Renal Dopamine Receptors, Oxidative Stress, and Hypertension

Abstract
:1. Introduction
1.1. Renal Dopaminergic System
1.2. Reactive Oxygen Species
2. Renal Dopamine Receptor Regulation of Oxidative Stress
2.1. D1-Like Receptors
2.1.1. Renal Expression of D1-Like Dopamine Receptors
2.1.2. D1-Like Receptors Negatively Regulate ROS Production
2.1.3. ROS Negatively Regulate D1-Like Receptor Expression and Function
2.2. D2-Like Receptors
2.2.1. Renal Expression of D2-Like Dopamine Receptors
2.2.2. D2R Negatively Regulates ROS Production
2.2.2.1. D2R Protects against Oxidative Stress: Role of NADPH Oxidase
2.2.2.2. D2R Protects against Oxidative Stress: Role of the Antioxidant, DJ-1
2.2.2.3. D2R Protects against Oxidative Stress: Role of the Antioxidant, PON2
2.2.2.4. D2R Protects against Oxidative Stress and Inflammation
2.3. D3R and Oxidative Stress
2.4. D4R and Oxidative Stress
3. Summary
Acknowledgments
Conflicts of Interest
References
- Prosser, B.L.; Ward, C.W.; Lederer, W.J. X-ROS signaling: Rapid mechano-chemo transduction in heart. Science 2011, 333, 1440–1445. [Google Scholar]
- Rhee, S.G. Cell signaling. H2O2, a necessary evil for cell signaling. Science 2006, 312, 1882–1883. [Google Scholar]
- Ristow, M.; Zarse, K. How increased oxidative stress promotes longevity and metabolic health: The concept of mitochondrial hormesis (mitohormesis). Exp. Gerontol 2010, 45, 410–418. [Google Scholar]
- Bretón-Romero, R.; Lamas, S. Hydrogen Peroxide signaling mediator in the activation of p38 MAPK in vascular endothelial cells. Methods Enzymol 2013, 528, 49–59. [Google Scholar]
- Chung, H.S.; Wang, S.B.; Venkatraman, V.; Murray, C.I.; van Eyk, J.E. Cysteine oxidative posttranslational modifications: Emerging regulation in the cardiovascular system. Circ. Res 2013, 112, 382–392. [Google Scholar]
- Evans, J.L.; Goldfine, I.D.; Maddux, B.A.; Grodsky, G.M. Are oxidative stress-activated signaling pathways mediators of insulin resistance and β-cell dysfunction? Diabetes 2003, 52, 1–8. [Google Scholar]
- Lassègue, B.; San Martín, A.; Griendling, K.K. Biochemistry, physiology, and pathophysiology of NADPH oxidases in the cardiovascular system. Circ. Res 2012, 110, 1364–1390. [Google Scholar]
- Sedeek, M.; Hébert, R.L.; Kennedy, C.R.; Burns, K.D.; Touyz, R.M. Molecular mechanisms of hypertension: Role of Nox family NADPH oxidases. Curr. Opin. Nephrol. Hypertens 2009, 18, 122–127. [Google Scholar]
- Chen, F.; Haigh, S.; Barman, S.; Fulton, D.J. From form to function: The role of Nox4 in the cardiovascular system. Front. Physiol 2012, 3, 1–12. [Google Scholar]
- Lo, J.; Patel, V.B.; Wang, Z.; Levasseur, J.; Kaufman, S.; Penninger, J.M.; Oudit, G.Y. Angiotensin-converting enzyme 2 antagonizes angiotensin II-induced pressor response and NADPH oxidase activation in Wistar-Kyoto rats and spontaneously hypertensive rats. Exp. Physiol. 2013, 98, 109–122. [Google Scholar]
- Wilcox, C.S. Asymmetric dimethylarginine and reactive oxygen species: Unwelcome twin visitors to the cardiovascular and kidney disease tables. Hypertension 2012, 59, 375–381. [Google Scholar]
- Wingler, K.; Wunsch, S.; Kreutz, R.; Rothermund, L.; Paul, M.; Schmidt, H.H. Upregulation of the vascular NAD(P)H oxidase isoforms Nox1 and Nox4 by the renin-angiotensin system in vitro and in vivo. Free Radic. Biol. Med 2001, 31, 1456–1464. [Google Scholar]
- Wind, S.; Beuerlein, K.; Armitagr, E.; Taye, A.; Kumar, H.; Janowitz, D.; Neff, C.; Shar, M.; Wingler, K.; Schmidt, H.H. Oxidative stress and endothelial dysfunction in aortas of aged spontaneously hypertensive rats by NOX1/2 is reversed by NADPH oxidase inhibition. Hypertension 2010, 56, 490–497. [Google Scholar]
- Yu, P.Y.; Han, W.X.; Zhang, Y.R.; Villar, V.A.M.; Li, H.W.; Felder, R.A.; Jose, P.A. NADPH oxidase 5 may play a role in human essential hypertension. Free Radic. Biol. Med 2011, 51, S53. [Google Scholar]
- Manea, A. NADPH oxidase-derived reactive oxygen species: Involvement in vascular physiology and pathology. Cell Tissue Res 2010, 342, 325–339. [Google Scholar]
- Morigi, M.; Macconi, D.; Zoja, C.; Donadelli, R.; Buelli, S.; Zanchi, C.; Ghilardi, M.; Remuzzi, G. Protein overload-induced NF-κB activation in proximal tubular cells requires H2O2 through a PKC-dependent pathway. J. Am. Soc. Nephrol 2002, 13, 1179–1189. [Google Scholar]
