The Role of Bacterial Biofilms and Surface Components in Plant-Bacterial Associations

Abstract
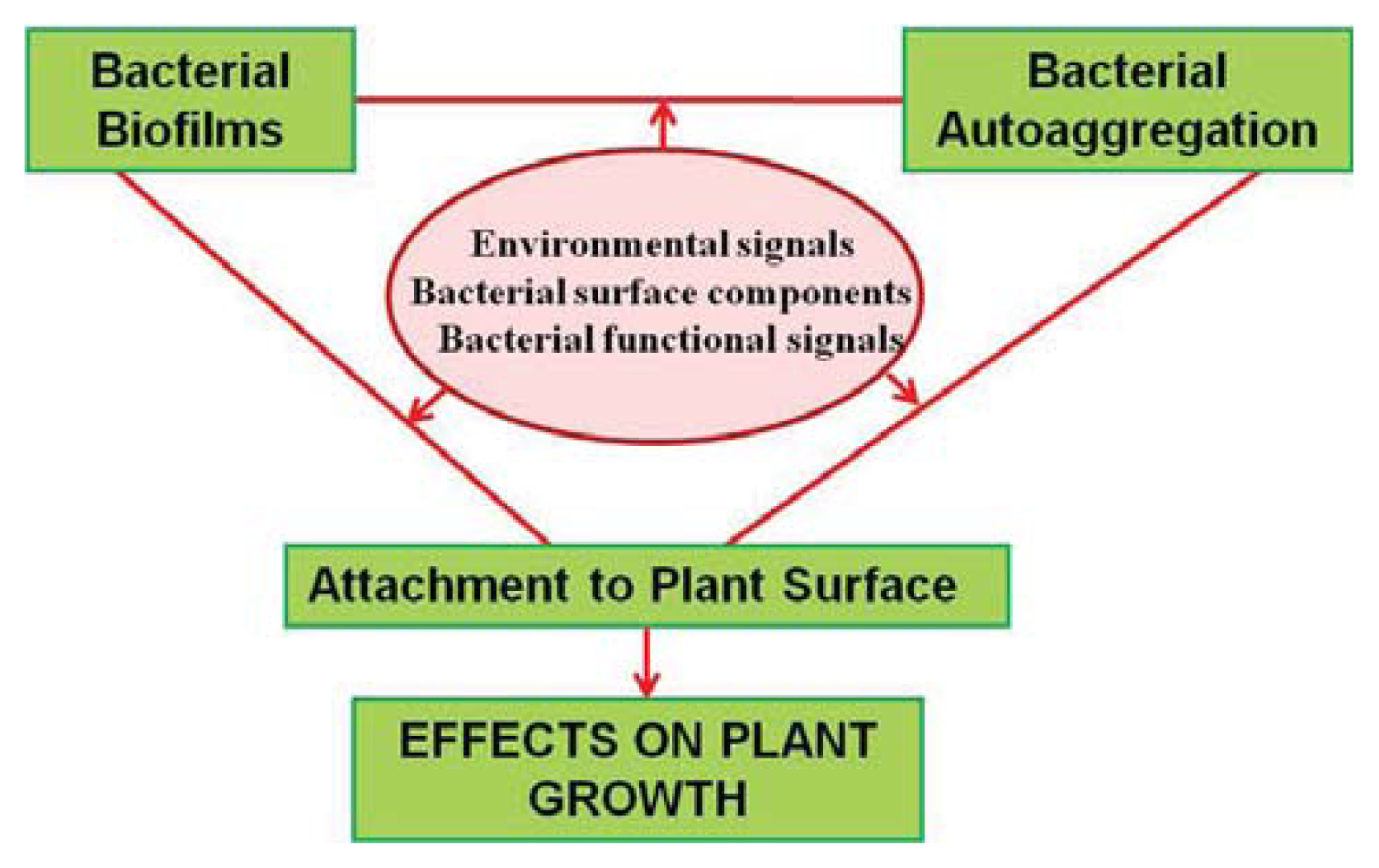
:1. Introduction
2. Cell–Cell Adhesive Interactions: Bacterial Autoaggregation
2.1. Surface Factors Involved in Bacterial Autoaggregation
2.2. Extracellular Factors Involved in Bacterial Autoaggregation
3. Cell–Cell and Cell–Surface Interactions: Bacterial Biofilm Formation
3.1. Structural and Functional Components Involved in Biofilm Formation
3.1.1. Surface Bacterial Factors
3.1.2. Extracellular Factors
4. Relationship between Biofilm Formation and Bacterial Autoaggregation
5. Intergeneric Adhesive Interactions: Coaggregation
6. Concluding Remarks
Acknowledgments
Conflict of Interest
References
- Branda, S.S.; Vik, S.; Friedman, L.; Kolter, R. Biofilms: The matrix revisited. Trends Microbiol 2005, 13, 20–26. [Google Scholar]
- Davey, M.E.; O’Toole, G.A. Microbial biofilms: From ecology to molecular genetics. Microbiol. Mol. Biol. Rev 2000, 64, 847–867. [Google Scholar]
- Schembri, M.A.; Christianesen, G.; Klemm, P. FimH-mediated autoaggregation of E. coli. Mol. Microbiol 2001, 41, 1419–1430. [Google Scholar]
- Costerton, J.W.; Lewandowski, Z.; Caldwell, D.E.; Korber, D.R.; Lappin-Scott, H.M. Microbial biofilms. Annu. Rev. Microbiol 1995, 49, 711–745. [Google Scholar]
- Rinaudi, L.V.; Giordano, W. An integrated view of biofilm formation in rhizobia. FEMS Microbiol. Lett 2010, 304, 1–11. [Google Scholar]
- Stanley, N.R.; Lazazzera, B.A. Environmental signals and regulatory pathways that influence biofilm formation. Mol. Microbiol 2004, 52, 917–924. [Google Scholar]
- Sutherland, I.W. Biofilm exopolysaccharides: A strong and sticky framework. Microbiology 2001, 147, 3–9. [Google Scholar]
- Flemming, H.C. Biofilms and environmental protection. Water Sci. Technol 1993, 27, 1–10. [Google Scholar]
- Gilbert, P.; Das, J.; Foley, I. Biofilms susceptibility to antimicrobials. Adv. Dent. Res 1997, 11, 160–167. [Google Scholar]
- Matthysse, A.G.; Marry, M.; Krall, L.; Kaye, M.; Ramey, B.E.; Fuqua, C.; White, A.R. The effect of cellulose overproduction on binding and biofilm formation on roots by Agrobacterium tumefaciens. Mol. Plant Microbe Interact 2005, 18, 1002–1010. [Google Scholar]
- Van de Mortel, M.; Halverson, L.J. Cell envelope components contributing to biofilm growth and survival of Pseudomonas putida in low-water-content habitats. Mol. Microbiol 2004, 52, 735–750. [Google Scholar]
- Rudrappa, T.; Biedrzycki, M.L.; Bais, H.P. Causes and consequences of plant-associated biofilms. FEMS Microbiol. Ecol 2008, 64, 153–166. [Google Scholar]
- Mansfield, J.; Genin, S.; Magori, S.; Citovsky, V.; Sriariyanum, M.; Ronald, P.; Dow, M.; Verdier, V.; Beer, S.V.; Machado, M.A.; et al. Top 10 plant pathogenic bacteria in molecular plant pathology. Mol. Plant Pathol 2012, 13, 614–629. [Google Scholar]
- Sorroche, F.G.; Rinaudi, L.V.; Zorreguieta, A.; Giordano, W. EPS II-dependent autoaggregation of Sinorhizobium meliloti planktonic cells. Curr. Microbiol 2010, 61, 465–470. [Google Scholar]
- Sorroche, F.G.; Spesia, M.B.; Zorreguieta, A.; Giordano, W. A positive correlation between bacterial autoaggregation and biofilm formation in native Sinorhizobium meliloti isolates from Argentina. Appl. Environ. Microbiol 2012, 78, 4092–4101. [Google Scholar]
- Neyra, C.; Sadasivan, L. Flocculation in Azospirillum brasilense and Azospirillum lipoferum: Exopolysacharides and cyst formation. J. Bacteriol 1985, 163, 716–723. [Google Scholar]
- Nikitina, V.E.; Ponomareva, E.G.; Alen’kina, S.A.; Konnova, S.A. The role of cell-surface lectins in the aggregation of Azospirilla. Microbiology 2001, 70, 471–476. [Google Scholar]
