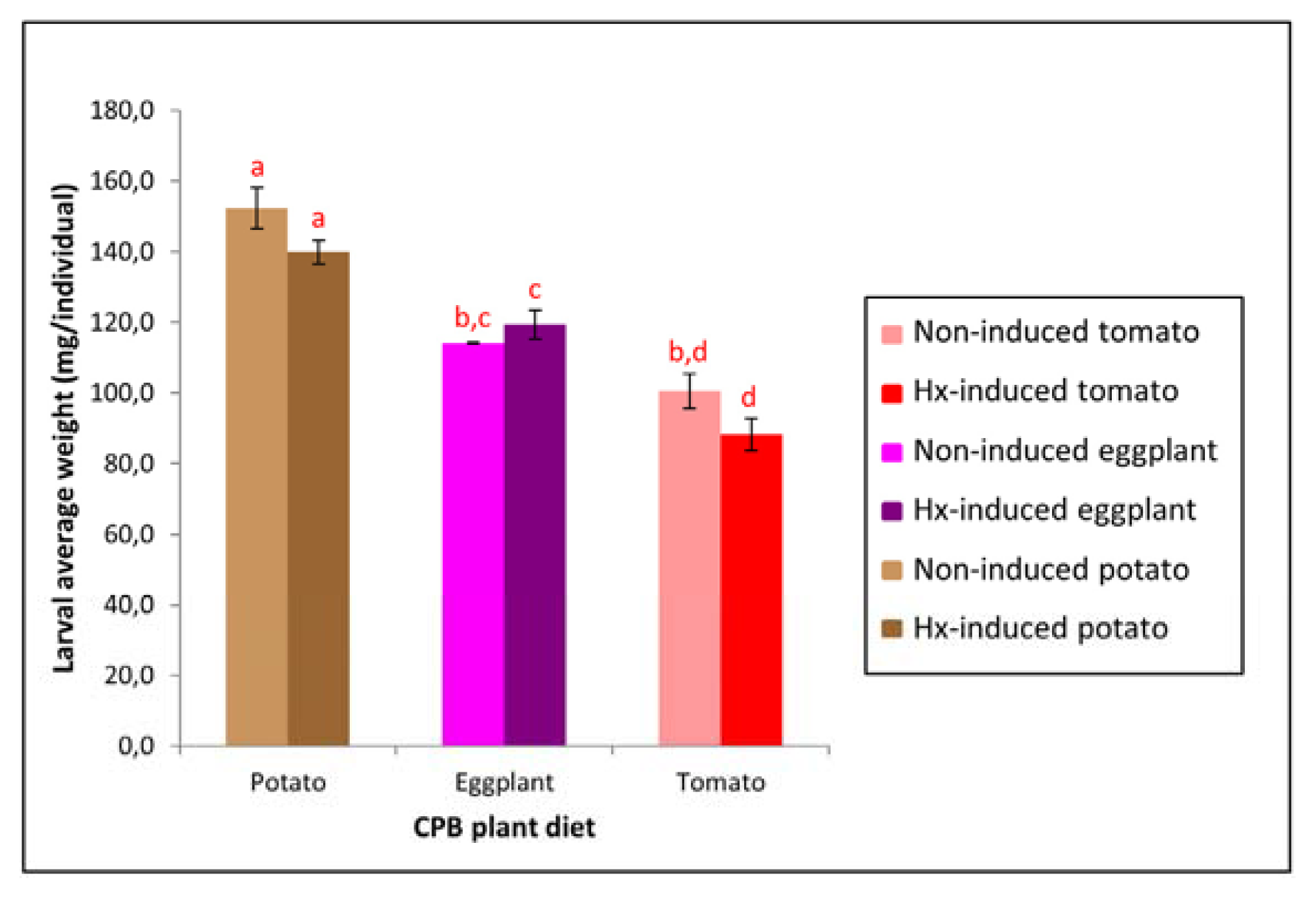
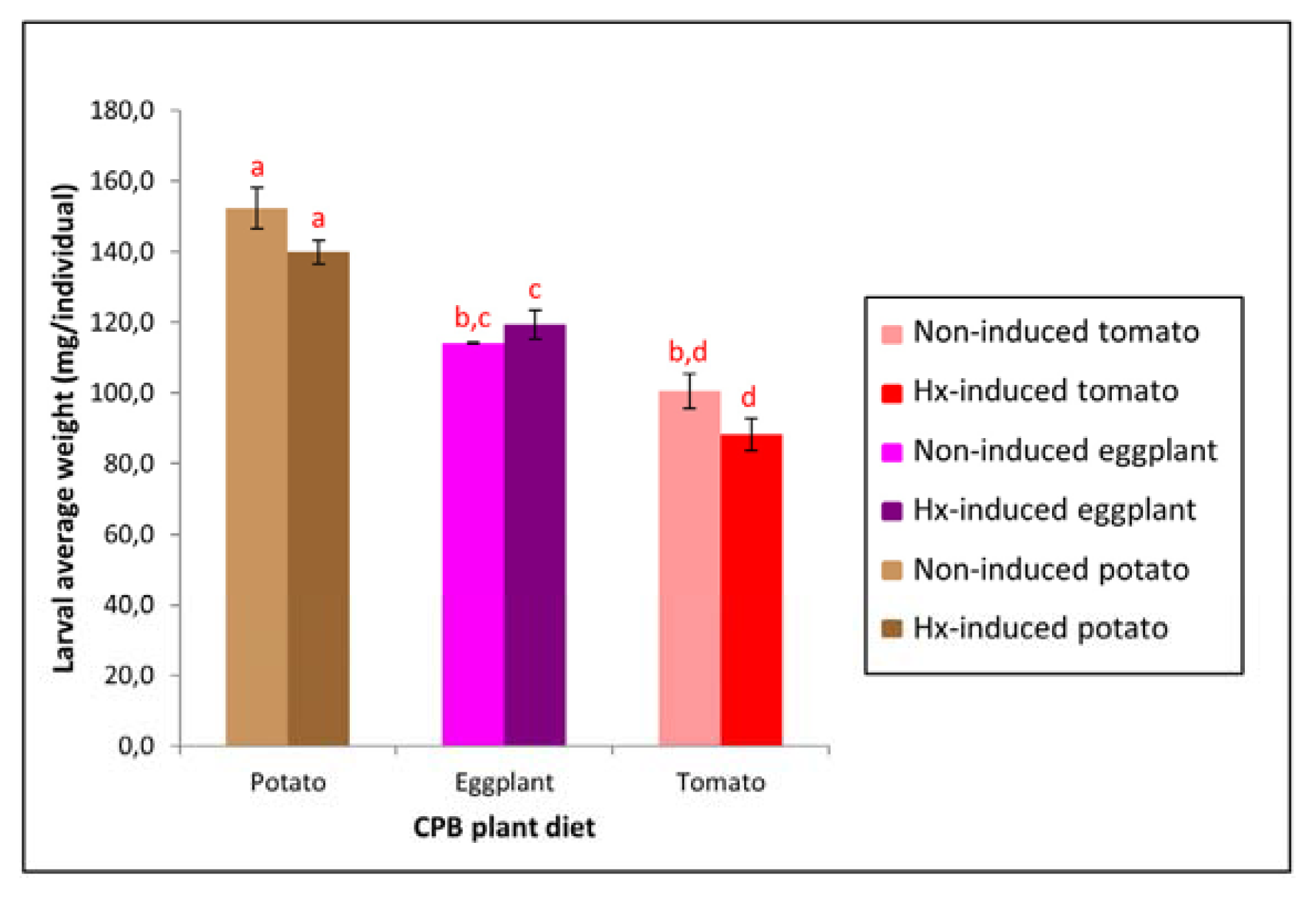
2.1. Host Plant Modifies the Growth of CPB Larvae, Affecting their Susceptibility to Bt Cry3Aa Toxin
2.2. Effect of CPB Larvae Diet on Cry3Aa Toxin Proteolysis
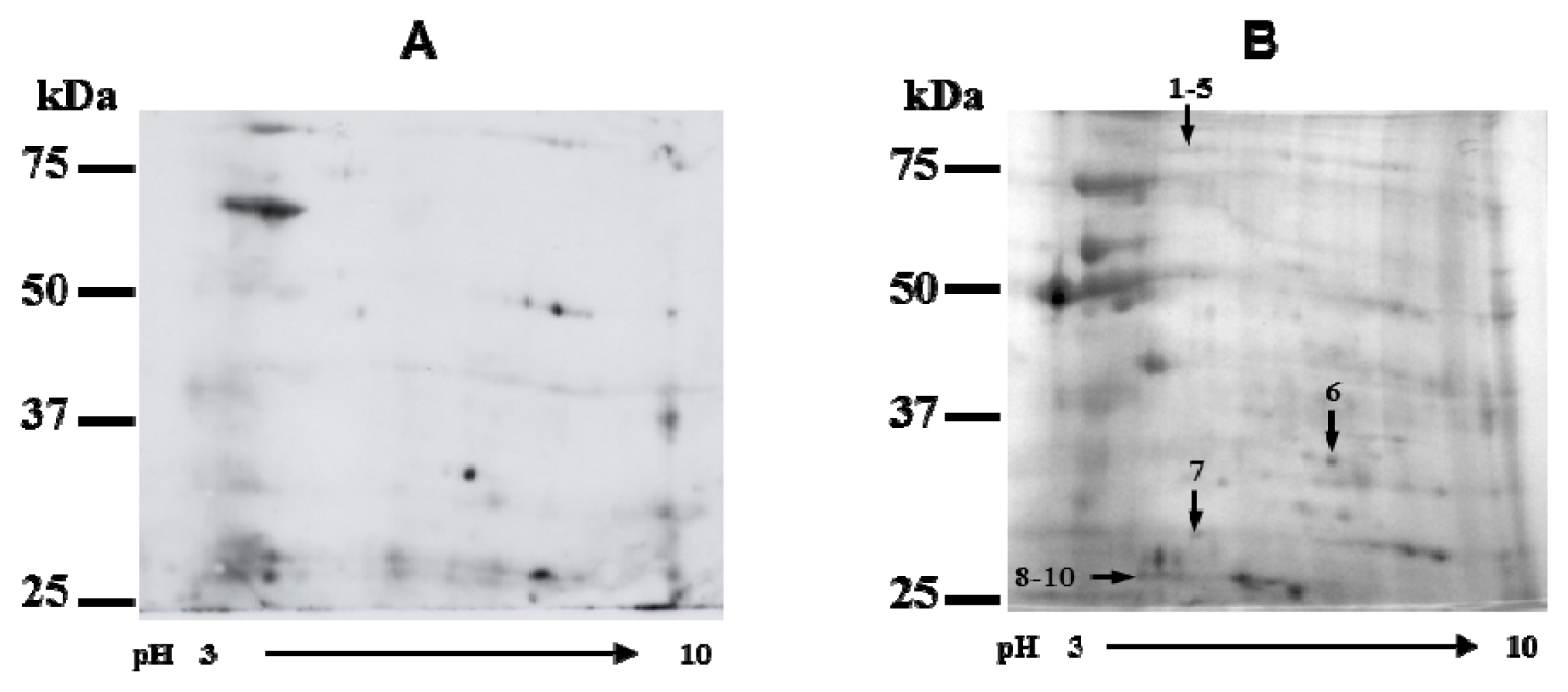
2.3. Identification of Cry3Aa Interacting Cysteine Proteases
2.4. Analysis of Papain Type Cysteine Protease Activity in CPB Larvae Fed on Different Host Plants
2.5. Effect of Plant Hx Treatment on Cry3Aa Toxin Insecticidal Action against CPB Larvae
2.5.1. Cry3Aa Mortality of CPB Larvae Fed on Three Hx-induced Solanaceous Plants
2.5.2. Feeding Performance of CPB Larvae Fed on Three Hx-Induced Solanaceous Plants
2.5.3. Cysteine Protease Activities on CPB Larvae fed on three Hx-induced Solanaceous Plants
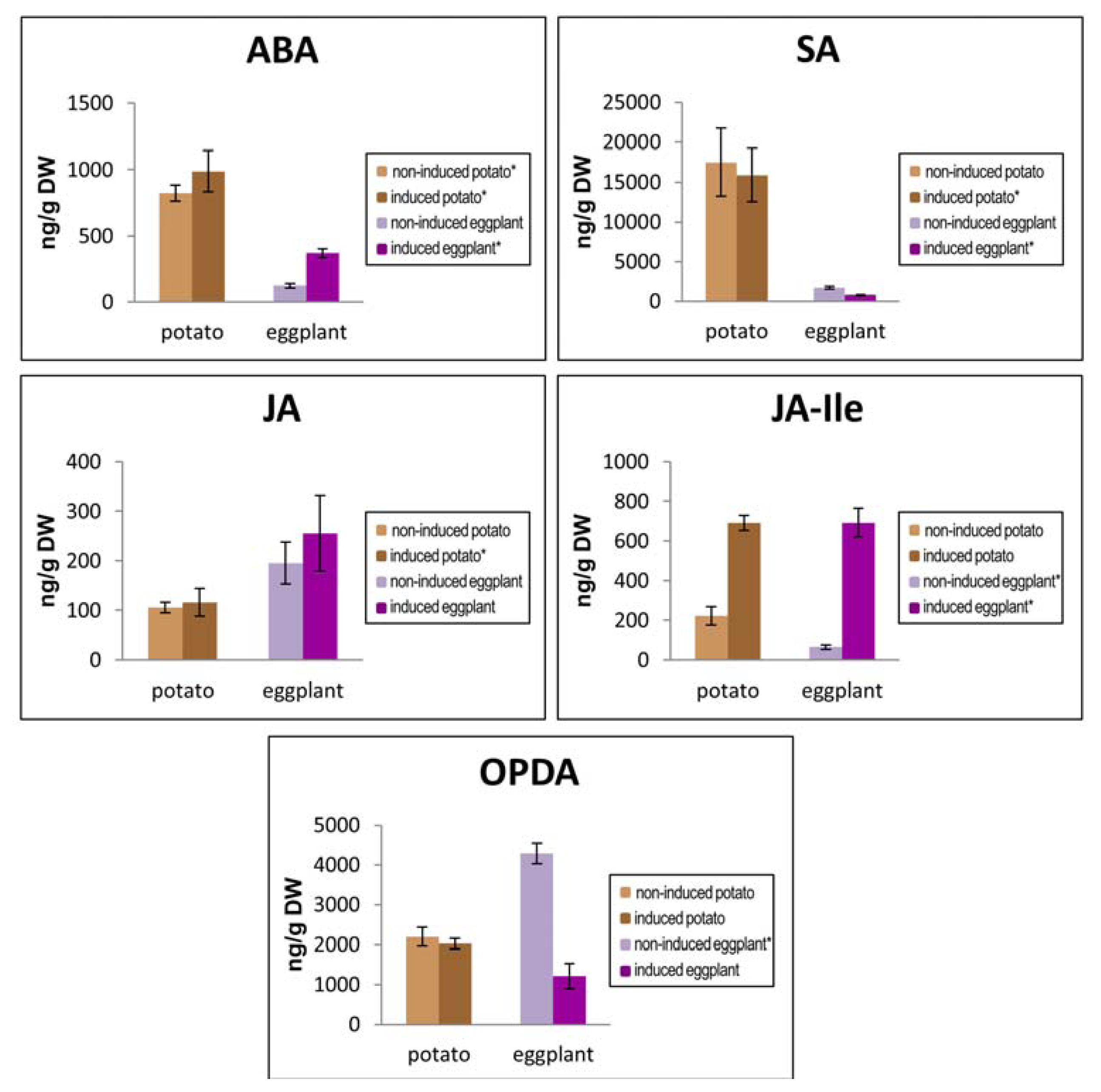
2.6. Study of the Hormonal Pattern in Eggplant and Potato Plants Following Hx Induction
3. Experimental Section
3.1. Plants
3.2. Insects
3.3. Hexanoic Acid Plant Defense Induction
3.4. Preparation of Brush Border Membrane Vesicles (BBMV)
3.5. Toxin Purification
3.6. Cry3Aa Toxin Cleavage Assays
3.7. Insect Toxicity Assay
3.8. Determination of ABA, SA, OPDA, JA and JA-Ile Levels
3.9. Photoactive Biotin Label Transfer Assays
3.10. Papain Type Cysteine Protease Assays in CPB Larvae fed on Different Host Plants
4. Conclusions
Acknowledgments
Conflict of Interest
References
- Alyokhin, A. Colorado Potato Beetle Management on Potatoes: Current Challenges and Future Prospects. In Potato II. Fruit, Vegetable and Cereal Science and Biotechnology 3 (Special Issue 1); Tennant, P., Benkeblia, N., Eds.; GSB Publisher: Isleworth, UK, 2004; pp. 10–19. [Google Scholar]