- Li, H.; Han, W.; Villar, V.A.; Keever, L.B.; Lu, Q.; Hopfer, U.; Quinn, M.T.; Felder, R.A.; Jose, P.A.; Yu, P. D1-like receptors regulate NADPH oxidase activity and subunit expression in lipid raft microdomains of renal proximal tubule cells. Hypertension 2009, 53, 1054–1061. [Google Scholar]
- Han, W.; Li, H.; Villar, V.A.; Pascua, A.M.; Dajani, M.I.; Wang, X.; Natarajan, A.; Quinn, M.T.; Felder, R.A.; Jose, P.A.; Yu, P. Lipid rafts keep NADPH oxidase in the inactive state in human renal proximal tubule cells. Hypertension 2008, 51, 481–487. [Google Scholar]
- Armando, I.; Villar, V.A.; Jose, P.A. Dopamine and renal function and blood pressure regulation. Compr. Physiol 2011, 1, 1075–1117. [Google Scholar]
- Zhang, M.Z.; Yao, B.; Wang, S.; Fan, X.; Wu, G.; Yang, H.; Yin, H.; Yang, S.; Harris, R.C. Intrarenal dopamine deficiency leads to hypertension and decreased longevity in mice. J. Clin. Invest 2011, 121, 2845–2854. [Google Scholar]
- Zhang, M.Z.; Yao, B.; Yang, S.; Yang, H.; Wang, S.; Fan, X.; Yin, H.; Fogo, A.B.; Moeckel, G.W.; Harris, R.C. Intrarenal dopamine inhibits progression of diabetic nephropathy. Diabetes 2012, 61, 2575–2584. [Google Scholar]
- Souza, B.R.; Romano-Silva, M.A.; Tropepe, V. Dopamine D2 receptor activity modulates Akt signaling and alters GABAergic neuron development and motor behavior in zebrafish larvae. J. Neurosci 2011, 31, 5512–5525. [Google Scholar]
- Pollack, A. Coactivation of D1 and D2 dopamine receptors: in marriage, a case of his, hers, and theirs. Sci. STKE 2004, 255, pe50. [Google Scholar]
- Yu, P.Y.; Asico, L.D.; Eisner, G.M.; Jose, P.A. Differential regulation of renal phospholipase C isoforms by catecholamines. J. Clin. Invest 1995, 95, 304–308. [Google Scholar]
- Yu, P.Y.; Eisner, G.M.; Yamaguchi, I.; Mouradian, M.M.; Felder, R.A.; Jose, P.A. Dopamine D1A receptor regulation of phospholipase C isoform. J. Biol. Chem 1996, 271, 19503–19508. [Google Scholar]
- Yao, B.; Harris, R.C.; Zhang, M.Z. Intrarenal dopamine attenuates deoxycorticosterone acetate/high salt-induced blood pressure elevation in part through activation of a medullary cyclooxygenase 2 pathway. Hypertension 2009, 54, 1077–1083. [Google Scholar]
- Cosentino, M.; Rasini, E.; Colombo, C.; Marino, F.; Blandini, F.; Ferrari, M.; Samuele, A.; Lecchini, S.; Nappi, G.; Frigo, G. Dopaminergic modulation of oxidative stress and apoptosis in human peripheral blood lymphocytes: Evidence for a D1-like receptor-dependent protective effect. Free Radic. Biol. Med 2004, 36, 1233–1240. [Google Scholar]
- Lev, N.; Ickowicz, D.; Barhum, Y.; Lev, S.; Melamed, E.; Offen, D. DJ-1 protects against dopamine toxicity. J. Neural Transm 2009, 116, 151–160. [Google Scholar]
- Luo, Y.; Roth, G.S. The roles of dopamine oxidative stress and dopamine receptor signaling in aging and age-related neurodegeneration. Antioxid. Redox Signal 2000, 2, 449–460. [Google Scholar]
- Eibl, J.K.; Abdallah, Z.; Ross, G.M. Zinc-metallothionein: A potential mediator of antioxidant defence mechanisms in response to dopamine-induced stress. Can. J. Physiol. Pharmacol 2010, 88, 305–312. [Google Scholar]
- Amenta, F.; Barili, P.; Bronzetti, E.; Ricci, A. Dopamine D1-like receptor subtypes in the rat kidney: A microanatomical study. Clin. Exp. Hypertens 1999, 21, 17–23. [Google Scholar]
- O’Connell, D.P.; Botkin, S.J.; Ramos, S.I.; Sibley, D.R.; Ariano, M.A.; Felder, R.A.; Carey, R.M. Localization of dopamine D1A receptor protein in rat kidneys. Am. J. Physiol 1995, 268, F1185–F1197. [Google Scholar]
- Wang, X.; Villar, V.A.; Armando, I.; Eisner, G.M.; Felder, R.A.; Jose, P.A. Dopamine, kidney, and hypertension:Studies in dopamine receptor knockout mice. Pediatr. Nephrol. 2008, 23, 2131–2146. [Google Scholar]
- Bek, M.; Fischer, K.G.; Greiber, S.; Hupfer, C.; Mundel, P.; Pavenstädt, H. Dopamine depolarizes podocytes via a D1-like receptor. Nephrol. Dial. Transplant 1999, 14, 581–587. [Google Scholar]
- Yamaguchi, I.; Yao, L.; Sanada, H.; Ozono, R.; Mouradian, M.M.; Jose, P.A.; Carey, R.M.; Felder, R.A. Dopamine D1A receptors and renin release in rat juxtaglomerular cells. Hypertension 1997, 29, 962–968. [Google Scholar]
- O’Connell, D.P.; Aherne, A.M.; Lane, E.; Felder, R.A.; Carey, R.M. Detection of dopamine receptor D1A subtype-specific mRNA in rat kidney by in situ amplification. Am. J. Physiol 1998, 274, F232–F341. [Google Scholar]
