Green Leaf Volatiles: A Plant’s Multifunctional Weapon against Herbivores and Pathogens


{kind=link}
{kind=link}
Abstract
:1. Introduction
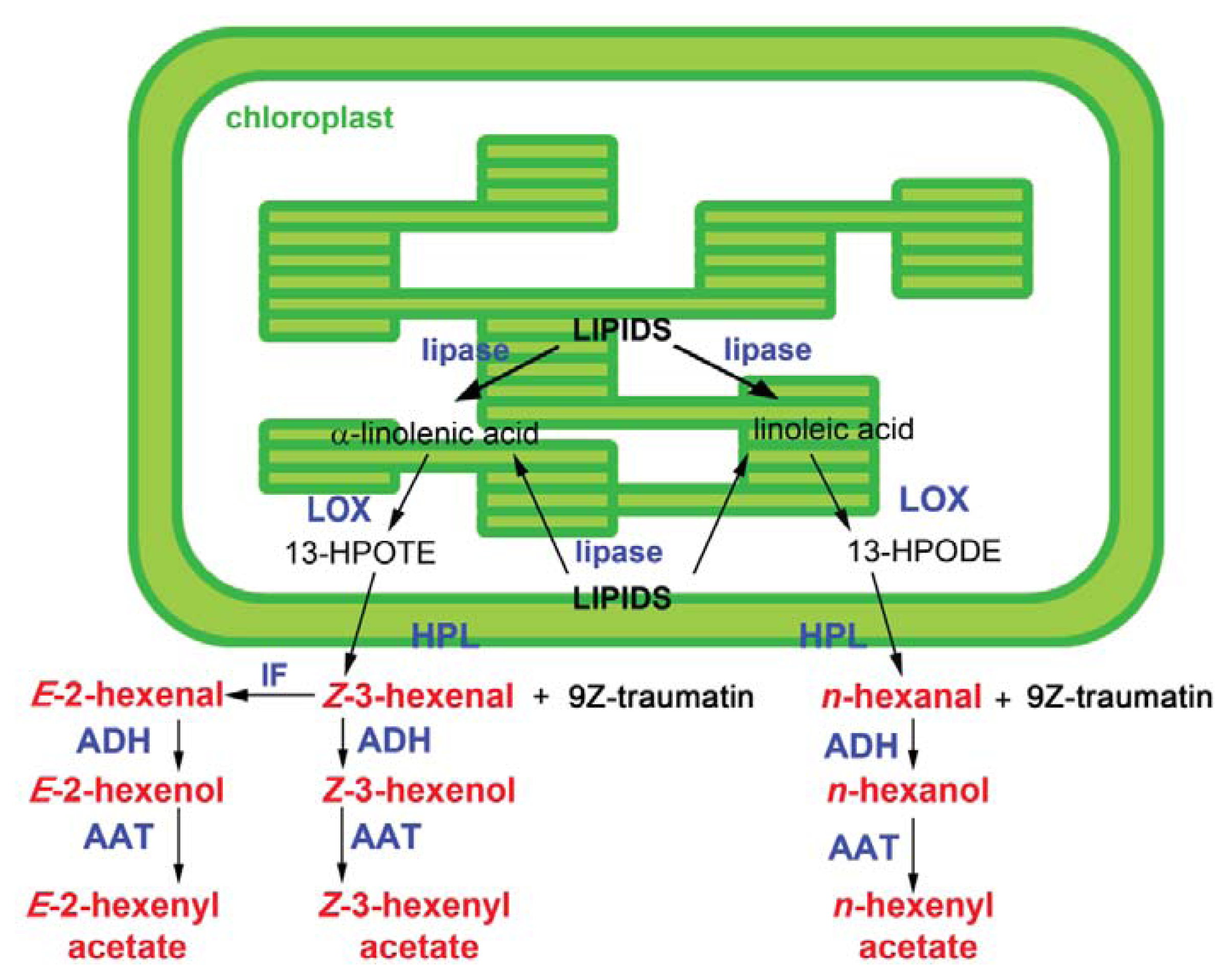
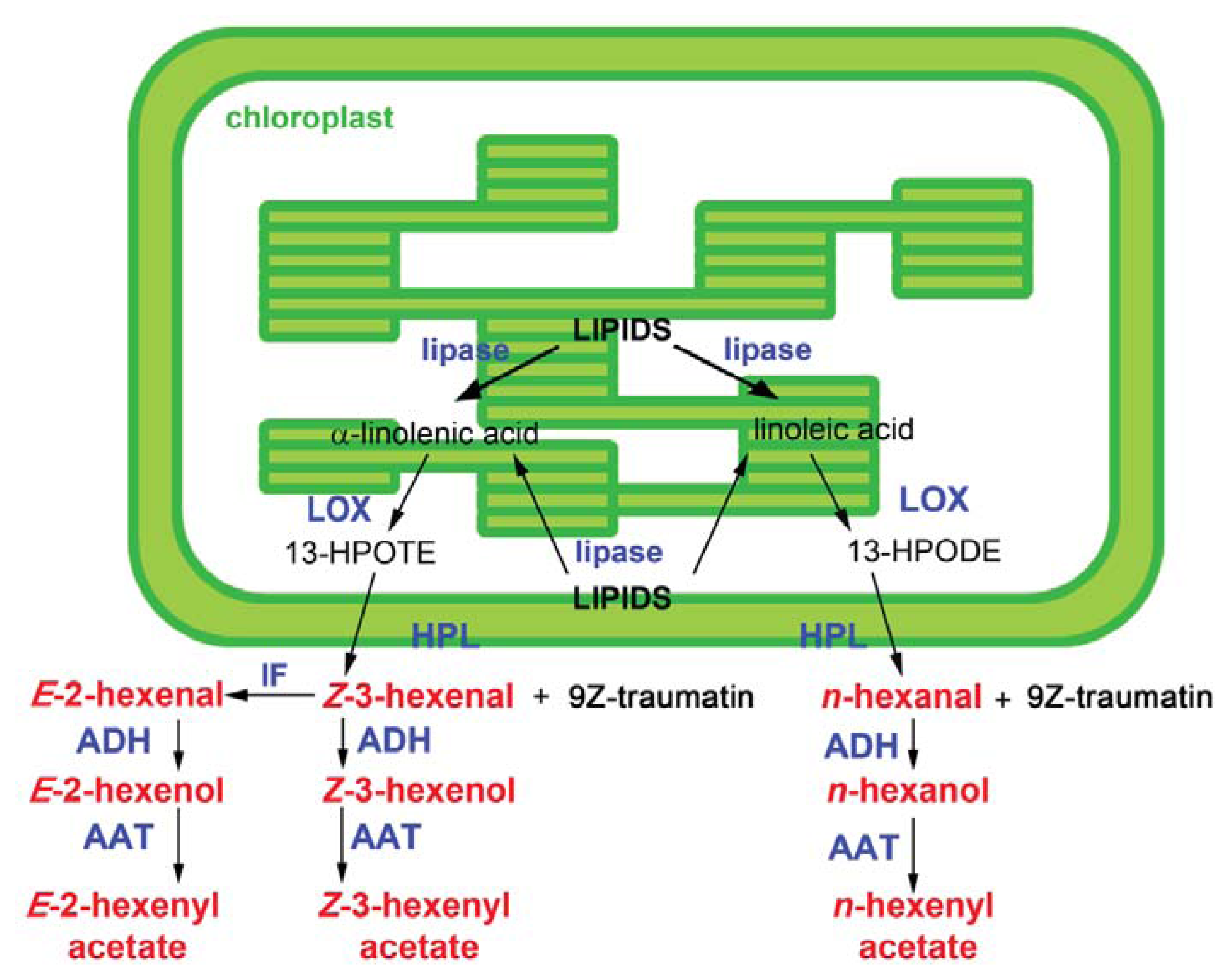
2. Green Leaf Volatile Biosynthesis
2.1. The Close Link between GLVs and JA: Substrate Competition or Different Regulation?
2.2. GLVs Are Emitted after Abiotic Stresses
2.3. GLVs Are Detoxified in Different Ways
3. Phytohormonal Crosstalk Orchestrates Plant Defenses
3.1. GLVs and JA
3.2. GLVs and Ethylene
4. GLVs and Plant Pathogens
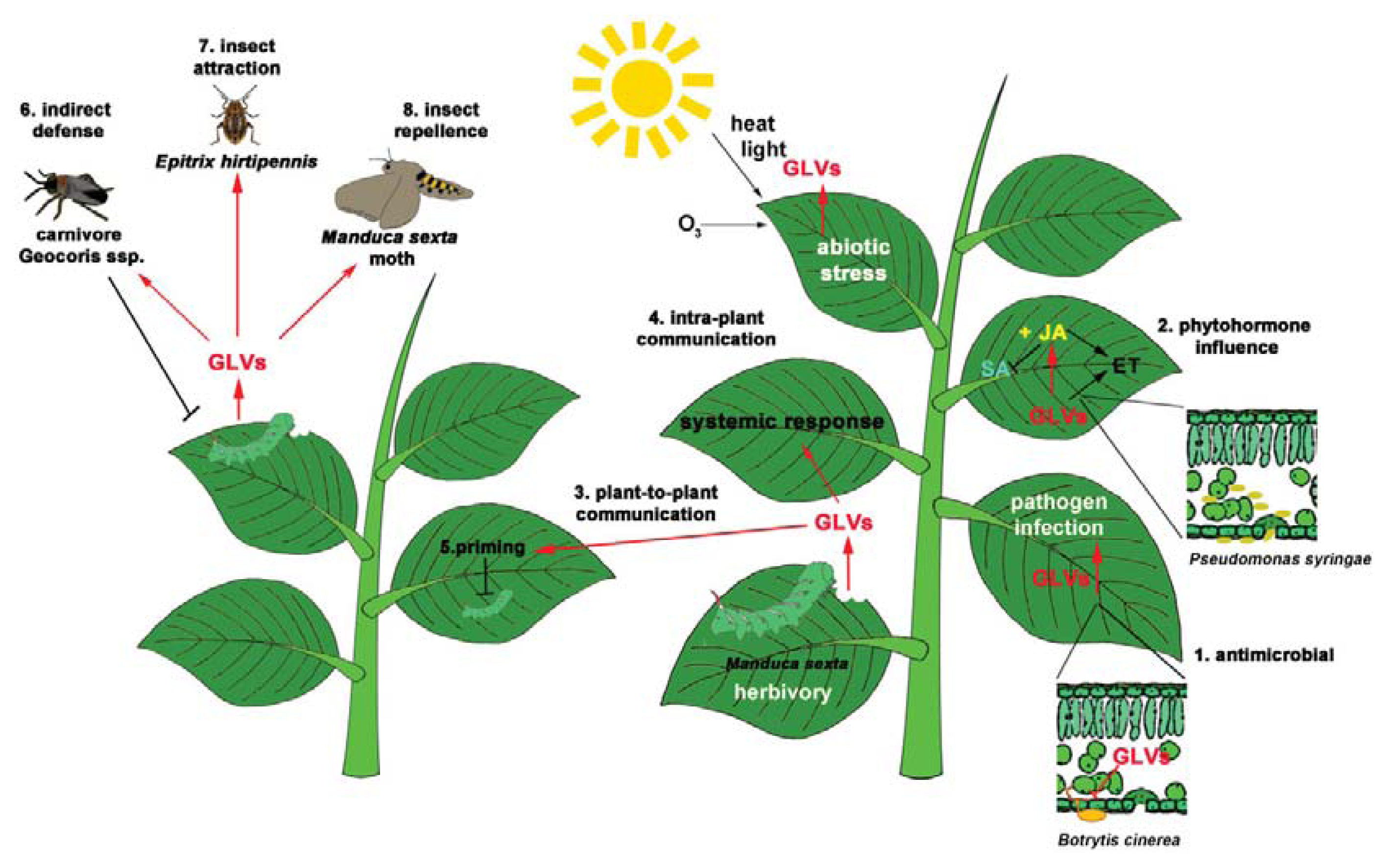
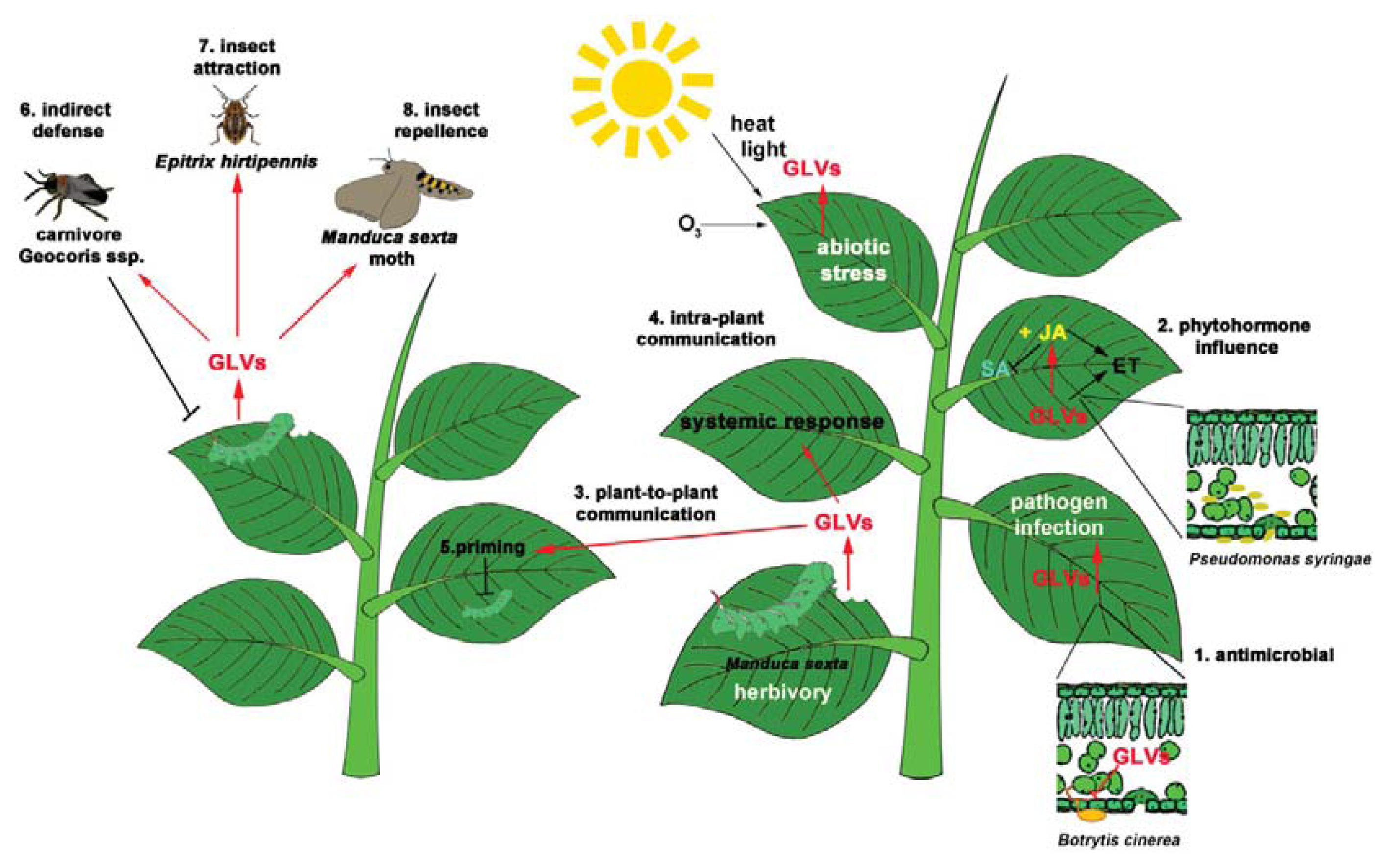
5. GLVs and Insects
5.1. Beneficial Insects: Indirect Defenses
5.2. Non-Beneficial Insects
5.2.1. GLVs Are Attractive to Insects
5.2.2. GLVs Are Repellent to Insects
6. GLV Perception, Downstream Signaling and “Plant Communication”
Priming by GLVs: Staying Ready for the Battle
7. Conclusions
Acknowledgments
Conflicts of Interest
References
- Matsui, K. Green leaf volatiles: Hydroperoxide lyase pathway of oxylipin metabolism. Curr. Opin. Plant Biol 2006, 9, 274–280. [Google Scholar]
- Hatanaka, A. The biogeneration of green odour by green leaves. Phytochemistry 1993, 34, 1201–1218. [Google Scholar]
- Halitschke, R.; Ziegler, J.; Keinänen, M.; Baldwin, I.T. Silencing of hydroperoxide lyase and allene oxide synthase reveals substrate and defense signaling crosstalk in Nicotiana attenuata. Plant J 2004, 40, 35–46. [Google Scholar]
- Gomi, K.; Yamasaki, Y.; Yamamoto, H.; Akimitsu, K. Characterization of a hydroperoxide lyase gene and effect of C6-volatiles on expression of genes of the oxylipin metabolism in Citrus. J. Plant Physiol 2003, 160, 1219–1231. [Google Scholar]
- Brilli, F.; Ruuskanen, T.M.; Schnitzhofer, R.; Müller, M.; Breitenlechner, M.; Bittner, V.; Wohlfahrt, G.; Loreto, F.; Hansel, A. Detection of plant volatiles after leaf wounding and darkening by proton transfer reaction “time-of-flight” mass spectrometry (PTR-TOF). PLoS One 2011, 6, e20419. [Google Scholar]
- Fall, R.; Karl, T.; Hansel, A.; Jordan, A.; Lindiger, W. Volatile organic compounds emitted after leaf wounding: On-line analysis by proton-transfer-reaction mass spectrometry. J. Geophys 1999, 104, 963–974. [Google Scholar]
- Turlings, T.; Loughrin, J. How caterpillar-damaged plants protect themselves by attracting parasitic wasps. Proc. Natl. Acad. Sci. USA 1995, 92, 4169–4174. [Google Scholar]
- Croft, K.; Juttner, F.; Slusarenko, A. Volatile products of the lipoxygenase pathway evolved from phaseolus vulgaris (1.) leaves lnoculated with pseudomonas syringae pv phaseolicola. Plant Physiol 1993, 101, 13–24. [Google Scholar]
- Heiden, A.C.; Kobel, K.; Langebartels, C. Emissions of oxygenated volatile organic compounds from plants Part I: Emissions from lipoxygenase activity. J. Atmos. Chem 2003, 45, 143–172. [Google Scholar]
- Shiojiri, K.; Kishimoto, K.; Ozawa, R.; Kugimiya, S.; Urashimo, S.; Arimura, G.; Horiuchi, J.; Nishioka, T.; Matsui, K.; Takabayashi, J. Changing green leaf volatile biosynthesis in plants: An approach for improving plant resistance against both herbivores and pathogens. Proc. Natl. Acad. Sci. USA 2006, 103, 16672–16676. [Google Scholar]
- Gouinguené, S.; Turlings, T. The effects of abiotic factors on induced volatile emissions in corn plants. Plant Physiol 2002, 129, 1296–1307. [Google Scholar]
- Allmann, S.; Baldwin, I.T. Insects betray themselves in nature to predators by rapid isomerization of green leaf volatiles. Science 2010, 329, 1075–1078. [Google Scholar]