- Bahat-Samet, E.; Castro-Sowinski, S.; Okon, Y. Arabinose content of extracellular polysaccharide plays a role in cell aggregation of Azospirillum brasilense. FEMS Microbiol. Lett 2004, 237, 195–203. [Google Scholar]
- Klebensberger, J.; Birkenmaier, A.; Geffers, R.; Kjelleberg, S.; Philipp, B. SiaA and SiaD are essential for inducing autoaggregation as a specific response to detergent stress in Pseudomonas aeruginosa. Environ. Microbiol 2009, 11, 3073–3086. [Google Scholar]
- Malik, A.; Sakamoto, M.; Hanazaki, S.; Osawa, M.; Susuki, T.; Tochigi, M.; Kakii, K. Coaggregation among nonfloculating bacteria isolated from activated sludge. Appl. Environ. Microbiol 2003, 69, 6056–6064. [Google Scholar]
- Burdman, S.; Jurkevitch, E.; Soria-Díaz, M.E.; Serrano, A.M.; Okon, Y. Extracellular polysaccharide composition of Azospirillum brasilense and its relation with cell aggregation. FEMS. Microbiol. Lett 2000, 189, 259–264. [Google Scholar]
- Lerouge, I.; Vanderlayden, J. O-antigen structural variation: Mechanisms and possible roles in animal/plant-microbe interactions. FEMS Microbiol. Rev 2001, 26, 17–47. [Google Scholar]
- Priefer, U.B. Genes involved in lipopolysaccharide production and symbiosis are clustered on the chromosome of Rhizobium leguminosarum biovar viciae VF39. J. Bacteriol 1989, 171, 6161–6168. [Google Scholar]
- Cava, J.R.; Elias, P.M.; Turowski, D.A.; Noel, K.D. Rhizobium leguminosarum CFN42 genetic region encoding lipopolysaccharide structures essential for complete nodule development on bean plants. J. Bacteriol 1989, 171, 8–15. [Google Scholar]
- García de los Santos, A.; Brom, S. Characterization of two plasmid-borne lps beta loci of Rhizobium etli required for lipopolysaccharide synthesis and for optimal interaction with plants. Mol. Plant Microbe Interact 1997, 10, 891–902. [Google Scholar]
- Clifford, J.C.; Rapicavoli, J.N.; Roper, M.C. A rhamnose-rich O-antigen mediates adhesion, virulence and host colonization for the xylem-limited phytopathogen, Xylella fastidiosa. Mol. Plant. Microbe Interact. 2013, 26, 676–685. [Google Scholar]
- Hopkins, D.L. Seasonal concentration of the Pierce’s disease bacterium in grapevine stems, petioles, and leaf veins. Phytopathology 1981, 71, 415–418. [Google Scholar]
- Tyson, G.E.; Stojanovic, B.J.; Kuklinski, R.F.; Divittorio, T.J.; Sullivan, M.L. Scanning electron microscopy of Pierce’s disease bacterium in petiolar xylem of grape leaves. Phytopathology 1985, 75, 264–269. [Google Scholar]
- Newman, K.L.; Almeida, R.P.P.; Purcell, A.H.; Lindow, S.E. Use of a green fluorescent strain for analysis of Xylella fastidiosa colonization of Vitis vinifera. Appl. Environ. Microbiol 2003, 69, 7319–7327. [Google Scholar]
- Burdman, S.; Jurkevitch, E.; Schwartsburd, B.; Okon, Y. Involvement of outer membrane proteins in the aggregation of Azospirillum brasilense. Microbiology 1999, 145, 1145–1152. [Google Scholar]
- Mora, P.; Rosconi, F.; Franco Fraguas, L.; Castro-Sowinski, S. Azospirillum brasilense Sp7 produces an outer-membrane lectin that specifically binds to surface-exposed extracellular polysaccharide produced by the bacterium. Arch. Microbiol 2008, 189, 519–524. [Google Scholar]
- Craig, L.; Pique, M.E.; Tainer, J.A. Type IV pilus structure and bacterial pathogenicity. Nature 2004, 2, 363–378. [Google Scholar]
- De La Fuente, L.; Burr, T.J.; Hoch, H.C. Autoaggregation of Xilella fastidiosa cells is influenced by type I and type IV pili. Appl. Environ. Microbiol 2008, 74, 5579–5581. [Google Scholar]
- González, J.E.; Reuhs, B.L.; Walker, G.C. Low molecular weight EPS II of Rhizobium meliloti allows nodule invasion in Medicago sativa. Proc. Natl. Acad. Sci. USA 1996, 93, 8636–8641. [Google Scholar]