- Ozono, R.; O’Connell, D.P.; Wang, Z.Q.; Moore, A.F.; Sanada, H.; Felder, R.A.; Carey, R.M. Localization of the dopamine D1 receptor protein in the human heart and kidney. Hypertension 1997, 30, 725–729. [Google Scholar]
- Jose, P.A.; Soares-da-Silva, P.; Eisner, G.M.; Felder, R.A. Dopamine and G protein-coupled receptor kinase 4 in the kidney: Role in blood pressure regulation. Biochim. Biophys. Acta 2010, 1802, 1259–1267. [Google Scholar]
- Moussa, C.E.; Tomita, Y.; Sidhu, A. Dopamine D1 receptor-mediated toxicity in human SK-N-MC neuroblastoma cells. Neurochem. Int 2006, 48, 226–234. [Google Scholar]
- Yu, Y.; Wang, J.R.; Sun, P.H.; Guo, Y.; Zhang, Z.J.; Jin, G.Z.; Zhen, X. Neuroprotective effects of atypical D1 receptor agonist SKF83959 are mediated via D1 receptor-dependent inhibition of glycogen synthase kinase-3 beta and a receptor-independent anti-oxidative action. J. Neurochem 2008, 104, 946–956. [Google Scholar]
- Yasunari, K.; Kohno, M.; Hasuma, T.; Horio, T.; Kano, H.; Yokokawa, K.; Minami, M.; Yoshikawa, J. Dopamine as a novel antimigration and antiproliferative factor of vascular smooth muscle cells through dopamine D1-like receptors. Arterioscler. Thromb. Vasc. Biol. 1997, 17, 3164–3173. [Google Scholar]
- Noh, J.S.; Gwag, B.J. Attenuation of oxidative neuronal necrosis by a dopamine D1 agonist in mouse cortical cell cultures. Exp. Neurol 1997, 146, 604–608. [Google Scholar]
- Yang, Z.; Asico, L.D.; Yu, P.; Wang, Z.; Jones, J.E.; Bai, R.K.; Sibley, D.R.; Felder, R.A.; Jose, P.A. D5 dopamine receptor regulation of phospholipase D. Am. J. Physiol. Heart. Circ. Physiol 2005, 288, H55–H61. [Google Scholar]
- Li, G.Y.; Li, T.; Fan, B.; Zheng, Y.C.; Ma, T.H. The D1 dopamine receptor agonist, SKF83959, attenuates hydrogen peroxide-induced injury in RGC-5 cells involving the extracellular signal-regulated kinase/p38 pathways. Mol. Vis 2012, 18, 2882–2895. [Google Scholar]
- Yasunari, K.; Kohno, M.; Kano, H.; Minami, M.; Yoshikawa, J. Dopamine as a novel antioxidative agent for rat vascular smooth muscle cells through dopamine D1-like receptors. Circulation 2000, 101, 2302–2308. [Google Scholar]
- George, L.E.; Lokhandwala, M.F.; Asghar, M. Novel role of NF-κB-p65 in antioxidant homeostasis in human kidney. Am. J. Physiol. Renal Physiol 2012, 302, F1440–F1446. [Google Scholar]
- Yu, P.; Han, W.X.; Sun, M.; Villar, V.A.M.; Jose, P.A. Protein kinase C inhibits NADPH oxidase activity via cross-talk with protein kinase A in HEK-293 heterologously expressing D1 receptor cells. J. Am. Soc. Nephrol 2009, 20, 533A. [Google Scholar]
- Lu, Q.; Yang, Y.; Villar, V.A.; Asico, L.; Jones, J.E.; Yu, P.; Li, H.; Weinman, E.J.; Eisner, G.M.; Jose, P.A. D5 dopamine receptor decreases NADPH oxidase, reactive oxygen species and blood pressure via heme oxygenase-1. Hypertens. Res 2013, 36, 684–690. [Google Scholar]
- Yang, Z.; Asico, L.D.; Yu, P.; Wang, Z.; Jones, J.E.; Escano, C.S.; Wang, X.; Quinn, M.T.; Sibley, D.R.; Romero, G.G.; Felder, R.A.; Jose, P.A. D5 dopamine receptor regulation of reactive oxygen species production, NADPH oxidase, and blood pressure. Am. J. Physiol. Regul. Integr. Comp. Physiol 2006, 290, R96–R104. [Google Scholar]
- Stoelting, M.; Geyer, M.; Reuter, S.; Reichelt, R.; Bek, M.J.; Pavenstädt, H. α/β hydrolase 1 is upregulated in D5 dopamine receptor knockout mice and reduces O2− production of NADPH oxidase. Biochem. Biophys. Res. Commun 2009, 379, 81–85. [Google Scholar]
- Onozato, M.L.; Tojo, A.; Kobayashi, N.; Goto, A.; Matsuoka, H.; Fujita, T. Dual blockade of aldosterone and angiotensin II additively suppresses TGF-β and NADPH oxidase in the hypertensive kidney. Nephrol. Dial. Transplant 2007, 22, 1314–1322. [Google Scholar]
- Fazeli, G.; Stopper, H.; Schinzel, R.; Ni, C.W.; Jo, H.; Schupp, N. Angiotensin II induces DNA damage via AT1 receptor and NADPH oxidase isoform Nox4. Mutagenesis 2012, 27, 673–681. [Google Scholar]
- Khan, F.; Spicarová, Z.; Zelenin, S.; Holtbäck, U.; Scott, L.; Aperia, A. Negative reciprocity between angiotensin II type 1 and dopamine D1 receptors in rat renal proximal tubule cells. Am. J. Physiol. Renal Physiol 2008, 295, F1110–F1116. [Google Scholar]
- Zeng, C.; Luo, Y.; Asico, L.D.; Hopfer, U.; Eisner, G.M.; Felder, R.A.; Jose, P.A. Perturbation of D1 dopamine and AT1 receptor interaction in spontaneously hypertensive rats. Hypertension 2003, 42, 787–792. [Google Scholar]