- D’Auria, J.C.; Pichersky, E.; Schaub, A.; Hansel, A.; Gershenzon, J. Characterization of a BAHD acyltransferase responsible for producing the green leaf volatile (Z)-3-hexen-1-yl acetate in Arabidopsis thaliana. Plant J 2007, 49, 194–207. [Google Scholar]
- Loughrin, J.; Manukian, A. Volatiles emitted by different cotton varieties damaged by feeding beet armyworm larvae. J. Chem.Ecol 1995, 21, 1217–1227. [Google Scholar]
- Birkett, M.A.; Chamberlain, K.; Guerrieri, E.; Pickett, J.A.; Wadhams, L.J.; Yasuda, T. Volatiles from whitefly-infested plants elicit a host-locating response in the parasitoid, Encarsia formosa. J. Chem. Ecol 2003, 29, 1589–1600. [Google Scholar]
- Gouinguené, S.; Pickett, J.A.; Wadhams, L.J.; Birkett, M.A.; Turlings, T.C.J. Antennal electrophysiological responses of three parasitic wasps to caterpillar-induced volatiles from maize (Zea mays mays), cotton (Gossypium herbaceum), and cowpea (Vigna unguiculata). J. Chem. Ecol 2005, 31, 1023–1038. [Google Scholar]
- Kessler, A.; Baldwin, I.T. Defensive function of herbivore-induced plant volatile emissions in nature. Science 2001, 291, 2141–2144. [Google Scholar]
- Heil, M.; Karban, R. Explaining evolution of plant communication by airborne signals. Trends Ecol. Evolut 2010, 25, 137–144. [Google Scholar]
- Baldwin, I.T.; Halitschke, R.; Paschold, A.; von Dahl, C.C.; Preston, C.A. Volatile signaling in plant-plant interactions: “Talking trees” in the genomics era. Science 2006, 311, 812–815. [Google Scholar]
- Farmer, E.E. Surface-to-air signals. Nature 2001, 411, 854–856. [Google Scholar]
- Frost, C.J.; Mescher, M.C.; Carlson, J.E.; de Moraes, C.M. Plant defense priming against herbivores: Getting ready for a different battle. Plant Physiol 2008, 146, 818–824. [Google Scholar]
- Christensen, S.; Nemchenko, A.; Borrego, E.; Murray, I.; Sobhy, I.S.; Bosak, L.; Deblasio, S.; Erb, M.; Robert, C.A.M.; Vaughn, K.A.; et al. The maize lipoxygenase, ZmLOX10, mediates green leaf volatile, jasmonate and herbivore-induced plant volatile production for defense against insect attack. Plant J 2013, 74, 59–73. [Google Scholar]
- Frost, C.J.; Mescher, M.C.; Dervinis, C.; Davis, J.M.; Carlson, J.E.; de Moraes, C.M. Priming defense genes and metabolites in hybrid poplar by the green leaf volatile cis-3-hexenyl acetate. New Phytol 2008, 180, 722–734. [Google Scholar]
- Liu, X.; Li, F.; Tang, J.; Wang, W.; Zhang, F.; Wang, G.; Chu, J.; Yan, C.; Wang, T.; Chu, C.; et al. Activation of the Jasmonic Acid Pathway by Depletion of the Hydroperoxide Lyase OsHPL3 Reveals Crosstalk between the HPL and AOS Branches of the Oxylipin Pathway in Rice. PLoS One 2012, 7, e50089. [Google Scholar]
- Engelberth, J.; Seidl-Adams, I.; Schultz, J.C.; Tumlinson, J.H. Insect elicitors and exposure to green leafy volatiles differentially upregulate major octadecanoids and transcripts of 12-oxo phytodienoic acid reductases in Zea mays. Mol. Plant Microbe Interact 2007, 20, 707–716. [Google Scholar]
- Scala, A.; Mirabella, R.; Mugo, C.; Matsui, K.; Haring, M.A.; Schuurink, R.C. E-2-hexenal promotes susceptibility to Pseudomonas syringae by activating jasmonic acid pathways in Arabidopsis. Front. Plant Sci 2013, 4, 1–11. [Google Scholar]
- Howe, G.A.; Schilmiller, A.L. Oxylipin metabolism in response to stress. Curr. Opin. Plant Biol. 2002, 5, 230–236. [Google Scholar]
- Matsui, K.; Kurishita, S.; Hisamitsu, A.; Kajiwara, T. A lipid-hydrolysing activity involved in hexenal formation. Biochem. Soc. Trans 2000, 28, 857–860. [Google Scholar]