- Hitchner, E.M.; Kuhar, T.P.; Dickens, J.C.; Youngman, R.R.; Schultz, P.B.; Pfeiffer, D.G. Host plant choice experiments of colorado potato beetle (Coleoptera: Chrysomelidae) in Virginia. J. Econ. Entomol 2008, 101, 859–865. [Google Scholar]
- Walters, D.; Walsh, D.; Newton, A.; Lyon, G. Induced resistance for plant disease control: Maximizing the efficacy of resistance elicitors. Phytopathology 2005, 95, 1368–1373. [Google Scholar]
- Bravo, A.; Gill, S.S.; Soberon, M. Mode of action of Bacillus thuringiensis Cry and Cyt toxins and their potential for insect control. Toxicon 2007, 49, 423–435. [Google Scholar]
- Zhang, X.; Candas, M.; Griko, N.B.; Taussig, R.; Bulla, L.A., Jr. A mechanism of cell death involving an adenylyl cyclase/PKA signaling pathway is induced by the Cry1Ab toxin of Bacillus thuringiensis. Proc. Natl. Acad. Sci. USA 2006, 103, 9897–9902. [Google Scholar]
- Mohan, S.; Ma, P.W.K.; Williams, W.P.; Luthe, D.S. A naturally occurring plant cysteine protease possesses remarkable toxicity against insect pests and synergizes Bacillus thuringiensis toxin. PLoS One 2008, 3, e1786. [Google Scholar]
- Rivard, D.; Cloutier, C.; Michaud, D. Colorado potato beetles show differential digestive compensatory responses to host plants expressing distinct sets of defense proteins. Arch. Insect Biochem. Physiol 2004, 55, 114–123. [Google Scholar]
- Cory, J.S.; Hoover, K. Plant-mediated effects in insect-pathogen interactions. Trends Ecol. Evol 2006, 21, 278–286. [Google Scholar]