- Zeng, C.; Wang, Z.; Hopfer, U.; Asico, L.D.; Eisner, G.M.; Felder, R.A.; Jose, P.A. Rat strain effects of AT1 receptor activation on D1 dopamine receptors in immortalized renal proximal tubule cells. Hypertension 2005, 46, 799–805. [Google Scholar]
- Li, H.; Armando, I.; Yu, P.; Escano, C.; Mueller, S.C.; Asico, L.; Pascua, A.; Lu, Q.; Wang, X.; Villar, V.A.; et al. Dopamine 5 receptor mediates Ang II type 1 receptor degradation via a ubiquitin-proteasome pathway in mice and human cells. J. Clin. Invest 2008, 118, 2180–2189. [Google Scholar]
- Zeng, C.; Yang, Z.; Wang, Z.; Jones, J.; Wang, X.; Altea, J.; Mangrum, A.J.; Hopfer, U.; Sibley, D.R.; Eisner, G.M.; et al. Interaction of angiotensin II type 1 and D5 dopamine receptors in renal proximal tubule cells. Hypertension 2005, 45, 804–810. [Google Scholar]
- Wang, Z.; Armando, I.; Asico, L.D.; Escano, C.; Wang, X.; Lu, Q.; Felder, R.A.; Schnackenberg, C.G.; Sibley, D.R.; Eisner, G.M.; et al. The elevated blood pressure of human GRK4gamma A142V transgenic mice is not associated with increased ROS production. Am. J. Physiol. Heart. Circ. Physiol 2007, 292, H2083–H2092. [Google Scholar]
- Asghar, M.; Chillar, A.; Lokhandwala, M.F. Renal proximal tubules from old Fischer 344 rats grow into epithelial cells in cultures and exhibit increased oxidative stress and reduced D1 receptor function. Am. J. Physiol. Cell Physiol 2008, 295, C1326–C1331. [Google Scholar]
- Fardoun, R.Z.; Asghar, M.; Lokhandwala, M. Role of oxidative stress in defective renal dopamine D1 receptor-G protein coupling and function in old Fischer 344 rats. Am. J. Physiol. Renal Physiol 2006, 291, F945–F951. [Google Scholar]
- Asghar, M.; George, L.; Lokhandwala, M.F. Exercise decreases oxidative stress and inflammation and restores renal dopamine D1 receptor function in old rats. Am. J. Physiol. Renal Physiol 2007, 293, F914–F919. [Google Scholar]
- Banday, A.A.; Marwaha, A.; Tallam, L.S.; Lokhandwala, M.F. Tempol reduces oxidative stress, improves insulin sensitivity, decreases renal dopamine D1 receptor hyperphosphorylation, and restores D1 receptor-G-protein coupling and function in obese Zucker rats. Diabetes 2005, 54, 2219–2226. [Google Scholar]
- Marwaha, A.; Lokhandwala, M.F. Tempol reduces oxidative stress and restores renal dopamine D1-like receptor-G protein coupling and function in hyperglycemic rats. Am. J. Physiol. Renal Physiol 2006, 291, F58–F66. [Google Scholar]
- Banday, A.A.; Fazili, F.R.; Lokhandwala, M.F. Oxidative stress causes renal dopamine D1 receptor dysfunction and hypertension via mechanisms that involve nuclear factor-kappaB and protein kinase C. J. Am. Soc. Nephrol 2007, 18, 1446–1457. [Google Scholar]
- George, L.; Lokhandwala, M.F.; Asghar, M. Exercise activates redox-sensitive transcription factors and restores renal D1 receptor function in old rats. Am. J. Physiol. Renal Physiol 2009, 297, F1174–F1180. [Google Scholar]
- Banday, A.A.; Lokhandwala, M.F. Oxidative stress reduces renal dopamine D1 receptor-Gq/11alpha G protein-phospholipase C signaling involving G protein-coupled receptor kinase 2. Am. J. Physiol. Renal Physiol 2007, 293, F306–F315. [Google Scholar]
- Asghar, M.; Banday, A.A.; Fardoun, R.Z.; Lokhandwala, M.F. Hydrogen peroxide causes uncoupling of dopamine D1-like receptors from G proteins via a mechanism involving protein kinase C and G-protein-coupled receptor kinase 2. Free Radic. Biol. Med 2006, 40, 13–20. [Google Scholar]
- Trivedi, M.; Lokhandwala, M.F. Rosiglitazone restores renal D1A receptor-Gs protein coupling by reducing receptor hyperphosphorylation in obese rats. Am. J. Physiol. Renal Physiol 2005, 289, F298–F304. [Google Scholar]
- Shin, Y.; Kumar, U.; Patel, Y.; Patel, S.C.; Sidhu, A. Differential expression of D2-like dopamine receptors in the kidney of the spontaneously hypertensive rat. J. Hypertens 2003, 21, 199–207. [Google Scholar]
- Gao, D.Q.; Canessa, L.M.; Mouradian, M.M.; Jose, P.A. Expression of the D2 subfamily of dopamine receptor genes in kidney. Am. J. Physiol 1994, 266, F646–F650. [Google Scholar]
- Nurnberger, A.; Rabiger, M.; Mack, A.; Diaz, J.; Sokoloff, P.; Muhlbauer, B.; Luippold, G. Subapical localization of the dopamine D3 receptor in proximal tubules of the rat kidney. J. Histochem. Cytochem 2004, 52, 1647–1655. [Google Scholar]