- Ausmees, N.; Jonsson, H.; Höglund, S.; Ljunggren, H.; Lindberg, M. Structural and putative regulatory genes involved in cellulose synthesis in Rhizobium leguminosarum bv trifolii. Microbiology 1999, 145, 1253–1262. [Google Scholar]
- Laus, M.C.; van Brussel, A.A.; Kijne, J.W. Role of cellulose fibrils and exopolysaccharides of Rhizobium leguminosarum in attachment to and infection of Vicia sativa root hairs. Mol. Plant Microbe Interact 2005, 18, 533–538. [Google Scholar]
- Williams, A.; Wilkinson, A.; Krehenbrink, M.; Russo, D.M.; Zorreguieta, A.; Downie, J.A. Glucomannan-mediated attachment of Rhizobium leguminosarum to pea root hairs is required for competitive nodule infection. J. Bacteriol 2008, 190, 4706–4715. [Google Scholar]
- Klebensberger, J.; Rui, O.; Fritz, E.; Schink, B.; Philipp, B. Cell aggregation of Pseudomonas aeruginosa strain PAO1 as an energy-dependent stress response during growth with sodium dodecyl sulfate. Arch. Microbiol 2006, 185, 417–427. [Google Scholar]
- Klebensberger, J.; Lautenschlager, K.; Bressler, D.; Wingender, J.; Philipp, B. Detergent-induced cell aggregation in subpopulations of Pseudomonas aeruginosa as a preadaptive survival strategy. Environ. Microbiol 2007, 9, 2247–2259. [Google Scholar]
- Hooshangi, S.; Bentley, W.E. From unicellular properties to multicellular behavior: Bacteria quorum sensing circuitry and applications. Curr. Opin. Biotech 2008, 19, 550–555. [Google Scholar]
- Klausen, M.; Gjermansen, M.; Kreft, J.U.; Tolker-Nielsen, T. Dynamics of development and dispersal in sessile microbial communities: Examples from Pseudomonas aeruginosa and Pseudomonas putida model biofilms. FEMS Microbiol. Lett 2006, 261, 1–11. [Google Scholar]
- Flemming, H.C.; Wingender, J. The biofilm matrix. Nature Rev. Microbiol 2010, 8, 623–633. [Google Scholar]
- Spiers, A.J.; Rainey, P.B. The Pseudomonas fluorescens SBW25 wrinkly spreader biofilm requires attachment factor, cellulose fibre and LPS interactions to maintain strength and integrity. Microbiology 2005, 151, 2829–2839. [Google Scholar]
- Lee, Y.W.; Jeong, S.Y.; In, Y.H.; Kim, K.Y.; So, J.S.; Chang, W.S. Lack of O-polysaccharide enhances biofilm formation by Bradyrhizobium japonicum. Lett. Appl. Microbiol 2010, 50, 452–456. [Google Scholar]
- Vanderlinde, E.M.; Muszynski, A.; Harrison, J.J.; Koval, S.F.; Foreman, D.L.; Ceri, H.; Kannenberg, E.L.; Carlson, R.W.; Yost, C.K. Rhizobium leguminosarum biovar viciae 3841, deficient in 27-hydroxyoctacosanoate-modified lipopolysaccharide, is impaired in desiccation tolerance, biofilm formation and motility. Microbiology 2009, 155, 3055–3069. [Google Scholar]
- Ferguson, G.P.; Roop, R.M., II; Walker, G.C. Deficiency of a Sinorhizobium meliloti bacA mutant in alfalfa symbiosis correlates with alteration of the cell envelope. J. Bacteriol. 2002, 184, 5625–5632. [Google Scholar]
- Hirsch, A.M.; Lum, M.R.; Fujishige, N.A. Microbial Encounters of a Symbiotic Kind-Attaching to Roots and Other Surfaces. In Root Hairs: Plant Cell Monographs; Emons, A.M.C., Ketelaar, T., Eds.; Springer-Verlag: Berlin, Germany, 2009; Volume 12, pp. 295–314. [Google Scholar]
- Lagares, A.; Caetano-Anolles, G.; Niehaus, K.; Lorenzen, J.; Ljunggren, H.D.; Pühler, A.; Favelukes, G. A Rhizobium meliloti lipopolysaccharide mutant altered in competitiveness for nodulation of alfalfa. J. Bacteriol 1992, 174, 5941–5952. [Google Scholar]
- Campbell, G.R.O.; Reuhs, B.L.; Walker, G.C. Chronic intracelular infection of alfalfa nodules by Sinorhizobium meliloti requires correct lipopolysaccharide core. Proc. Natl. Acad. Sci. USA 2002, 99, 3938–3943. [Google Scholar]
- Sorroche, F.G. National University of Río Cuarto: Córdoba, Argentina, Unpublished work; 2011.