- Bari, R.; Jones, J.D.G. Role of plant hormones in defence responses. Plant Mol. Biol 2009, 69, 473–488. [Google Scholar]
- Vicedo, B.; Flors, V.; Leyva, M.O.; Finiti, I.; Kravchuk, Z.; Real, M.D.; García-Agustín, P.; González-Bosch, C. Hexanoic acid induced resistance against Botrytis cinerea in tomato plants. Mol. Plant Microbe Interact 2009, 22, 1455–1465. [Google Scholar]
- Kravchuk, Z.; Vicedo, B.; Flors, V.; Camanes, G.; Gonzalez-Bosch, C.; Garcia-Agustin, P. Priming for JA-dependent defenses using hexanoic acid is an effective mechanism to protect Arabidopsis against B. cinerea. J. Plant Physiol 2011, 168, 359–366. [Google Scholar]
- Scalschi, L.; Vicedo, B.; Camañes, G.; Fernández-Crespo, E.; Lapeña, L.; González-Bosch, C.; García-Agustín, P. Hexanoic acid is a resistance inducer that protects tomato plants against Pseudomonas syringae by priming the jasmonic acid and salicylic acid pathways. Mol. Plant Pathol 2013, 14, 342–355. [Google Scholar]
- Llorens, E.; Fernández-Crespo, E.; Vicedo, B.; Lapeña, L.; García-Agustín, P. Enhancement of the citrus immune system provides effective resistance against Alternaria brown spot disease. J. Plant Physiol 2013, 170, 146–154. [Google Scholar]
- Wasternack, C. Jasmonates: An update on biosynthesis, signal transduction and action in plant stress response, growth and development. Ann. Bot 2007, 100, 681–697. [Google Scholar]
- Howe, G.A.; Jander, G. Plant immunity to insect herbivores. Annu. Rev. Plant Biol 2008, 59, 41–66. [Google Scholar]
- Browse, J. Jasmonate passes muster: A receptor and targets for the defence hormone. Annu. Rev. Plant Biol 2009, 60, 183–205. [Google Scholar]