- Kombrink, E. Chemical and genetic exploration of jasmonate biosynthesis and signaling paths. Planta 2012, 236, 1351–1366. [Google Scholar]
- Ellinger, D.; Stingl, N.; Kubigsteltig, I.I.; Bals, T.; Juenger, M.; Pollmann, S.; Berger, S.; Schuenemann, D.; Mueller, M.J. DONGLE and DEFECTIVE IN ANTHER DEHISCENCE1 lipases are not essential for wound- and pathogen-induced jasmonate biosynthesis: Redundant lipases contribute to jasmonate formation. Plant Physiol 2010, 153, 114–127. [Google Scholar]
- Bonaventure, G.; Schuck, S.; Baldwin, I.T. Revealing complexity and specificity in the activation of lipase-mediated oxylipin biosynthesis: A specific role of the Nicotiana attenuata GLA1 lipase in the activation of jasmonic acid biosynthesis in leaves and roots. Plant Cell Environ 2011, 34, 1507–1520. [Google Scholar]
- Qi, J.; Zhou, G.; Yang, L.; Erb, M.; Lu, Y.; Sun, X.; Cheng, J.; Lou, Y. The chloroplast-localized phospholipases D α4 and α5 regulate herbivore-induced direct and indirect defenses in rice. Plant Physiol 2011, 157, 1987–1999. [Google Scholar]
- León, J.; Royo, J.; Vancanneyt, G.; Sanz, C.; Silkowski, H.; Griffiths, G.; Sánchez-Serrano, J.J. Lipoxygenase H1 gene silencing reveals a specific role in supplying fatty acid hydroperoxides for aliphatic aldehyde production. J. Biol. Chem 2002, 277, 416–423. [Google Scholar]
- Griffiths, A.; Prestage, S.; Linforth, R.; Zhang, J.; Taylor, A.; Grierson, D. Fruit-specific lipoxygenase suppression in antisense-transgenic tomatoes. Postharvest Biol. Technol 1999, 17, 163–173. [Google Scholar]
- Allmann, S.; Halitschke, R.; Schuurink, R.C.; Baldwin, I.T. Oxylipin channelling in Nicotiana attenuata: Lipoxygenase 2 supplies substrates for green leaf volatile production. Plant Cell Environ 2010, 33, 2028–2040. [Google Scholar]
- Wang, R.; Shen, W.; Liu, L.; Jiang, L.; Liu, Y.; Su, N.; Wan, J. A novel lipoxygenase gene from developing rice seeds confers dual position specificity and responds to wounding and insect attack. Plant Mol. Biol 2008, 66, 401–414. [Google Scholar]
- Porta, H.; Figueroa-Balderas, R.E.; Rocha-Sosa, M. Wounding and pathogen infection induce a chloroplast-targeted lipoxygenase in the common bean (Phaseolus vulgaris L.). Planta 2008, 227, 363–373. [Google Scholar]
- Heitz, T.; Bergey, D.R.; Ryan, C.A. A gene encoding a chloroplast-targeted lipoxygenase in tomato leaves 1s transiently lnduced by wounding. Plant Physiol 1996, 114, 1085–1093. [Google Scholar]
- Salas, J.J.; Sánchez, C.; García-González, D.L.; Aparicio, R. Impact of the suppression of lipoxygenase and hydroperoxide lyase on the quality of the green odor in green leaves. J. Agric. Food Chem 2005, 53, 1648–1655. [Google Scholar]
- Bannenberg, G.; Martínez, M.; Hamberg, M.; Castresana, C. Diversity of the enzymatic activity in the lipoxygenase gene family of Arabidopsis thaliana. Lipids 2009, 44, 85–95. [Google Scholar]
- Fammartino, A.; Cardinale, F.; Göbel, C.; Mène-Saffrané, L.; Fournier, J.; Feussner, I.; Esquerré-Tugayé, M.-T. Characterization of a divinyl ether biosynthetic pathway specifically associated with pathogenesis in tobacco. Plant Physiol 2007, 143, 378–388. [Google Scholar]
- Mita, G.; Quarta, A.; Fasano, P.; de Paolis, A.; di Sansebastiano, G.P.; Perrotta, C.; Iannacone, R.; Belfield, E.; Hughes, R.; Tsesmetzis, N.; et al. Molecular cloning and characterization of an almond 9-hydroperoxide lyase, a new CYP74 targeted to lipid bodies. J. Exp. Bot 2005, 56, 2321–2333. [Google Scholar]