- Lau, P.C.Y.; Lindhout, T.; Beveridge, T.J.; Dutcher, J.R.; Lam, J.S. Differential lipopolysaccharide core capping leads to quantitative and correlated modifications of mechanical and structural properties in Pseudomonas aeruginosa biofilms. J. Bacteriol 2009, 191, 6618–6631. [Google Scholar]
- Deng, W.L.; Lin, Y.C.; Lin, R.H.; Wei, C.F.; Huang, Y.C.; Peng, H.L.; Huang, H.C. Effects of galU mutation on Pseudomonas syringae—Plant interactions. Mol. Plant Microbe Interact 2010, 23, 1184–1196. [Google Scholar]
- Li, J.; Wang, N. The wxacO gene of Xanthomonas citri ssp. citri encodes a protein with a role in lipopolysaccharide biosynthesis, biofilm formation, stress tolerance and virulence. Mol. Plant Pathol 2011, 12, 381–396. [Google Scholar]
- Li, J.; Wang, N. Genome-wide mutagenesis of Xanthomonas axonopodis pv. citri reveals novel genetic determinants and regulation mechanisms of biofilm formation. PLoS One 2011, 6, e21804. [Google Scholar]
- Dhakal, B.K.; Bower, J.M.; Mulvey, M.A.; Pili, Fimbriae. Encyclopedia of Microbiology; Moselio, S., Ed.; Academy Press: Oxford, UK, 2009; p. 470. [Google Scholar]
- Conrad, J.C. Physics of bacterial near-surface motility using flagella and type IV pili: Implications for biofilm formation. Res. Microbiol 2012, 163, 619–629. [Google Scholar]
- Petrova, O.E.; Sauer, K. Sticky situations: Key components that control bacterial surface attachment. J. Bacteriol 2012, 194, 2413–2425. [Google Scholar]
- Jin, F.; Conrad, J.C.; Gibiansky, M.L.; Wong, G.C.L. Bacteria use type-IV pili to slingshot on surfaces. Proc. Natl. Acad. Sci. USA 2011, 108, 12617–12622. [Google Scholar]
- Wood, T.K.; González Barrios, A.F.; Herzberg, M.; Lee, J. Motility influences biofilm architecture in Escherichia coli. Appl. Microbiol. Biotechnol 2006, 72, 361–367. [Google Scholar]
- Barken, K.B.; Pamp, S.J.; Yang, L.; Gjermansen, M.; Bertrand, J.J.; Klausen, M.; Givskov, M.; Whitchurch, C.B.; Engel, J.B.; Tolker-Nielsen, T. Roles of type IV pili, flagellum-mediated motility and extracellular DNA in the formation of mature multicellular structures in Pseudomonas aeruginosa biofilms. Environ. Microbiol 2008, 10, 2331–2343. [Google Scholar]
- Bahar, O.; de la Fuente, L.; Burdman, S. Assessing adhesion, biofilm formation and motility of Acidovorax citrulli using microfluidic flow chambers. FEMS Microbiol. Lett 2010, 312, 33–39. [Google Scholar]
- De la Fuente, L.; Montanes, E.; Meng, Y.; Li, Y.; Burr, T.J.; Hoch, H.C.; Wu, M. Assessing adhesion forces of type I and type IV pili of Xylella fastidiosa bacteria by use of a microfluidic flow chamber. Appl. Environ. Microbiol 2007, 73, 2690–2696. [Google Scholar]
- Kang, Y.; Liu, H.; Genin, S.; Schell, M.A.; Denny, T.P. Ralstonia solanacearum requires type 4 pili to adhere to multiple surfaces and for natural transformation and virulence. Mol. Microbiol 2002, 2, 427–437. [Google Scholar]
- Wang, L.; Makino, S.; Subedee, A.; Bogdanove, A.J. Novel candidate virulence factors in rice pathogen Xanthomonas oryzae pv. oryzicola as revealed by mutational analysis. Appl. Environ. Microbiol 2007, 73, 8023–8027. [Google Scholar]
- Jarrell, K.F.; McBride, M.J. The surprisingly diverse ways that prokaryotes move. Nat. Rev. Microbiol 2008, 6, 466–476. [Google Scholar]
- Klausen, M.; Heydorn, A.; Ragas, P.; Lambertsen, L.; Aaes-Jørgensen, A.; Molin, S.; Tolker-Nielsen, T. Biofilm formation by Pseudomonas aeruginosa wild type, flagella and type IV pili mutants. Mol. Microbiol 2003, 48, 1511–1524. [Google Scholar]
- Verstraeten, N.; Braeken, K.; Debkumari, B.; Fauvart, M.; Fransaer, J.; Vermant, J.; Michiels, J. Living on a surface: Swarming and biofilm formation. Trends Microbiol 2008, 16, 496–506. [Google Scholar]
- Fujishige, N.A.; Kapadia, N.N.; de Hoff, P.L.; Hirsch, A.M. Investigations of Rhizobium biofilm formation. FEMS Microbiol. Ecol 2006, 56, 195–206. [Google Scholar]
- Nievas, F.; Bogino, P.; Sorroche, F.; Giordano, W. Detection, characterization, and biological effect of quorum-sensing signaling molecules in peanut-nodulating bradyrhizobia. Sensors 2012, 12, 2851–2873. [Google Scholar]
- Merritt, P.M.; Danhorn, T.; Fuqua, C. Motility and chemotaxis in Agrobacterium tumefaciens surface attachment and biofilm formation. J. Bacteriol 2007, 189, 8005–8014. [Google Scholar]
- Kirov, S.M.; Castrisios, M.; Shaw, J.G. Aeromonas flagella (polar and lateral) are enterocyte adhesins that contribute to biofilm formation on surfaces. Infect. Immun 2004, 72, 1939–1945. [Google Scholar]
- Lemon, K.P.; Higgins, D.E.; Kolter, R. Flagellar motility is critical for Listeria monocytogenes biofilm formation. J. Bacteriol 2007, 189, 4418–4424. [Google Scholar]
- Brunings, A.M.; Gabriel, D.W. Xanthomonas citri: Breaking the surface. Mol. Plant Pathol 2003, 4, 141–157. [Google Scholar]
- Rigano, L.A.; Siciliano, F.; Enrique, R.; Sendín, L.; Filippone, P.; Torres, P.S.; Qüesta, J.; Dow, J.M.; Castagnaro, A.P.; Vojnov, A.A.; et al. Biofilm formation, epiphytic fitness, and canker development in Xanthomonas axonopodis pv citri. Mol. Plant Microbe Interact 2007, 20, 1222–1230. [Google Scholar]
- Malamud, F.; Torres, P.S.; Roeschlin, R.; Rigano, L.A.; Enrique, R.; Bonomi, H.R.; Castagnaro, A.P.; Marano, M.R.; Vojnov, A.A. The Xanthomonas axonopodis pv. citri flagellum is required for mature biofilm and canker development. Microbiology 2011, 157, 819–829. [Google Scholar]
- Lee, M.C.; Weng, S.F.; Tseng, Y.H. Flagellin gene fliC of Xanthomonas campestris is upregulated by transcription factor Clp. Biochem. Biophys. Res. Commun 2003, 307, 647–652. [Google Scholar]