- Wu, J.; Baldwin, I.T. Herbivory-induced signalling in plants: Perception and action. Plant Cell Environ 2009, 32, 1161–1174. [Google Scholar]
- Seo, H.S.; Song, J.T.; Cheong, J.J.; Lee, Y.H.; Lee, Y.W.; Hwang, I.; Lee, J.S.; Choi, Y.D. Jasmonic acid carboxyl methyltransferase: A key enzyme for jasmonate regulated plant responses. Proc. Natl. Acad. Sci. USA 2001, 98, 4788–4793. [Google Scholar]
- Staswick, P.E.; Tiryaki, I. The oxylipin signal jasmonic acid is activated by an enzyme that conjugates it to isoleucine in Arabidopsis. Plant Cell 2004, 16, 2117–2127. [Google Scholar]
- Staswick, P.E.; Tiryaki, I.; Rowe, M.L. Jasmonate response locus JAR1 and several related Arabidopsis genes encode enzymes of the firefly luciferase superfamily that show activity on jasmonic, salicylic, and indole-3-acetic acids in an assay for adenylation. Plant Cell 2002, 14, 1405–1415. [Google Scholar]
- Wang, L.; Allmann, S.; Wu, J.; Baldwin, I.T. Comparisons of LIPOXYGENASE3- and JASMONATE-RESISTANT4/6-silenced plants reveal that jasmonic acid and jasmonic acid-amino acid conjugates play different roles in herbivore resistance of Nicotiana attenuata. Plant Physiol 2008, 146, 904–915. [Google Scholar]
- Overney, S.; Fawe, A.; Yelle, S.; Michaud, D. Diet-related plasticity of the digestive proteolytic system in larvae of the Colorado Potato Beetle (Leptinotarsa decemlineata Say). Arch. Insect Biochem. Physiol 1997, 36, 241–250. [Google Scholar]
- Kouassi, K.C.; Lorenzetti, F.; Guertin, C.; Cabana, J.; Mauffette, Y. Variation in the susceptibility of the forest tent caterpillar (Lepidoptera: Lasiocampidae) to Bacillus thuringiensis variety kurstaki HD-1: Effect of the host plant. J. Econ. Entomol 2001, 94, 1135–1141. [Google Scholar]