- Hamberg, M. An epoxy alcohol synthase pathway in higher plants: Biosynthesis of antifungal trihydroxy oxylipins in leaves of potato. Lipids 1999, 34, 1131–1142. [Google Scholar]
- Huang, F.-C.; Schwab, W. Cloning and characterization of a 9-lipoxygenase gene induced by pathogen attack from Nicotiana benthamiana for biotechnological application. BMC Biotechnol. 2011, 11. [Google Scholar] [CrossRef]
- Gobel, C.; Feussner, I.; Rosahl, S. Lipid peroxidation during the hypersensitive response in potato in the absence of 9-lipoxygenases. J. Biol. Chem 2003, 278, 52834–52840. [Google Scholar]
- Fournier, J.; Pouhat, M.; de Biochirnie, L.; de Mddecine, F. Purification and characterization of elicitor-induced lipoxygenase in tobacco cells. Plant J 1991, 3, 63–70. [Google Scholar]
- Véronési, C.; Rickauer, M.; Fournier, J.; Pouénat, M. Lipoxygenase gene expression in the tobacco-phytophtora parasitica nicotianae Interaction. Plant Physiol 1996, 112, 997–1004. [Google Scholar]
- Kolomiets, M.V.; Chen, H.; Gladon, R.J.; Braun, E.J.; Hannapel, D.J. A leaf lipoxygenase of potato induced specifically by pathogen infection. Plant Physiol 2000, 124, 1121–1130. [Google Scholar]
- Fidantsef, A.; Bostock, R. Characterization of potato tuber lipoxygenase cDNAs and lipoxygenase expression in potato tubers and leaves. Physiol. Plant 1998, 102, 257–271. [Google Scholar]
- Göbel, C.; Feussner, I.; Schmidt, A.; Scheel, D.; Sanchez-Serrano, J.; Hamberg, M.; Rosahl, S. Oxylipin profiling reveals the preferential stimulation of the 9-lipoxygenase pathway in elicitor-treated potato cells. J. Biol. Chem 2001, 276, 6267–6273. [Google Scholar]
- Grechkin, A.N.; Fazliev, F.N.; Mukhtarova, L.S. The lipoxygenase pathway in garlic (Allium sativum L.) bulbs: Detection of the novel divinyl ether oxylipins. FEBS Lett 1995, 371, 159–162. [Google Scholar]
- Hamberg, M. A pathway for biosynthesis of divinyl ether fatty acids in green leaves. Lipids 1998, 33, 1061–1071. [Google Scholar]
- Katsir, L.; Chung, H.S.; Koo, A.J.K.; Howe, G.A. Jasmonate signaling: A conserved mechanism of hormone sensing. Curr. Opin. Plant Biol 2008, 11, 428–435. [Google Scholar]
- Piazza, G.J.; Nuñez, A.; Foglia, T.A. Isolation of unsaturated diols after oxidation of conjugated linoleic acid with peroxygenase. Lipids 2003, 38, 255–261. [Google Scholar]
- Creelman, R.; Mulpuri, R. The oxylipin pathway in. Arabidopsis. Arabidopsis Book 2002. [Google Scholar] [CrossRef]
- Pinot, F.; Beisson, F. Cytochrome P450 metabolizing fatty acids in plants: Characterization and physiological roles. FEBS J 2011, 278, 195–205. [Google Scholar]
- Kuroda, H.; Oshima, T.; Kaneda, H.; Takashio, M. Identification and functional analyses of two cDNAs that encode fatty acid 9-/13-hydroperoxide lyase (CYP74C) in rice. Biosci. Biotechnol. Biochem 2005, 69, 1545–1554. [Google Scholar]
- Matsui, K.; Minami, A.; Hornung, E.; Shibata, H.; Kishimoto, K.; Ahnert, V.; Kindl, H.; Kajiwara, T.; Feussner, I. Biosynthesis of fatty acid derived aldehydes is induced upon mechanical wounding and its products show fungicidal activities in cucumber. Phytochemistry 2006, 67, 649–657. [Google Scholar]
- Tijet, N.; Schneider, C.; Muller, B.L.; Brash, A.R. Biogenesis of volatile aldehydes from fatty acid hydroperoxides: Molecular cloning of a hydroperoxide lyase (CYP74C) with specificity for both the 9- and 13-hydroperoxides of linoleic and linolenic acids. Arch. Biochem. Biophys 2001, 386, 281–289. [Google Scholar]
- Noordermeer, M.A.; Veldink, G.A.; Vliegenthart, J.F. Alfalfa contains substantial 9-hydroperoxide lyase activity and a 3Z:2E-enal isomerase. FEBS Lett 1999, 443, 201–204. [Google Scholar]