- Ricci, A.; Marchal-Victorion, S.; Bronzetti, E.; Parini, A.; Amenta, F.; Tayebati, S.K. Dopamine D4 receptor expression in rat kidney: Evidence for pre- and postjunctional localization. J. Histochem. Cytochem 2002, 50, 1091–1096. [Google Scholar]
- Sun, D.; Wilborn, T.W.; Schafer, J.A. Dopamine D4 receptor isoform mRNA and protein are expressed in the rat cortical collecting duct. Am. J. Physiol 1998, 275, F742–F751. [Google Scholar]
- Sanada, H.; Yao, L.; Jose, P.A.; Carey, R.M.; Felder, R.A. Dopamine D3 receptors in rat juxtaglomerular cells. Clin. Exp. Hypertens 1997, 19, 93–105. [Google Scholar]
- Matsumoto, M.; Hidaka, K.; Tada, S.; Tasaki, Y.; Yamaguchi, T. Full-length cDNA cloning and distribution of human dopamine D4 receptor. Brain Res. Mol. Brain Res 1995, 29, 157–162. [Google Scholar]
- Wiederkehr, M.R.; di Sole, F.; Collazo, R.; Quiñones, H.; Fan, L.; Murer, H.; Helmle-Kolb, C.; Moe, O.W. Characterization of acute inhibition of Na/H exchanger NHE-3 by dopamine in opossum kidney cells. Kidney Int 2001, 59, 197–209. [Google Scholar]
- Eklof, A.C. The natriuretic response to a dopamine DA1 agonist requires endogenous activation of dopamine DA2 receptors. Acta Physiol. Scand 1997, 160, 311–314. [Google Scholar]
- Kitamura, Y.; Taniguchi, T.; Shimohama, S.; Akaike, A.; Nomura, Y. Neuroprotective mechanisms of antiparkinsonian dopamine D2-receptor subfamily agonists. Neurochem. Res 2003, 28, 1035–1040. [Google Scholar]
- Iida, M.; Miyazaki, I.; Tanaka, K.; Kabuto, H.; Iwata-Ichikawa, E.; Ogawa, N. Dopamine D2 receptor-mediated antioxidant and neuroprotective effects of ropinirole, a dopamine agonist. Brain Res 1999, 838, 51–59. [Google Scholar]
- Ogawa, N.; Tanaka, K.; Asanuma, M.; Kawai, M.; Masumizu, T.; Kohno, M.; Mori, A. Bromocriptine protects mice against 6-hydroxydopamine and scavenges hydroxyl free radicals in vitro. Brain Res 1994, 657, 207–213. [Google Scholar]
- Polydoro, M.; Schröder, N.; Lima, M.N.; Caldana, F.; Laranja, D.C.; Bromberg, E.; Roesler, R.; Quevedo, J.; Moreira, J.C.; Dal-Pizzol, F. Haloperidol- and clozapine-induced oxidative stress in the rat brain. Pharmacol. Biochem. Behav. 2004, 78, 751–756. [Google Scholar]
- Charvin, D.; Vanhoutte, P.; Pagès, C.; Borrelli, E.; Caboche, J. Unraveling a role for dopamine in Huntington’s disease: The dual role of reactive oxygen species and D2 receptor stimulation. Proc. Natl. Acad. Sci. USA 2005, 102, 12218–12223. [Google Scholar]
- Sawada, H.; Ibi, M.; Kihara, T.; Urushitani, M.; Akaike, A.; Kimura, J.; Shimohama, S. Dopamine D2-type agonists protect mesencephalic neurons from glutamate neurotoxicity: Mechanisms of neuroprotective treatment against oxidative stress. Ann. Neurol 1998, 44, 110–118. [Google Scholar]
- Takashima, H.; Tsujihata, M.; Kishikawa, M.; Freed, W.J. Bromocriptine protects dopaminergic neurons from levodopa-induced toxicity by stimulating D2 receptors. Exp. Neurol 1999, 159, 98–104. [Google Scholar]
- Cuevas, S.; Zhang, Y.; Yang, Y.; Escano, C.; Asico, L.; Jones, J.E.; Armando, I.; Jose, P.A. Role of renal DJ-1 in the pathogenesis of hypertension associated with increased reactive oxygen species production. Hypertension 2012, 59, 446–452. [Google Scholar]
- Yang, Y.; Zhang, Y.; Cuevas, S.; Villar, V.A.; Escano, C.; Asico, L.D.; Yu, P.; Grandy, D.K.; Felder, R.A.; Armando, I.; et al. Paraoxonase 2 decreases renal reactive oxygen species production, lowers blood pressure, and mediates dopamine D2 receptor-induced inhibition of NADPH oxidase. Free Radic. Biol. Med 2012, 53, 437–446. [Google Scholar]
- Ueda, A.; Ozono, R.; Oshima, T.; Yano, A.; Kambe, M.; Teranishi, Y.; Katsuki, M.; Chayama, K. Disruption of the type 2 dopamine receptor gene causes a sodium-dependent increase in blood pressure in mice. Am. J. Hypertens 2003, 16, 853–858. [Google Scholar]
- Armando, I.; Wang, X.; Villar, V.A.; Jones, J.E.; Asico, L.D.; Escano, C.; Jose, P.A. Reactive oxygen species-dependent hypertension in dopamine D2 receptor-deficient mice. Hypertension 2007, 49, 672–678. [Google Scholar]
- Bellner, L.; Martinelli, L.; Halilovic, A.; Patil, K.; Puri, N.; Dunn, M.W.; Regan, R.F.; Schwartzman, M.L. Heme oxygenase-2 deletion causes endothelial cell activation marked by oxidative stress, inflammation, and angiogenesis. J. Pharmacol. Exp. Ther 2009, 331, 925–932. [Google Scholar]
- Nagakubo, D.; Taira, T.; Kitaura, H.; Ikeda, M.; Tamai, K.; Iguchi-Ariga, S.M.; Ariga, H. DJ-1, a novel oncogene which transforms mouse NIH3T3 cells in cooperation with ras. Biochem. Biophys. Res. Commun 1997, 231, 509–513. [Google Scholar]