- Janmaat, A.F.; Myers, J.H. Host-plant effects the expression of resistance to Bacillus thuringiensis kurstaki in Trichoplusia ni (Hubner): An important factor in resistance evolution. J. Evol. Biol 2007, 20, 62–69. [Google Scholar]
- Novillo, C.; Castañera, P.; Ortego, F. Characterization and distribution of chymotrypsin-like and other digestive proteases in Colorado potato beetle larvae. Arch. Insect Biochem. Physiol 1997, 36, 181–201. [Google Scholar]
- Petek, M.; Turnsek, N.; Buh Gasparic, M.; Pompe Novak, M.; Gruden, K.; Slapar, N.; Popovic, T.; Strukelj, B.; Jongsma, M.A. A complex of genes involved in adaptation of Leptinotarsa decemlineata larvae to induced potato defense. Arch. Insect Biochem. Physiol 2012, 79, 153–181. [Google Scholar]
- Loseva, O.; Ibrahim, M.; Candas, M.; Koller, N.C.; Bauer, L.S.; Bulla, L.A., Jr. Changes in protease activity and Cry3A toxin binding in the Colorado potato beetle: Implications for insect resistance to Bacillus thuringiensis toxins. Insect Biochem. Mol. Biol 2002, 32, 567–577. [Google Scholar]
- Oppert, B.; Kramer, K.J.; Beeman, R.W.; Johnson, D.; McGaughey, W.H. Proteinase-mediated insect resistance to Bacillus thuringiensis toxins. J. Biol. Chem 1997, 272, 23473–23476. [Google Scholar]
- Forcada, C.; Alcacer, E.; Garcera, M.D.; Martinez, R. Differences in the midgut proteolytic activity of two Heliothis virescens strains, one susceptible and one resistant to Bacillus thuringiensis toxins. Arch. Insect Biochem. Physiol 1996, 3, 1257–1272. [Google Scholar]