- Fauconnier, M.; Perez, A. Purification and characterization of tomato leaf (Lycopersicon esculentum Mill.) hydroperoxide lyase. J. Agric. Food Chem 1997, 8561, 4232–4236. [Google Scholar]
- Itoh, A.; Vick, B.A. The purification and characterization of fatty acid hydroperoxide lyase in sunflower. Biochim. Biophys. Acta 1999, 1436, 531–540. [Google Scholar]
- Bate, N.J.; Sivasankar, S.; Moxon, C.; Riley, J.M.; Thompson, J.E.; Rothstein, S.J. Molecular characterization of an Arabidopsis gene encoding hydroperoxide lyase, a cytochrome P-450 that is wound inducible. Plant Physiol 1998, 117, 1393–1400. [Google Scholar]
- Matsui, K.; Shibutani, M.; Hase, T.; Kajiwara, T. Bell pepper fruit fatty acid hydroperoxide lyase is a cytochrome P450 (CYP74B). FEBS Lett 1996, 394, 21–24. [Google Scholar]
- Hatanaka, A.; Kajiwara, T.; Sekiya, J. Biosynthetic pathway For C6-aldehydes formation from linolenic acid in green leaves. Chem. Phys. Lipids 1987, 44, 341–361. [Google Scholar]
- Blee, E.; Joyard, J. Envelope membranes from spinach chloroplasts are a site of metabolism of fatty acid hydroperoxides. Plant Physiol 1996, 110, 445–454. [Google Scholar]
- Froehlich, J.; Wilkerson, C.; Keith Ray, W.; McAndrew, R.; Osteryoung, K.; Gage, D.; Phinney, B. Proteomic study of the arabidopsis t haliana chloroplastic envelope membrane utilizing alternatives to traditional two-dimensional electrophoresis. J. Proteome Res 2003, 2, 413–425. [Google Scholar]
- Froehlich, J.E.; Itoh, A.; Howe, G.A. Tomato allene oxide synthase and fatty acid hydroperoxide lyase, two cytochrome P450s involved in oxylipin metabolism, are targeted to different membranes of chloroplast envelope. Plant Physiol 2001, 125, 306–317. [Google Scholar]
- Duan, H.; Huang, M.; Palacio, K.; Schuler, M. Variations in CYP74B2 (hydroperoxide lyase) gene expression differentially affect hexenal signaling in the Columbia and Landsberg erecta ecotypes of Arabidopsis. Plant Physiol 2005, 139, 1529–1544. [Google Scholar]
- Mosblech, A.; Feussner, I.; Heilmann, I. Oxylipins: Structurally diverse metabolites from fatty acid oxidation. Plant Physiol. Biochem 2009, 47, 511–517. [Google Scholar]
- Kishimoto, K.; Matsui, K.; Ozawa, R.; Takabayashi, J. Direct fungicidal activities of C6-aldehydes are important constituents for defense responses in Arabidopsis against Botrytis cinerea. Phytochemistry 2008, 69, 2127–2132. [Google Scholar]
- Bate, N.; Riley, J. Quantitative and qualitative differences in C6- volatile production from the lipoxygenase pathway in an alcohol dehydrogenase mutant of Arabidopsis thaliana. Physiol. Plant 1998, 104, 97–104. [Google Scholar]
- Takamura, H.; Gardner, H.W. Oxygenation of (3 Z) -alkenal to (2 E) -4-hydroxy-2-alkenal in soybean seed (Glycine max L.). Biochim. Biophys. Acta 1996, 1303, 83–91. [Google Scholar]
- Chang, C.; Meyerowitz, E.M. Molecular cloning and DNA sequence of the Arabidopsis thaliana alcohol dehydrogenase gene. Proc. Natl. Acad. Sci. USA 1986, 83, 1408–1412. [Google Scholar]
- Strommer, J. The plant ADH gene family. Plant J 2011, 66, 128–142. [Google Scholar]
- Dolferus, R.; van Den Bossche, D.; Jacobs, M. Sequence analysis of two null-mutant alleles of the single Arabidopsis Adh locus. Mol. Gen. Genet 1990, 224, 297–302. [Google Scholar]
- D’Auria, J.C. Acyltransferases in plants: A good time to be BAHD. Curr. Opin. Plant Biol 2006, 9, 331–340. [Google Scholar]