- Martinat, C.; Shendelman, S.; Jonason, A.; Leete, T.; Beal, M.F.; Yang, L.; Floss, T.; Abeliovich, A. Sensitivity to oxidative stress in DJ-1-deficient dopamine neurons: An ES- derived cell model of primary Parkinsonism. PLoS Biol 2004, 2, e327. [Google Scholar]
- Taira, T.; Saito, Y.; Niki, T.; Iguchi-Ariga, S.M.; Takahashi, K.; Ariga, H. DJ-1 has a role in antioxidative stress to prevent cell death. EMBO Rep 2004, 5, 213–218. [Google Scholar]
- Aleyasin, H.; Rousseaux, M.W.; Phillips, M.; Kim, R.H.; Bland, R.J.; Callaghan, S.; Slack, R.S.; During, M.J.; Mak, T.W.; Park, D.S. The Parkinson’s disease gene DJ-1 is also a key regulator of stroke-induced damage. Proc. Natl. Acad. Sci. USA 2007, 104, 18748–18753. [Google Scholar]
- Andres-Mateos, E.; Perier, C.; Zhang, L.; Blanchard-Fillion, B.; Greco, T.M.; Thomas, B.; Ko, H.S.; Sasaki, M.; Ischiropoulos, H.; Przedborski, S.; et al. DJ-1 gene deletion reveals that DJ-1 is an atypical peroxiredoxin-like peroxidase. Proc. Natl. Acad. Sci. USA 2007, 104, 14807–14812. [Google Scholar]
- Liu, F.; Nguyen, J.L.; Hulleman, J.D.; Li, L.; Rochet, J.C. Mechanisms of DJ-1 neuroprotection in a cellular model of Parkinson’s disease. J. Neurochem. 2008, 105, 2435–2453. [Google Scholar]
- Zhou, W.; Freed, C.R. DJ-1 up-regulates glutathione synthesis during oxidative stress and inhibits A53T alpha-synuclein toxicity. J. Biol. Chem 2005, 280, 43150–43158. [Google Scholar]
- Zhong, N.; Xu, J. Synergistic activation of the human MnSOD promoter by DJ-1 and PGC-1alpha: Regulation by SUMOylation and oxidation. Hum. Mol. Genet 2008, 17, 3357–3367. [Google Scholar]
- Vasseur, S.; Afzal, S.; Tardivel-Lacombe, J.; Park, D.S.; Iovanna, J.L.; Mak, T.W. DJ-1/PARK7 is an important mediator of hypoxia-induced cellular responses. Proc. Natl. Acad. Sci. USA 2009, 106, 1111–1116. [Google Scholar]
- Fan, J.; Ren, H.; Jia, N.; Fei, E.; Zhou, T.; Jiang, P.; Wu, M.; Wang, G. DJ-1 decreases Bax expression through repressing p53 transcriptional activity. J. Biol. Chem 2008, 283, 4022–4030. [Google Scholar]
- Van der Brug, M.P.; Blackinton, J.; Chandran, J.; Hao, L.Y.; Lal, A.; Mazan-Mamczarz, K.; Martindale, J.; Xie, C.; Ahmad, R.; Thomas, K.J.; et al. RNA binding activity of the recessive parkinsonism protein DJ-1 supports involvement in multiple cellular pathways. Proc. Natl. Acad. Sci. USA 2008, 105, 10244–10249. [Google Scholar]
- Mullett, S.J.; di Maio, R.; Greenamyre, J.T.; Hinkle, D.A. DJ-1 expression modulates astrocyte-mediated protection against neuronal oxidative stress. J. Mol. Neurosci 2013, 49, 507–511. [Google Scholar]
- Lev, N.; Barhum, Y.; Ben-Zur, T.; Melamed, E.; Steiner, I.; Offen, D. Knocking out DJ-1 attenuates astrocytes neuroprotection against 6-hydroxydopamine toxicity. J. Mol. Neurosci 2013, 50, 542–550. [Google Scholar]
- Sun, S.Y.; An, C.N.; Pu, X.P. DJ-1 protein protects dopaminergic neurons against 6-OHDA/MG-132-induced neurotoxicity in rats. Brain Res. Bull 2012, 88, 609–616. [Google Scholar]
- Nguyen, T.A.; Frank-Cannon, T.; Martinez, T.N.; Ruhn, K.A.; Marvin, M.; Casey, B.; Treviño, I.; Hong, J.J.; Goldberg, M.S.; Tansey, M.G. Analysis of inflammation-related nigral degeneration and locomotor function in DJ-1−/− mice. J. Neuroinflamm 2013, 10, 50. [Google Scholar]
- Goldberg, M.S.; Pisani, A.; Haburcak, M.; Vortherms, T.A.; Kitada, T.; Costa, C.; Tong, Y.; Martella, G.; Tscherter, A.; Martins, A.; et al. Nigrostriatal dopaminergic deficits and hypokinesia caused by inactivation of the familial Parkinsonism-linked gene DJ-1. Neuron 2005, 45, 489–496. [Google Scholar]
- Billia, F.; Hauck, L.; Grothe, D.; Konecny, F.; Rao, V.; Kim, R.H.; Mak, T.W. Parkinson-susceptibility gene DJ-1/PARK7 protects the murine heart from oxidative damage in vivo. Proc. Natl. Acad. Sci. USA 2013, 110, 6085–6090. [Google Scholar]
- Xu, X.; Martin, F.; Friedman, J.S. The familial Parkinson’s disease gene DJ-1 (PARK7) is expressed in red cells and plays a role in protection against oxidative damage. Blood Cells Mol. Dis 2010, 45, 227–232. [Google Scholar]
- Inberg, A.; Linial, M. Protection of pancreatic β-cells from various stress conditions is mediated by DJ-1. J. Biol. Chem 2010, 285, 25686–25698. [Google Scholar]