- Rajagopal, R.; Arora, N.; Sivakumar, S.; Rao, N.G.V.; Nimbalkar, S.A.; Bhatnagar, R.K. Resistance of Helicoverpa armigera to Cry1Ac toxin from Bacillus thuringiensis is due to improper processing of the protoxin. Biochem. J 2009, 419, 309–316. [Google Scholar]
- Tetreau, G.; Stalinski, R.; David, J.P.; Despres, L. Increase in larval gut proteolytical activities and Bti resistance in the dengue fever mosquito. Arch. Insect Biochem. Physiol 2013, 82, 71–83. [Google Scholar]
- Walters, F.S.; Stacy, C.M.; Lee, M.K.; Palekar, N.; Chen, J.S. An engineered chymotrypsin/cathepsin G site domain I renders Bacillus thuringiensis Cry3A active against western corn rootworm larvae. Appl. Environ. Microbiol 2008, 74, 367–374. [Google Scholar]
- Gruden, K.; Popovic, T.; Cimerman, N.; Krizaj, I.; Strukelj, B. Diverse enzymatic specificities of digestive proteases, “intestains”, enable Colorado potato beetle larvae to counteract the potato defence mechanism. Biol. Chem 2003, 384, 305–310. [Google Scholar]
- Sainsbury, F.; Rheáume, A.J.; Goulet, M.C.; Vorster, J.; Michaud, D. Discrimination of differentially inhibited cysteine proteases by activity-based profiling using cystatin variants with tailored specificities. J. Proteome Res 2012, 11, 5983–5993. [Google Scholar]
- Gruden, K.; Kuipers, A.G.; Guncar, G.; Slapar, N.; Strukelj, B.; Jongsma, M.A. Molecular basis of Colorado potato beetle adaptation to potato plant defence at the level of digestive cysteine proteinases. Insect Biochem. Mol. Biol 2004, 34, 365–375. [Google Scholar]
- Clustal omega. Available online: http://www.ebi.ac.uk/Tools/msa/clustalo (accessed on 15 April 2013).