- Pieterse, C.M.J.; Leon-Reyes, A.; van der Ent, S.; van Wees, S.C.M. Networking by small-molecule hormones in plant immunity. Nat. Chem. Biol 2009, 5, 308–316. [Google Scholar]
- Song, W.C.; Funk, C.D.; Brash, A.R. Molecular cloning of an allene oxide synthase: A cytochrome P450 specialized for the metabolism of fatty acid hydroperoxides. Proc. Natl. Acad. Sci. USA 1993, 90, 8519–8523. [Google Scholar]
- Lee, D.-S.; Nioche, P.; Hamberg, M.; Raman, C.S. Structural insights into the evolutionary paths of oxylipin biosynthetic enzymes. Nature 2008, 455, 363–368. [Google Scholar]
- Chehab, E.W.; Kaspi, R.; Savchenko, T.; Rowe, H.; Negre-Zakharov, F.; Kliebenstein, D.; Dehesh, K. Distinct roles of jasmonates and aldehydes in plant-defense responses. PLoS One 2008, 3, e1904. [Google Scholar]
- Tong, X.; Qi, J.; Zhu, X.; Mao, B.; Zeng, L.; Wang, B.; Li, Q.; Zhou, G.; Xu, X.; Lou, Y.; et al. The rice hydroperoxide lyase OsHPL3 functions in defense responses by modulating the oxylipin pathway. Plant J 2012, 71, 763–775. [Google Scholar]
- Laudert, D.; Pfannschmidt, U.; Lottspeich, F.; Holländer-Czytko, H.; Weiler, E.W. Cloning, molecular and functional characterization of Arabidopsis thaliana allene oxide synthase (CYP 74), the first enzyme of the octadecanoid pathway to jasmonates. Plant Mol. Biol 1996, 31, 323–335. [Google Scholar]
- Farmaki, T.; Sanmartín, M.; Jiménez, P.; Paneque, M.; Sanz, C.; Vancanneyt, G.; León, J.; Sánchez-Serrano, J.J. Differential distribution of the lipoxygenase pathway enzymes within potato chloroplasts. J. Exp. Bot 2007, 58, 555–568. [Google Scholar]
- Gounaris, K.; Brain, A.R.R.; Quinn, P.J.; Williams, W.P. Structural reorganization of chloroplast thylakoid membranes in response to heat stress. Biochim. Biophys. Acta 1984, 766, 198–208. [Google Scholar]
- Larkindale, J.; Huang, B. Changes of lipid composition and saturation level in leaves and roots for heat-stressed and heat-acclimated creeping bentgrass (Agrostis stolonifera). Environ. Exp. Bot 2004, 51, 57–67. [Google Scholar]
- Xu, S.; Li, J.; Zhang, X.; Wei, H.; Cui, L. Effects of heat acclimation pretreatment on changes of membrane lipid peroxidation, antioxidant metabolites, and ultrastructure of chloroplasts in two cool-season turfgrass species under heat stress. Environ. Ex. Bot 2006, 56, 274–285. [Google Scholar]
- Loreto, F.; Barta, C.; Brilli, F.; Nogues, I. On the induction of volatile organic compound emissions by plants as consequence of wounding or fluctuations of light and temperature. Plant Cell Environ 2006, 29, 1820–1828. [Google Scholar]
- Holzinger, R.; Sandoval-Soto, L.; Rottenberger, S.; Crutzen, P.; Kesselmeier, J. Emissions of volatile organic compounds from Quercus ilex L. measured by Proton Transfer Reaction Mass Spectrometry using a fast Proton Transfer Reaction Mass. J. Geophys. Res 2000, 105, 573–579. [Google Scholar]
- Graus, M.; Schnitzler, J.; Hansel, A. Transient release of oxygenated volatile organic compounds during light-dark transitions in grey poplar leaves. Plant Physiol 2004, 135, 1967–1975. [Google Scholar]
- Sharkey, T.D.; Wiberley, A.E.; Donohue, A.R. Isoprene emission from plants: why and how. Ann. Bot 2008, 101, 5–18. [Google Scholar]
- Hartikainen, K.; Riikonen, J.; Nerg, A.-M.; Kivimäenpää, M.; Ahonen, V.; Tervahauta, A.; Kärenlampi, S.; Mäenpää, M.; Rousi, M.; Kontunen-Soppela, S.; et al. Impact of elevated temperature and ozone on the emission of volatile organic compounds and gas exchange of silver birch (Betula pendula Roth). Environ. Exp. Bot 2012, 84, 33–43. [Google Scholar]