- Ng, C.J.; Hama, S.Y.; Bourquard, N.; Navab, M.; Reddy, S.T. Adenovirus mediated expression of human paraoxonase 2 protects against the development of atherosclerosis in apolipoprotein E-deficient mice. Mol. Genet. Metab 2006, 89, 368–373. [Google Scholar]
- Rajkovic, M.G.; Rumora, L.; Barisic, K. The paraoxonase 1, 2 and 3 in humans. Biochem. Med 2011, 21, 122–130. [Google Scholar]
- Ng, C.J.; Wadleigh, D.J.; Gangopadhyay, A.; Hama, S.; Grijalva, V.R.; Navab, M.; Fogelman, A.M.; Reddy, S.T. Paraoxonase-2 is a ubiquitously expressed protein with antioxidant properties and is capable of preventing cell-mediated oxidative modification of low density lipoprotein. J. Biol. Chem 2001, 276, 44444–44449. [Google Scholar]
- Fuhrman, B.; Khateeb, J.; Shiner, M.; Nitzan, O.; Karry, R.; Volkova, N.; Aviram, M. Urokinase plasminogen activator upregulates paraoxonase 2 expression in macrophages via an NADPH oxidase-dependent mechanism. Arterioscler. Thromb. Vasc. Biol 2008, 28, 1361–1367. [Google Scholar]
- Horke, S.; Witte, I.; Wilgenbus, P.; Krüger, M.; Strand, D.; Förstermann, U. Paraoxonase-2 reduces oxidative stress in vascular cells and decreases endoplasmic reticulum stress-induced caspase activation. Circulation 2007, 115, 2055–2064. [Google Scholar]
- Ng, C.J.; Bourquard, N.; Hama, S.; Shih, D.M.; Grijalva, V.; Navab, M.; Fogelman, A.M.; Reddy, S.T. Adenovirus-mediated expression of human paraoxonase 3 protects against the progression of atherosclerosis in apolipoprotein E-deficient mice. Arterioscler. Thromb. Vasc. Biol 2007, 27, 1368–13674. [Google Scholar]
- Shiner, M.; Fuhrman, B.; Aviram, M. Paraoxonase 2 (PON2) expression is upregulated via a reduced-nicotinamide-adenine-dinucleotide-phosphate (NADPH)-oxidase-dependent mechanism during monocytes differentiation into macrophages. Free Radic. Biol. Med 2004, 37, 2052–2063. [Google Scholar]
- Reddy, S.T.; Devarajan, A.; Bourquard, N.; Shih, D.; Fogelman, AM. Is it just paraoxonase 1 or are other members of the paraoxonase gene family implicated in atherosclerosis? Curr. Opin. Lipidol. 2008, 19, 405–408. [Google Scholar]
- Larouche, A.; Berube, P.; Sarret, P.; Grignon, S. Subacute H2O2, but not poly(IC), upregulates dopamine D2 receptors in retinoic acid differentiated SH-SY5Y neuroblastoma. Synapse 2008, 62, 70–73. [Google Scholar]
- Cachofeiro, V.; Goicochea, M.; de Vinuesa, S.G.; Oubiña, P.; Lahera, V.; Luño, J. Oxidative stress and inflammation, a link between chronic kidney disease and cardiovascular disease. Kidney Int 2008, 74, S4–S9. [Google Scholar]
- Bowie, A.; O’Neill, L.A. Oxidative stress and nuclear factor-κB activation: a reassessment of the evidence in the light of recent discoveries. Biochem. Pharmacol 2000, 59, 13–23. [Google Scholar]
- Besser, M.J.; Ganor, Y.; Levite, M. Dopamine by itself activates either D2, D3 or D1/D5 dopaminergic receptors in normal human T-cells and triggers the selective secretion of either IL-10, TNFαor both. J. Neuroimmunol 2005, 169, 161–171. [Google Scholar]
- Laengle, U.W.; Markstein, R.; Pralet, D.; Seewald, W.; Roman, D. Effect of GLC756, a novel mixed dopamine D1 receptor antagonist and dopamine D2 receptor agonist, on TNF-α release in vitro from activated rat mast cells. Exp. Eye Res 2006, 83, 1335–1339. [Google Scholar]
- Zhang, Y.; Cuevas, S.; Asico, L.D.; Escano, C.; Yang, Y.; Pascua, A.M.; Wang, X.; Jones, J.E.; Grandy, D.; Eisner, G.; et al. Deficient dopamine D2 receptor function causes renal inflammation independently of high blood pressure. PLoS One 2012, 7, e38745. [Google Scholar]
- Laaksoa, A.; Pohjalainena, T.; Bergmanb, J.; Kajandera, J.; Haaparantac, M.; Solinb, O.; Syvalahtia, E.; Hietalad, J. The A1 allele of the human D2 dopamine receptor gene is associated with increased activity of striatal l-amino acid decarboxylase in healthy subjects. Pharmacogenet. Genomics 2005, 15, 387–391. [Google Scholar]
- Ritchie, T.; Noble, E.P. Association of seven polymorphisms of the D2 dopamine receptor gene with brain receptor-binding characteristics. Neurochem. Res 2003, 28, 73–82. [Google Scholar]
- Rosmond, R.; Rankinen, T.; Chagnon, M.; Perusse, L.; Chagnon, Y.C.; Bouchard, C.; Bjorntorp, P. Polymorphism in exon 6 of the dopamine D2 receptor gene (DRD2) is associated with elevated blood pressure and personality disorders in men. J. Hum. Hypertens 2001, 15, 553–558. [Google Scholar]