- Tans-Kersten, J.; Huang, H.; Allen, C. Ralstonia solanacearum needs motility for invasive virulence on tomato. J. Bacteriol 2001, 183, 3597–3605. [Google Scholar]
- Herrera, C.M.; Koutsoudis, M.D.; Wang, X.; von Bodman, S.B. Pantoea stewartii subsp. stewartii exhibits surface motility, which is a critical aspect of Stewart’s wilt disease development on maize. Mol. Plant Microbe Interact 2008, 21, 1359–1370. [Google Scholar]
- Jahn, C.E.; Willis, D.K.; Charkowski, A.O. The flagellar sigma factor fliA is required for Dickeya dadantii virulence. Mol. Plant Microbe Interact 2008, 21, 1431–1442. [Google Scholar]
- Cucarella, J.; Solano, C.; Valle, J.; Amorena, B.; Lasa, I.; Penadés, J.R. Bap, a Staphylococcus aureus surface protein involved in biofilm formation. J. Bacteriol 2001, 183, 2888–2896. [Google Scholar]
- Toledo-Arana, A.; Valle, J.; Solano, C.; Arrizubieta, M.J.; Cucarella, J.; Lamata, M. The enterococcal surface protein, Esp, is involved in Enterococcus faecalis biofilm formation. Appl. Environ. Microbiol 2001, 67, 4538–4545. [Google Scholar]
- Ausmees, N.; Jacobsson, K.; Lindberg, M. A unipolarly located, cell-surface-associated agglutinin, RapA, belongs to a family of Rhizobium-adhering proteins (Rap) in Rhizobium leguminosarum bv trifolii. Microbiology 2001, 147, 549–559. [Google Scholar]
- Russo, D.M.; Williams, A.; Edwards, A.; Posadas, D.M.; Finnie, C.; Dankert, M.; Downie, J.A.; Zorreguieta, A. Proteins exported via the PrsD-PrsE type I secretion system and the acidic exopolysaccharide are involved in biofilm formation by Rhizobium leguminosarum. J. Bacteriol 2006, 188, 4474–4486. [Google Scholar]
- Laus, M.C.; Logman, T.J.; Lamers, G.E.; van Brussel, A.A.; Carlson, R.W.; Kijne, J.W. A novel polar surface polysaccharide from Rhizobium leguminosarum binds host plant lectin. Mol. Microbiol 2006, 59, 1704–1713. [Google Scholar]
- Hinsa, S.M.; Espinosa-Urgel, M.; Ramos, J.L.; O’Toole, G.A. Transition from reversible to irreversible attachment during biofilm formation by Pseudomonas fluorescens WCS365 requires an ABC transporter and a large secreted protein. Mol. Microbiol 2003, 49, 905–918. [Google Scholar]
- Martínez-Gil, M.; Yousef-Coronado, F.; Espinosa-Urgel, M. LapF, the second largest Pseudomonas putida protein, contributes to plant root colonization and determines biofilm architecture. Mol. Microbiol 2010, 77, 549–561. [Google Scholar]
- Ryder, C.; Byrd, M.; Wozniak, D.J. Role of exopolysaccharides in Pseudomonas aeruginosa biofilm development. Curr. Opin. Microbiol 2007, 10, 644–648. [Google Scholar]
- Vaningelgem, F.; Zamfir, M.; Mozzi, F.; Adriany, T.; Vancanneyt, M.; Swings, J.; de Vuyst, L. Biodiversity of exopolysaccharides produced by Streptococcus thermophilus strains is reflected in their production and their molecular and functional characteristics. Appl. Environ. Microbiol 2004, 70, 900–912. [Google Scholar]
- Jefferson, K.K. Bacterial Polysaccharides. Current Innovations and Future Trends; Ullrich, M., Ed.; Caister Academic: Norfolk, UK, 2009; pp. 175–186. [Google Scholar]
- Sutherland, I.W. Biofilm Exopolysaccharides. In Microbial Extracellular Polymeric Substances; Wingender, J., Neu, T.R., Flemming, H.C., Eds.; Springer: Berlin, Germany, 1999; pp. 73–92. [Google Scholar]
- Reuber, T.L.; Walker, G.C. Biosynthesis of succinoglycan, a symbiotically important exopolysaccharide of Rhizobium meliloti. Cell 1993, 74, 269–280. [Google Scholar]
- Cheng, H.P.; Walker, G.C. Succinoglycan is required for initiation and elongation of infection threads during nodulation of alfalfa by Rhizobium meliloti. J. Bacteriol. 1998, 180, 5183–5191. [Google Scholar]
- Her, G.R.; Glazebroock, J.; Walker, G.C.; Reinhold, V.N. Structural studies of a novel exopolysaccharide produced by a mutant of Rhizobium meliloti strain Rm1021. Carbohyd. Res 1990, 198, 305–312. [Google Scholar]
- Rinaudi, L.V.; González, J.E. The low-molecular-weight fraction of exopolysaccharide II from Sinorhizobium meliloti is a crucial determinant of biofilm formation. J. Bacteriol 2009, 191, 7216–7224. [Google Scholar]
- Robertson, B.K.; Aman, P.; Darvill, A.G.; McNeil, M.; Albersheim, P. Host-symbiont interactions. The structure of acidic extracellular polysaccharides secreted by Rhizobium leguminosarum and Rhizobium trifolii. Plant Physiol 1981, 67, 389–400. [Google Scholar]
- Philip-Hollingsworth, S.; Hollingsworth, R.I.; Dazzo, F.B. Host-range related structural features of the acidic extracellular polysaccharides of Rhizobium trifolii and Rhizobium leguminosarum. J. Biol. Chem 1989, 264, 1461–1466. [Google Scholar]
- Janczarek, M.; Kutkowska, J.; Piersiak, T.; Skorupska, A. Rhizobium leguminosarum bv. trifolii rosR is required for interaction with clover, biofilm formation and adaptation to the environment. BMC Microbiol. 2010, 10. [Google Scholar] [CrossRef]
- Rolfe, B.G.; Carlson, R.W.; Ridge, R.W.; Dazzo, R.W.; Mateos, F.B.; Pankhurst, C.E. Defective infection and nodulation of clovers by exopolysaccharide mutants of Rhizobium leguminosarum bv trifolii. Aust. J. Plant Physiol 1996, 23, 285–303. [Google Scholar]
- Van Workum, W.A.T.; van Slageren, S.; van Brussel, A.A.N.; Kijne, J.W. Role of exopolysaccharides of Rhizobium leguminosarum bv. viciae as host plant specific molecules required for infection thread formation during nodulation of Vicia sativa. Mol. Plant Microbe Interact 1998, 11, 1233–1241. [Google Scholar]