- Bolter, C.J.; Jongsma, M.A. Colorado potato beetles (Leptinotarsa decemlineata) adapt to proteinase inhibitors induced in potato leaves by methyl jasmonate. Insect. Physiol 1995, 41, 1071–1078. [Google Scholar]
- Abo-Elyousr, K.A.M.; Ibrahim, Y.E.; Balabel, N.M. Induction of disease defensive enzymes in response to treatment with acibenzolar-S-methyl (ASM) and Pseudomonas fluorescens Pf2 and inoculation with Ralstonia solanacearum race 3, biovar 2 (phylotype II). J. Phytopathol 2012, 160, 382–389. [Google Scholar]
- Myresiotis, C.K.; Karaoglanidis, G.S.; Vryzas, Z.; Papadopoulou-Mourkidou, E. Evaluation of plant growth promoting rhizobacteria, acibenzolar-S-methyl and hymexol for integrated control of Fusarium crown and root rot on tomato. Pest. Manag. Sci 2012, 68, 404–411. [Google Scholar]
- Gurr, G.M.; Kvedaras, O.L. Synergizing biological control: Scope for sterile insect technique, induced plant defences and cultural techniques to enhance natural enemy impact. Biol. Control 2010, 52, 198–207. [Google Scholar]
- Ton, J.; D’Alessandro, M.; Jourdie, V.; Jakab, G.; Karlen, D.; Held, M.; Mauch-Mani, B.; Turlings, T.C.J. Priming by airborne signals boosts direct and indirect resistance in maize. Plant J 2007, 49, 16–26. [Google Scholar]
- Conrath, U.; Beckers, G.J.M.; Flors, V.; García-Agustín, P.; Jakab, G.; Mauch, F.; Newman, M.A.; Pieterse, C.M.J.; Poinssot, B.; Pozo, M.J.; et al. Priming: Getting ready for battle. Mol. Plant Microbe Interact 2006, 19, 1062–1071. [Google Scholar]
- Wolfesberger, M.; Lüthy, P.; Maurer, A.; Parenti, F.; Sacchi, V.; Giordana, B.; Hanozet, G.M. Preparation and partial characterization of amino acid transporting brush border membrane vesicles form the larval midgut of the cabbage butterfly (Pieris brassicae). Comp. Biochem. Physiol 1987, 86A, 301–308. [Google Scholar]
- Reuveni, M.; Dunn, P.E. Differential inhibition by Bacillus thuringiensis delta endotoxin of leucine and aspartic uptake in BBMV from midgut of Manduca sexta. Biochem. Biophys. Res. Commun 1991, 181, 1089–1093. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem 1976, 72, 248–254. [Google Scholar]
- Thomas, W.E.; Ellar, D.J. Bacillus thuringiensis var. israeliensis crystal delta-endotoxin: Effects on insect and mammalian cells in vitro and in vivo. J. Cell Sci 1983, 60, 181–197. [Google Scholar]
- Rausell, C.; Ochoa-Campuzano, C.; Martínez-Ramírez, A.C.; Bravo, A.; Real, M.D. A membrane associated metalloprotease cleaves Cry3Aa Bacillus thuringiensis toxin reducing pore formation in Colorado potato beetle brush border membrane vesicles. Biochim. Biophys. Acta 2007, 1768, 2293–2299. [Google Scholar]
- Pinfield-Wells, H.; Rylott, E.L.; Gilday, A.D.; Graham, S.; Job, K.; Larson, T.R.; Graham, I.A. Sucrose rescues seedling establishment but not germination of Arabidopsis mutants disrupted in peroxisomal fatty acid catabolism. Plant J 2005, 43, 861–872. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CPB plant diet | Weight (mg/individual) | % relative weight increase * | Cry3Aa mortality (%) | |