- Thomas, G.N.; Tomlinson, B.; Critchley, J.A. Modulation of blood pressure and obesity with the dopamine D2 receptor gene Taq1 polymorphism. Hypertension 2000, 36, 177–182. [Google Scholar]
- Carvey, P.M.; McGuire, S.O.; Ling, Z.D. Neuroprotective effects of D3 dopamine receptor agonists. Parkinsonism Relat. Disord 2001, 7, 213–223. [Google Scholar]
- Zou, L.; Xu, J.; Jankovic, J.; He, Y.; Appel, S.H.; Le, W. Pramipexole inhibits lipid peroxidation and reduces injury in the substantia nigra induced by the dopaminergic neurotoxin 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine in C57BL/6 mice. Neurosci. Lett 2000, 281, 167–170. [Google Scholar]
- Le, W.D.; Jankovic, J.; Xie, W.; Appel, S.H. Antioxidant property of pramipexole independent of dopamine receptor activation in neuroprotection. J. Neural Transm 2000, 107, 1165–1173. [Google Scholar]
- Ferrari-Toninelli, G.; Maccarinelli, G.; Uberti, D.; Buerger, E.; Memo, M. Mitochondria-targeted antioxidant effects of S(−) and R(+) pramipexole. BMC Pharmacol 2010, 10, 2. [Google Scholar]
- Li, C.; Biswas, S.; Li, X.; Dutta, A.K.; Le, W. Novel D3 dopamine receptor-preferring agonist D-264: Evidence of neuroprotective property in Parkinson’s disease animal models induced by 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine and lactacystin. J. Neurosci. Res 2010, 88, 2513–2523. [Google Scholar]
- Asico, L.D.; Ladines, C.; Fuchs, S.; Accili, D.; Carey, R.M.; Semeraro, C.; Pocchiari, F.; Felder, R.A.; Eisner, G.M.; Jose, P.A. Disruption of the dopamine D3 receptor gene produces renin-dependent hypertension. J. Clin. Invest 1998, 102, 493–498. [Google Scholar]
- Wang, X.; Escano, C.S.; Asico, L.; Jones, J.E.; Barte, A.; Lau, Y.S.; Jose, P.A.; Armando, I. Upregulation of renal D5 dopamine receptor ameliorates the hypertension in D3 dopamine receptor-deficient mice. Hypertension 2013, 62, 295–301. [Google Scholar]
- Gribkoff, V.K.; Bozik, M.E. KNS-760704 [(6R)-4,5,6,7-tetrahydro-N6-propyl-2, 6-benzothiazole-diamine dihydrochloride monohydrate] for the treatment of amyotrophic lateral sclerosis. CNS Neurosci. Ther 2008, 14, 215–226. [Google Scholar]
- Okada, Y.; Sakai, H.; Kohiki, E.; Suga, E.; Yanagisawa, Y.; Tanaka, K.; Hadano, S.; Osuga, H.; Ikeda, J.E. A dopamine D4 receptor antagonist attenuates ischemia-induced neuronal cell damage via upregulation of neuronal apoptosis inhibitory protein. J. Cereb. Blood. Flow Metab 2005, 25, 794–806. [Google Scholar]
- Shimada, S.; Hirabayashi, M.; Ishige, K.; Kosuge, Y.; Kihara, T.; Ito, Y. Activation of dopamine D4 receptors is protective against hypoxia/reoxygenation-induced cell death in HT22 cells. J. Pharmacol. Sci 2010, 114, 217–224. [Google Scholar]
- Ishige, K.; Chen, Q.; Sagara, Y.; Schubert, D. The activation of dopamine D4 receptors inhibits oxidative stress-induced nerve cell death. J. Neurosci 2001, 21, 6069–6076. [Google Scholar]
- Bek, M.J.; Wang, X.; Asico, L.D.; Jones, J.E.; Zheng, S.; Li, X.; Eisner, G.M.; Grandy, D.K.; Carey, R.M.; Soares-da-Silva, P.; et al. Angiotensin-II type 1 receptor-mediated hypertension in D4 dopamine receptor-deficient.mice. Hypertension 2006, 47, 288–295. [Google Scholar]
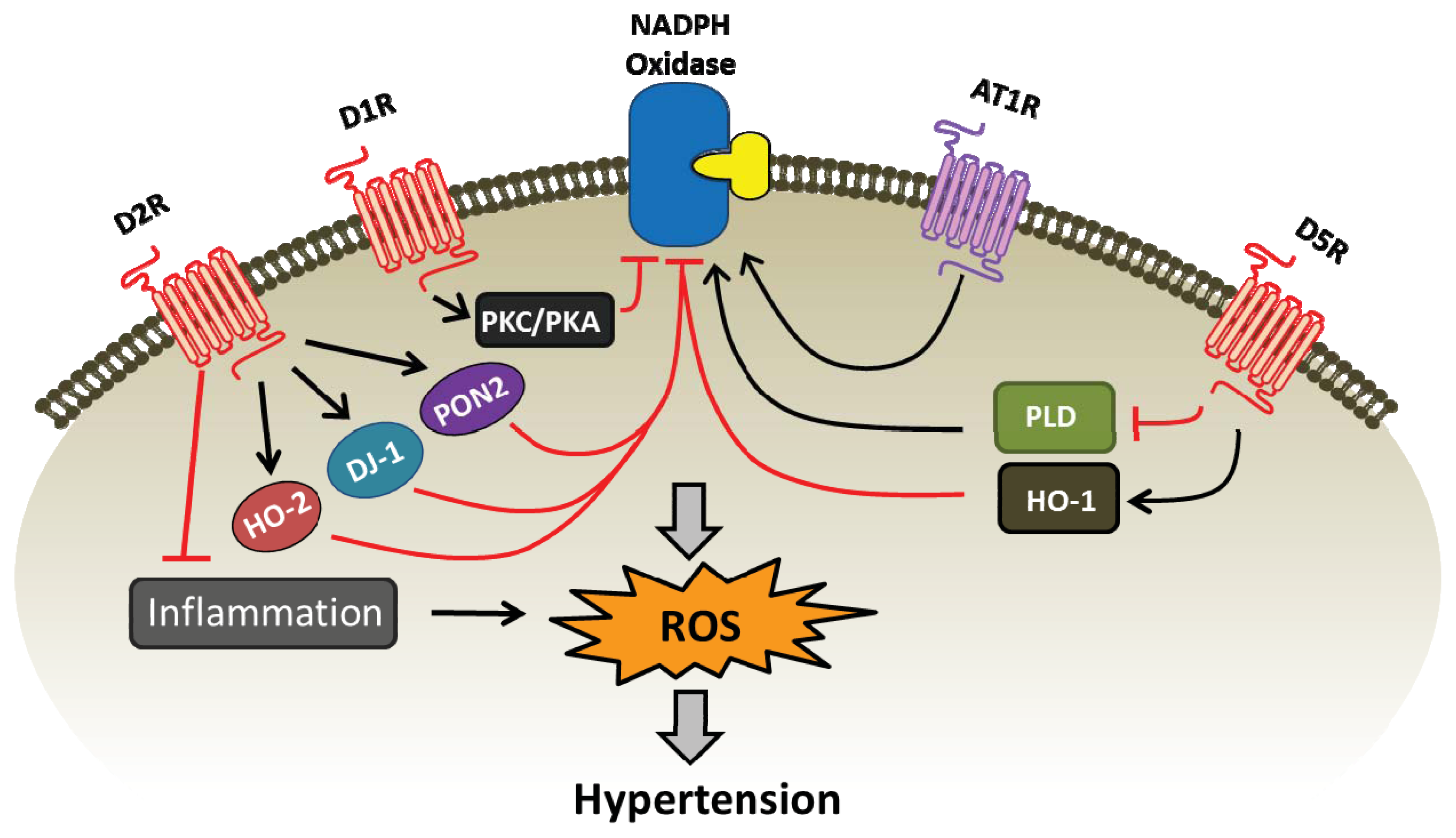

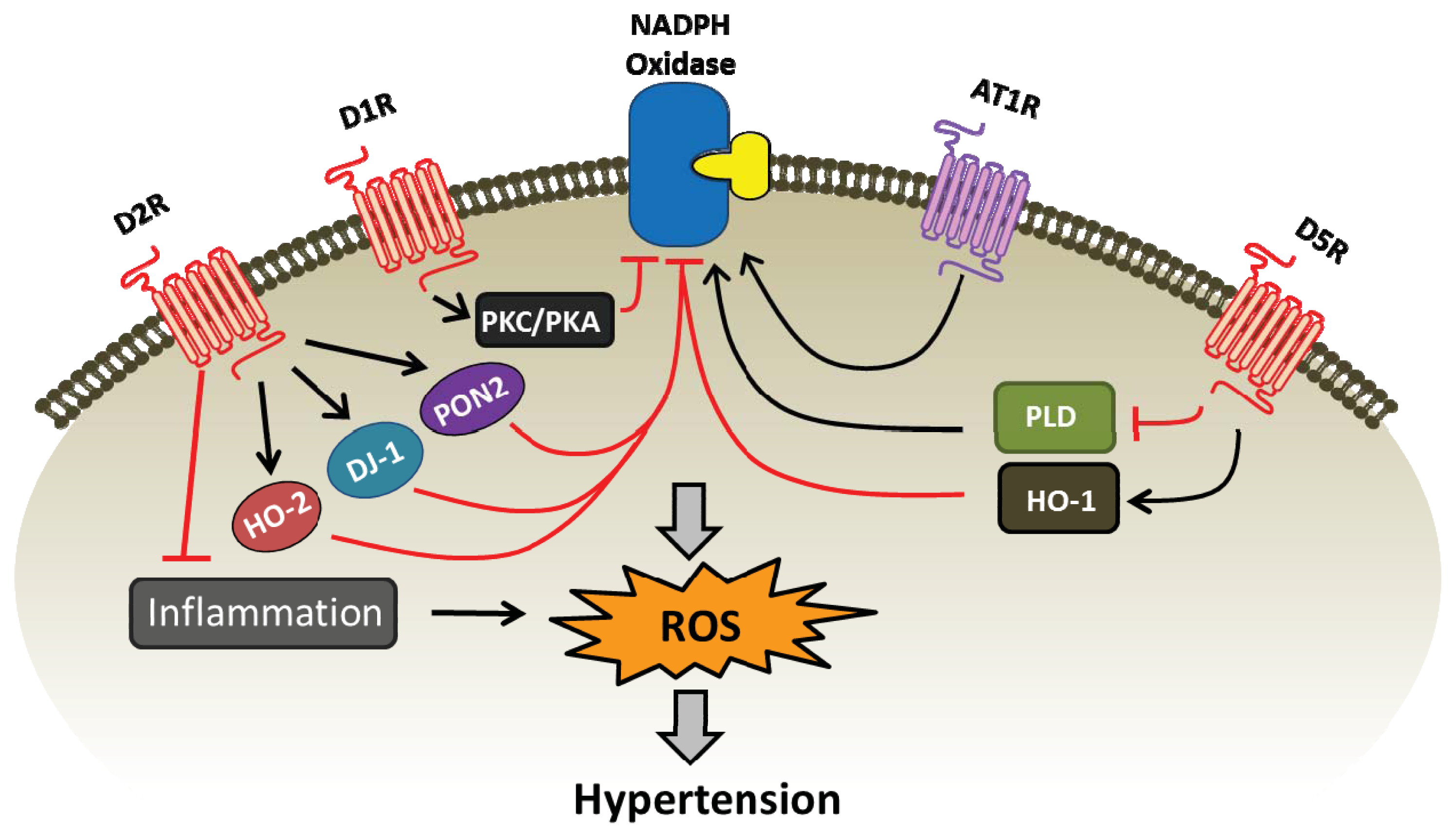{kind=link}
| Dopamine receptor subtype | Pro-oxidant enzymes (inhibition) | Anti-oxidant enzymes (stimulation) |
|---|---|---|
| D1R | NADPH oxidase, via PKA/PKC cross talk [19,45,47] | SOD, glutathione peroxidase, glutamyl cysteine transferase, and HO-1 [46,65] |
| D2R | NADPH oxidase [19,85,86,88] | DJ-1, PON2, and HO-2 [19,85,86] glutathione, catalase, and SOD [79] |
| D5R | NADPH oxidase, via PLD2 [43,49] | SOD, glutathione peroxidase, glutamyl cysteine transferase, and HO-1 [46,48] |
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Cuevas, S.; Villar, V.A.; Jose, P.A.; Armando, I. Renal Dopamine Receptors, Oxidative Stress, and Hypertension. Int. J. Mol. Sci. 2013, 14, 17553-17572. https://doi.org/10.3390/ijms140917553
Cuevas S, Villar VA, Jose PA, Armando I. Renal Dopamine Receptors, Oxidative Stress, and Hypertension. International Journal of Molecular Sciences. 2013; 14(9):17553-17572. https://doi.org/10.3390/ijms140917553
Chicago/Turabian StyleCuevas, Santiago, Van Anthony Villar, Pedro A. Jose, and Ines Armando. 2013. "Renal Dopamine Receptors, Oxidative Stress, and Hypertension" International Journal of Molecular Sciences 14, no. 9: 17553-17572. https://doi.org/10.3390/ijms140917553