- Farmer, E.E.; Davoine, C. Reactive electrophile species. Curr. Opin. Plant Biol 2007, 10, 380–386. [Google Scholar]
- Matsui, K.; Sugimoto, K.; Mano, J.; Ozawa, R.; Takabayashi, J. Differential metabolisms of green leaf volatiles in injured and intact parts of a wounded leaf meet distinct ecophysiological requirements. PLoS One 2012, 7, e36433. [Google Scholar]
- Yan, Z.; Wang, C. Wound-induced green leaf volatiles cause the release of acetylated derivatives and a terpenoid in maize. Phytochemistry 2006, 67, 34–42. [Google Scholar]
- Mirabella, R.; Rauwerda, H.; Struys, E.A.; Jakobs, C.; Triantaphylidès, C.; Haring, M.A.; Schuurink, R.C.; Triantaphylides, C. The Arabidopsis her1 mutant implicates GABA in E-2-hexenal responsiveness. Plant J: Cell Mol. Biol 2008, 53, 197–213. [Google Scholar]
- Prost, I.; Dhondt, S.; Rothe, G. Evaluation of the antimicrobial activities of plant oxylipins supports their involvement in defense against pathogens. Plant Physiol 2005, 139, 1902–1913. [Google Scholar]
- Bate, N.J.; Rothstein, S.J. C6-volatiles derived from the lipoxygenase pathway induce a subset of defense-related genes. Plant J. Cell Mol. Biol 1998, 16, 561–569. [Google Scholar]
- Farag, M.A.; Fokar, M.; Abd, H.; Zhang, H.; Allen, R.D.; Paré, P.W. (Z)-3-Hexenol induces defense genes and downstream metabolites in maize. Planta 2005, 220, 900–909. [Google Scholar]
- Farag, M.A.; Paré, P.W. C6-Green leaf volatiles trigger local and systemic VOC emissions in tomato. Phytochemistry 2002, 61, 545–554. [Google Scholar]
- Kishimoto, K.; Matsui, K.; Ozawa, R.; Takabayashi, J. Volatile C6-aldehydes and Allo-ocimene activate defense genes and induce resistance against Botrytis cinerea in Arabidopsis thaliana. Plant Cell Physiol 2005, 46, 1093–1102. [Google Scholar]
- Kishimoto, K.; Matsui, K.; Ozawa, R.; Takabayashi, J. ETR1-, JAR1- and PAD2-dependent signaling pathways are involved in C6-aldehyde-induced defense responses of Arabidopsis. Plant Sci.: Int. J. Exp. Plant Biol 2006, 171, 415–423. [Google Scholar]
- Paschold, A.; Halitschke, R.; Baldwin, I.T. Using “mute” plants to translate volatile signals. Plant J 2006, 45, 275–291. [Google Scholar]
- Zeringue, H.J., Jr. Effects of C6-C10 alkenals and alkanals on eliciting a defence response in the developing cotton boll. Phytochemistry 1992, 31, 2305–2308. [Google Scholar]
- Engelberth, J.; Alborn, H.T.; Schmelz, E.A.; Tumlinson, J.H. Airborne signals prime plants against insect herbivore attack. Proc. Natl. Acad. Sci. USA 2004, 101, 1781–1785. [Google Scholar]
- Heil, M.; Kost, C. Priming of indirect defences. Ecol. Lett 2006, 9, 813–817. [Google Scholar]
- Fujita, M.; Hossain, M.Z. Modulation of pumpkin glutathione S-transferases by aldehydes and related compounds. Plant Cell Physiol 2003, 44, 481–490. [Google Scholar]
- Davoine, C.; Falletti, O.; Douki, T.; Iacazio, G.; Ennar, N.; Montillet, J.L.; Triantaphylides, C. Adducts of oxylipin electrophiles to glutathione reflect a 13 specificity of the downstream lipoxygenase pathway in the tobacco hypersensitive response. Plant Physiol 2006, 140, 1484–1493. [Google Scholar]
- Bouché, N.; Fromm, H. GABA in plants: Just a metabolite? Trends Plant Sci 2004, 9, 110–115. [Google Scholar]
- López, M.A.; Bannenberg, G.; Castresana, C. Controlling hormone signaling is a plant and pathogen challenge for growth and survival. Curr. Opin. Plant Biol 2008, 11, 420–427. [Google Scholar]
- Pieterse, C.M.J.; van der Does, D.; Zamioudis, C.; Leon-Reyes, A.; van Wees, S.C.M. Hormonal modulation of plant immunity. Annu. Rev. Cell Dev. Biol 2012, 28, 489–521. [Google Scholar]