- Minamisawa, K. Comparison of extracellular polysaccharide composition, rhizobiotoxine production, and hydrogenase phenotype among various strains of Bradyrhizobium japonicum. Plant Cell Physiol 1989, 30, 877–884. [Google Scholar]
- Poveda, A.; Santamaria, M.; Bernabe, M.; Prieto, A.; Briux, M.; Corzo, J.; Jimenez-Barbero, J. Studies on the structure and the solution conformation of an acidic extracellular polysaccharide isolated from Bradyrhizobium. Carbohyd. Res 1997, 304, 209–217. [Google Scholar]
- Pérez-Giménez, J.; Mongiardini, E.J.; Althabegoiti, M.J.; Covelli, J.; Quelas, J.I.; López-García, S.L.; Lodeiro, A.R. Soybean lectin enhances biofilm formation by Bradyrhizobium japonicum in the absence of plants. Int. J. Microbiol. 2009, 2009. [Google Scholar] [CrossRef]
- Wang, P.; Zhong, Z.; Zhou, J.; Cai, T.; Zhu, J. Exopolysaccharide biosynthesis is important for Mesorhizobium tianshanense: Plant host interaction. Arch. Microbiol 2008, 189, 525–530. [Google Scholar]
- Cangelosi, G.A.; Hung, L.; Puvanesarajah, V.; Stacey, G.; Ozga, D.A.; Leigh, J.A.; Nester, E.W. Common loci for Agrobacterium tumefaciens and Rhizobium meliloti exopolysaccharide synthesis and their roles in plant interactions. J. Bacteriol 1987, 169, 2086–2091. [Google Scholar]
- Tomlinson, A.D.; Ramey-Hartung, B.; Day, T.W.; Merritt, P.M.; Fuqua, C. Agrobacterium tumefaciens ExoR represses succinoglycan biosynthesis and is required for biofilm formation and motility. Microbiology 2010, 156, 2670–2681. [Google Scholar]
- Da Silva, F.R.; Vettore, A.L.; Kemper, E.L.; Leite, A.; Arruda, P. Fastidian gum: The Xylella fastidiosa exopolysaccharide possibly involved in bacterial pathogenicity. FEMS Microbiol. Lett 2001, 203, 165–171. [Google Scholar]
- Roper, M.C.; Greve, L.C.; Labavitch, J.M.; Kirkpatrick, B.C. Detection and visualization of an exopolysaccharide produced by Xylella fastidiosa in vitro and in planta. Appl. Environ. Microbiol 2007, 73, 7252–7258. [Google Scholar]
- Jansson, P.E.; Kenne, L.; Lindberg, B. Structure of the extracellular polysaccharide from Xanthomonas campestris. Carbohyd. Res 1975, 45, 274–282. [Google Scholar]
- Rigano, L.A.; Payette, C.; Brouillard, G.; Marano, M.R.; Abramowicz, L.; Torres, P.S.; Yun, M.; Castagnaro, A.P.; Oirdi, M.E.; Dufour, V.; et al. Bacterial cyclic beta-(1,2)-glucan acts in systemic suppression of plant immune responses. Plant Cell 2007, 19, 2077–2089. [Google Scholar]
- Torres, P.S.; Malamud, F.; Rigano, L.A.; Russo, D.M.; Marano, M.R.; Castagnaro, A.P.; Zorreguieta, A.; Bouarab, K.; Dow, J.M.; Vojnov, A.A. Controlled synthesis of the DSF cell-cell signal is required for biofilm formation and virulence in Xanthomonas campestris. Environ. Microbiol 2007, 9, 2101–2109. [Google Scholar]
- Nimtz, M.; Mort, A.; Wray, V.; Domke, T.; Zhang, Y.; Coplin, D.L.; Geider, K. Structure of stewartan, the capsular exopolysaccharide from the corn pathogen Erwinia stewartii. Carbohyd. Res 1996, 288, 189–201. [Google Scholar]
- Koutsoudis, M.D.; Tsaltas, D.; Minogue, T.D.; von Bodman, S.B. Quorum-sensing regulation governs bacterial adhesion, biofilm development, and host colonization in Pantoea stewartii subspecies stewartii. Proc. Natl. Acad. Sci. USA 2006, 103, 5983–5988. [Google Scholar]
- Nimtz, M.; Mort, A.; Domke, T.; Wray, V.; Zhang, Y.; Qiu, F.; Coplin, D.; Geider, K. Structure of amylovoran, the capsular exopolysaccharide from the fire blight pathogen Erwinia amylovora. Carbohyd. Res 1996, 287, 59–76. [Google Scholar]
- Koczan, J.M.; McGrath, M.J.; Zhao, Y.; Sundin, G.W. Contribution of Erwinia amylovora exopolysaccharides amylovoran and levan to biofilm formation: Implications in pathogenicity. Phytopathology 2009, 99, 1237–1244. [Google Scholar]
- Geier, G.; Geider, K. Characterization and influence on virulence of the levansucrase gene from the fireblight pathogen Erwinia amylovora. Physiol. Mol. Plant Pathol 1993, 42, 387–404. [Google Scholar]
- Cook, D.; Sequeira, L. Genetic and biochemical characterization of a gene cluster from Pseudomonas solanacearum required for extracellular polysaccharide production and for virulence. J. Bacteriol 1991, 173, 1654–1662. [Google Scholar]
- Chapman, M.R.; Kao, C.C. EpsR modulates production of extracellular polysaccharides in the bacterial wilt pathogen Ralstonia (Pseudomonas) solanacearum. J. Bacteriol 1998, 180, 27–34. [Google Scholar]
- Zogaj, X.; Nimtz, M.; Rohde, M.; Bokranz, W.; Römling, U. The multicellular morphotypes of Salmonella typhimurium and Escherichia coli produce cellulose as the second component of the extracellular matrix. Mol. Microbiol 2001, 39, 1452–1463. [Google Scholar]
- Matthysse, A.G.; White, S.; Lightfoot, R. Genes required for cellulose synthesis in Agrobacterium tumefaciens. J. Bacteriol 1995, 177, 1069–1075. [Google Scholar]
- Robledo, M.; Rivera, L.; Jiménez-Zurdo, J.I.; Rivas, R.; Dazzo, F.; Velázquez, E.; Martínez-Molina, E.; Hirsch, A.M.; Mateos, P.F. Role of Rhizobium endoglucanase CelC2 in cellulose biosynthesis and biofilm formation on plant roots and abiotic surfaces. Microb. Cell Fact 2012, 11, 125. [Google Scholar]
- Beauregard, P.B.; Chai, Y.; Vlamakis, H.; Losick, R.; Kolter, R. Bacillus subtilis biofilm induction by plant polysaccharides. Proc. Natl. Acad. Sci. USA 2013, 110, 1621–1630. [Google Scholar]
- Zhang, X.; Bishop, P. Biodegradability of biofilm extracellular polymeric substances. Chemosphere 2003, 50, 63–69. [Google Scholar]