|---|---|---|---|---|
| Day 0 | Day 7 | |||
| Potato | 50.3 ± 0.8 a | 152.4 ± 5.8 | 203 | 61 ± 5 |
| Eggplant | 51.6 ± 2.1 a | 114.1 ± 0.2 b | 121 | 79 ± 8 |
| Tomato | 51.2 ± 0.6 a | 100.5 ± 4.8 b | 96 | 89 ± 6 |
| No. | NCBI accession No. | Name | Species | Mascot score | MW (kDa) | No. identified peptides |
|---|---|---|---|---|---|---|
| 1 | XP_974860 | Aconitase | Tribolium castaneum | 140 | 86.23 | 29 |
| 2 | XP_001663037 | Aconitase | Aedes aegypti | 154 | 86.41 | 29 |
| 3 | XP_974860 | Aconitase | T. castaneum | 120 | 86.23 | 29 |
| 4 | XP_317642 | Aconitase | Anopheles gambiae | 134 | 85.92 | 27 |
| 5 | XP_317642 | Aconitase | A. gambiae | 162 | 85.92 | 26 |
| 6 | XP_624353 | Aldo-keto reductase | Apis mellifera | 124 | 36.46 | 5 |
| 7 | XP_974606 | Prohibitin-1 | T. castaneum | 280 | 30.32 | 11 |
| 8 | ABM55486 | Intestain D-6 | Leptinotarsa decemlineata | 86 | 36.75 | 12 |
| ABM55485 | Intestain D-5 | L. decemlineata | 86 | 36.76 | 12 | |
| 9 | AAN77410 | Intestain B-2 | L. decemlineata | 76 | 22.33 | 6 |
| AAS20591 | Intestain B-11 | L. decemlineata | 73 | 36.67 | 6 | |
| AAS20592 | Intestain B-12 | L. decemlineata | 73 | 36.65 | 6 | |
| 10 | ABM55486 | Intestain D-6 | L. decemlineata | 206 | 36.75 | 20 |
| ABM55485 | Intestain D-5 | L. decemlineata | 206 | 36.75 | 20 | |
| ABM55484 | Intestain D-4 | L. decemlineata | 145 | 36.80 | 18 | |
| ABM55491 | Intestain D-11 | L. decemlineata | 126 | 10.88 | 9 | |
| ABM55488 | Intestain D-8 | L. decemlineata | 124 | 10.34 | 9 | |
| ABM55495 | Intestain D-15 | L. decemlineata | 87 | 29.74 | 13 |
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
García-Robles, I.; Ochoa-Campuzano, C.; Fernández-Crespo, E.; Camañes, G.; Martínez-Ramírez, A.C.; González-Bosch, C.; García-Agustín, P.; Rausell, C.; Real, M.D. Combining Hexanoic Acid Plant Priming with Bacillus thuringiensis Insecticidal Activity against Colorado Potato Beetle. Int. J. Mol. Sci. 2013, 14, 12138-12156. https://doi.org/10.3390/ijms140612138
García-Robles I, Ochoa-Campuzano C, Fernández-Crespo E, Camañes G, Martínez-Ramírez AC, González-Bosch C, García-Agustín P, Rausell C, Real MD. Combining Hexanoic Acid Plant Priming with Bacillus thuringiensis Insecticidal Activity against Colorado Potato Beetle. International Journal of Molecular Sciences. 2013; 14(6):12138-12156. https://doi.org/10.3390/ijms140612138
Chicago/Turabian StyleGarcía-Robles, Inmaculada, Camila Ochoa-Campuzano, Emma Fernández-Crespo, Gemma Camañes, Amparo C. Martínez-Ramírez, Carmen González-Bosch, Pilar García-Agustín, Carolina Rausell, and María Dolores Real. 2013. "Combining Hexanoic Acid Plant Priming with Bacillus thuringiensis Insecticidal Activity against Colorado Potato Beetle" International Journal of Molecular Sciences 14, no. 6: 12138-12156. https://doi.org/10.3390/ijms140612138