- Laue, H.; Schenk, A.; Li, H.; Lambertsen, L.; Neu, T.R.; Molin, S.; Ullrich, M.S. Contribution of alginate and levan production to biofilm formation by Pseudomonas syringae. Microbiology 2006, 152, 2909–2918. [Google Scholar]
- Dafna, T.A.; Rosenberg, T.; Navon, N.; Burdman, S. A secreted lipolytic enzyme from Xanthomonas campestris pv. vesicatoria is expressed in planta and contributes to its virulence. Mol. Plant Pathol 2012, 13, 556–567. [Google Scholar]
- Hu, J.; Qian, W.; He, C. The Xanthomonas oryzae pv. oryzae eglXoB endoglucanase gene is required for virulence to rice. FEMS Microbiol. Lett 2007, 269, 273–279. [Google Scholar]
- Lynch, D.J.; Fountain, T.L.; Mazurkiewicz, J.E.; Banas, J.A. Glucan-binding proteins are essential for shaping Streptococcus mutans biofilm architecture. FEMS Microbiol. Lett 2007, 268, 158–165. [Google Scholar]
- Tielker, D.; Hacker, S.; Loris, R.; Strathmann, M.; Wingender, J.; Wilhelm, S.; Rosenau, F.; Jaeger, K.E. Pseudomonas aeruginosa lectin LecB is located in the outer membrane and is involved in biofilm formation. Microbiology 2005, 151, 1313–1323. [Google Scholar]
- Diggle, S.P.; Stacey, R.E.; Dodd, C.; Cámara, M.; Williams, P.; Winzer, K. The galactophilic lectin, LecA, contributes to biofilm development in Pseudomonas aeruginosa. Environ. Microbiol 2010, 8, 1095–1104. [Google Scholar]
- Branda, S.S.; Chu, F.; Kearns, D.B.; Losick, R.; Kolter, R. A major protein component of the Bacillus subtilis biofilm matrix. Mol. Microbiol 2006, 59, 1229–1238. [Google Scholar]
- Toyofuku, M.; Roschitzki, B.; Riedel, K.; Eberl, L. Identification of proteins associated with the Pseudomonas aeruginosa biofilm extracellular matrix. J. Proteome Res 2012, 11, 4906–4915. [Google Scholar]
- Larsen, P.; Nielsen, J.L.; Dueholm, M.S.; Wetzel, R.; Otzen, D.; Nielsen, P.H. Amyloid adhesins are abundant in natural biofilms. Environ. Microbiol 2007, 9, 3077–3090. [Google Scholar]
- Molin, S.; Tolker-Nielsen, T. Gene transfer occurs with enhanced efficiency in biofilms and induces enhanced stabilisation of the biofilm structure. Curr. Opin. Biotechnol 2003, 14, 255–261. [Google Scholar]
- Dominiak, D.M.; Nielsen, J.L.; Nielsen, P.H. Extracellular DNA is abundant and important for microcolony strength in mixed microbial biofilms. Environ. Microbiol 2011, 13, 710–721. [Google Scholar]
- Yang, L.; Barken, K.B.; Skindersoe, M.E.; Christensen, A.B.; Givskov, M.; Tolker-Nielsen, T. Effects of iron on DNA release and biofilm development by Pseudomonas aeruginosa. Microbiology 2007, 153, 1318–1328. [Google Scholar]
- Vilain, S.; Pretorius, J.M.; Theron, J.; Broezel, V.S. DNA as an adhesion: Bacillus cereus requires extracellular DNA to form biofilms. Appl. Environ. Microbiol 2009, 75, 2861–2868. [Google Scholar]
- Böckelmann, U.; Janke, A.; Kuhn, R.; Neu, T.R.; Wecke, J.; Lawrence, J.R.; Szewzyk, U. Bacterial extracellular DNA forming a defined network-like structure. FEMS Microbiol. Lett 2006, 262, 31–38. [Google Scholar]
- Allesen-Holm, M.; Barken, K.B.; Yang, L.; Klausen, M.; Webb, J.S.; Kjelleberg, S.; Molin, S.; Givskov, M.; Tolker-Nielsen, T. A characterization of DNA release in Pseudomonas aeruginosa cultures and biofilms. Mol. Microbiol 2006, 59, 1114–1128. [Google Scholar]
- Wang, J.; Bayles, K.W. Programmed cell death in plants: Lessons from bacteria? Trends Plant Sci 2013, 18, 133–139. [Google Scholar]
- Das, T.; Prashant, S.K.; Busscher, H.J.; van der Mei, H.C.; Krom, B.P. Role of extracellular DNA in initial bacterial adhesion and surface aggregation. Appl. Environ. Microbiol 2010, 76, 3405–3408. [Google Scholar]
- Conrad, A.; Suutari, M.K.; Keinänen, M.M.; Cadoret, A.; Faure, P.; Mansuy-Huault, L.; Block, J.C. Fatty acid lipid fractions in extracellular polymeric substances of activated sludge flocs. Lipids 2003, 38, 1093–1105. [Google Scholar]
- Ron, E.Z.; Rosenberg, E. Natural role of biosurfactants. Environ. Microbiol 2001, 3, 229–236. [Google Scholar]
- Pamp, S.J.; Tolker-Nielsen, T. Multiple roles of biosurfactants in structural biofilm development by Pseudomonas aeruginosa. J. Bacteriol 2007, 189, 2531–2539. [Google Scholar]
- Stoodley, P.; Sauer, K.; Davies, D.G.; Costerton, J.W. Biofilms as complex differentiated communities. Annu. Rev. Microbiol 2002, 56, 187–209. [Google Scholar]
- Sherlock, O.; Schembri, M.A.; Reisner, A.; Klemm, P. Novel roles for the AIDA adhesin from diarrheagenic Escherichia coli: Cell aggregation and biofilm formation. J. Bacteriol 2004, 186, 8058–8065. [Google Scholar]
- Jacobs, A.; Chenia, H.Y. Biofilm-forming capacity, surface hydrophobicity and aggregation characteristics of Myroides odoratus isolated from South African Oreochromis mossambicus fish. J. Appl. Microbiol 2009, 107, 1957–1966. [Google Scholar]
- Yamaguchi, M.; Sato, K.; Yukitake, H.; Noiri, Y.; Ebisu, S.; Nakayama, K. Porphyromonas gingivalis mutant defective in a putative glycosyltransferase exhibits defect in biosynthesis of polysaccharide portions of lipopolysaccharide, decreased gingipain activities, strong autoaggregation and increased biofilm formation. Infec. Immun 2010, 78, 3801–3812. [Google Scholar]
- Voegel, T.M.; Doddapaneni, H.; Cheng, D.W.; Lin, H.; Stenger, D.C.; Kirkpatrick, B.C.; Roper, M.C. Identification of a response regulator involved in surface attachment, cell–cell aggregation, exopolysaccharide production and virulence in the plant pathogen Xylella fastidiosa. Mol. Plant Pathol 2013, 14, 256–264. [Google Scholar]
- Nievas, F. National University of Río Cuarto: Córdoba, Argentina, Unpublished work; 2012.
- Rickard, A.H.; Gilbert, P.; High, N.J.; Kolenbrander, P.E.; Handley, P.S. Bacterial Coaggregation: An integral process in the development of multi-species biofilms. Trends Microbiol 2003, 11, 94–100. [Google Scholar]
- Phuong, K.; Kakii, K.; Nikata, T. Intergeneric coaggregation of non-flocculating Acinetobacter spp. isolates with other sludge-constituting bacteria. J. Biosci. Bioeng 2009, 107, 394–400. [Google Scholar]
- Joe, M.M.; Jaleel, C.A.; Sivakumar, P.K.; Zhao, C.X.; Karthikeyan, B. Co-aggregation in Azospirillum brasilensense MTCC-125 with other PGPR strains: Effect of physical and chemical factors and stress endurance ability. J. Taiwan Inst. Chem. E 2009, 40, 491–499. [Google Scholar]
- Malik, A.; Kakii, K. Pair dependent co-aggregation behavior of nonflocculating sludge bacteria. Biotechnol Lett 2003, 25, 981–986. [Google Scholar]
- Ledder, R.G.; Timperley, A.S.; Friswell, M.K.; MacFarlane, S.; McBain, A.J. Coaggregation between and among human intestinal and oral bacteria. FEMS Microbiol. Ecol 2008, 66, 630–636. [Google Scholar]


{kind=link}
| Bacteria/plant association | Exopolysaccharide | Chemical composition | Function |
|---|---|---|---|
| E. meliloti symbiosis with Medicago sativa | Succinoglycan (EPS I), LMW-HMW [91] | Octasaccharide units (glucose:galactose 7:1, bearing succinyl, acetyl, and pyruvyl substituents) | Required for biofilm formation [68] EPS I LMW symbiotically active [92] |
| Galactoglucan (EPS II), LMW-HMW [93] | Disaccharide units (acetylated glucose-pyruvylated galactose) | EPS II LMW symbiotically active [34] EPS II LMW controls biofilm formation [94] | |
| R. leguminosarum symbiosis with Trifolium, Pisum, Vicia and Phaseolus spp. | Acidic EPS [95,96] | Octasaccharide units (glucose:glucuronic acid:galactose 5:2:1, modified by acetyl, pyruvyl and 3-hydroxybutanoyl groups) | Development of a structured biofilm [83,97] Required for infection and nodulation [98,99] |
| B. japonicum symbiosis with Glycine max | EPS [100,101] | Pentasaccharide units (mannose:galacturonic acid:glucose:galactose 1:1:2:1) | Biofilm formation on both inert and biotic surfaces. Roles during the early stages of interaction with the host plant (initial attachment of rhizobia to root epidermal cells) [102] |
| M. tianshanense symbiosis with Glycyrrhiza uralensis | EPS | ND | Involved in biofilm formation and successful establishment of symbiosis [103] |
| A. tumefaciens ubiquitous plant pathogen | Succinoglycan [104] | See above | Increased production of succinoglycan results in reduced attachment and biofilm formation [105] |
| X. fastidiosa plant pathogen | Putative Fastidian gum [106] | Putative tetrasaccharide units (glucose-1-phosphate, glucose, mannose, and glucuronic acid) | Possibly involved in bacterial pathogenicity [106] Cell attachment and overall biofilm formation [107] |
| X. campestris X. axonopodis plant pathogens | Xanthan gum [108] | Pentasaccharide units (glucose:mannose:glucuronic acid 2:2:1 derivatized with acetyl and pyruvyl moieties) | Essential for microcolony formation [74] Formation of structured biofilms on abiotic surfaces and in infected plants [109,110] |
| P. stewartii plant pathogen | Stewartan [111] | Heptasaccharide units (glucose:galactose:glucuronic acid 3:3:1) | Essential for appropriate adhesion and for maturation of biofilm structure. Also a virulence factor required for effective host colonization and efficient dissemination through xylem vessels [112] |
| E. amylovora plant pathogen | Amylovoran [113] | Pentasaccharide units (galactose:glucose 4:1, and pyruvate residues) | Pathogenicity factor required for biofilm formation [114] |
| Levan [115] | Homopolymer of fructose | Virulence factor. Also contributes to biofilm formation [114] | |
| R. solanacearum plant pathogen | Acidic EPS I [116] | Putative structure composed by N-acetylgalactosamine and amino sugars (bacillosamine, galactosaminuronic acid) | Major virulence factor [117] |
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Bogino, P.C.; Oliva, M.D.l.M.; Sorroche, F.G.; Giordano, W. The Role of Bacterial Biofilms and Surface Components in Plant-Bacterial Associations. Int. J. Mol. Sci. 2013, 14, 15838-15859. https://doi.org/10.3390/ijms140815838
Bogino PC, Oliva MDlM, Sorroche FG, Giordano W. The Role of Bacterial Biofilms and Surface Components in Plant-Bacterial Associations. International Journal of Molecular Sciences. 2013; 14(8):15838-15859. https://doi.org/10.3390/ijms140815838
Chicago/Turabian StyleBogino, Pablo C., María De las Mercedes Oliva, Fernando G. Sorroche, and Walter Giordano. 2013. "The Role of Bacterial Biofilms and Surface Components in Plant-Bacterial Associations" International Journal of Molecular Sciences 14, no. 8: 15838-15859. https://doi.org/10.3390/ijms140815838
APA StyleBogino, P. C., Oliva, M. D. l. M., Sorroche, F. G., & Giordano, W. (2013). The Role of Bacterial Biofilms and Surface Components in Plant-Bacterial Associations. International Journal of Molecular Sciences, 14(8), 15838-15859. https://doi.org/10.3390/ijms140815838