- Glazebrook, J. Contrasting mechanisms of defense against biotrophic and necrotrophic pathogens. Annu. Rev. Phytopathol 2005, 43, 205–227. [Google Scholar]
- Thomma, B.; Penninckx, I.; Broekaert, W.F. The complexity of disease signaling in Arabidopsis. Curr. Opin. Immunol. 2001, 13, 63–68. [Google Scholar]
- Koo, A.; Howe, G. The wound hormone jasmonate. Phytochemistry 2009, 70, 1571–1580. [Google Scholar]
- Kant, M.R.; Sabelis, M.W.; Haring, M.A.; Schuurink, R.C. Intraspecific variation in a generalist herbivore accounts for differential induction and impact of host plant defences. Proc. Biol. Sci. R. Soc 2008, 275, 443–452. [Google Scholar]
- Doherty, H.; Selvendran, R.; Bowles, D. The wound response of tomato plants can be inhibited by aspirin and related hydroxy-benzoic acids. Physiol. Mol. Plant Pathol 1988, 33, 377–384. [Google Scholar]
- Penninckx, I.A.; Eggermont, K.; Terras, F.R.; Thomma, B.P.; de Samblanx, G.W.; Buchala, A.; Métraux, J.P.; Manners, J.M.; Broekaert, W.F. Pathogen-induced systemic activation of a plant defensin gene in Arabidopsis follows a salicylic acid-independent pathway. Plant Cell 1996, 8, 2309–2323. [Google Scholar]
- Spoel, S.; Koornneef, A. NPR1 modulates cross-talk between salicylate-and jasmonate-dependent defense pathways through a novel function in the cytosol. Plant Cell 2003, 15, 760–770. [Google Scholar]
- Koornneef, A.; Leon-Reyes, A.; Ritsema, T.; Verhage, A.; den Otter, F.C.; van Loon, L.C.; Pieterse, C.M.J. Kinetics of salicylate-mediated suppression of jasmonate signaling reveal a role for redox modulation. Plant Physiol 2008, 147, 1358–1368. [Google Scholar]
- Penninckx, I.A.; Thomma, B.P.; Buchala, A.; Métraux, J.P.; Broekaert, W.F. Concomitant activation of jasmonate and ethylene response pathways is required for induction of a plant defensin gene in Arabidopsis. Plant Cell 1998, 10, 2103–2113. [Google Scholar]
- Van Wees, S.C.; de Swart, E.A.; van Pelt, J.A.; van Loon, L.C.; Pieterse, C.M. Enhancement of induced disease resistance by simultaneous activation of salicylate- and jasmonate-dependent defense pathways in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2000, 97, 8711–8716. [Google Scholar]
- Schenk, P.M.; Kazan, K.; Wilson, I.; Anderson, J.P.; Richmond, T.; Somerville, S.C.; Manners, J.M. Coordinated plant defense responses in Arabidopsis revealed by microarray analysis. Proc. Natl. Acad. Sci. USA 2000, 97, 11655–11660. [Google Scholar]
- Lawton, K.A.; Potter, S.L.; Uknes, S.; Ryals, J. Acquired resistance signal transduction in arabidopsis is ethylene independent. Plant Cell 1994, 6, 581–588. [Google Scholar]
- De Vos, M.; van Zaanen, W.; Koornneef, A.; Korzelius, J.P.; Dicke, M.; van Loon, L.C.; Pieterse, C.M.J. Herbivore-induced resistance against microbial pathogens in Arabidopsis. Plant Physiol 2006, 142, 352–363. [Google Scholar]
- Mur, L.; Kenton, P.; Atzorn, R. Outcomes of concentration-specific interactions between salicylate and jasmonate signaling include synergy, antagonism, and oxidative stress leading to cell death. Plant Physiol 2006, 140, 249–262. [Google Scholar]
- Pré, M.; Atallah, M.; Champion, A.; de Vos, M.; Pieterse, C.M.J.; Memelink, J. The AP2/ERF domain transcription factor ORA59 integrates jasmonic acid and ethylene signals in plant defense. Plant Physiol 2008, 147, 1347–1357. [Google Scholar]
- Ruther, J.; Kleier, S. Plant-plant signaling: Ethylene synergizes volatile emission in Zea mays induced by exposure to (Z)-3-hexen-1-ol. J. Chem. Ecol 2005, 31, 2217–2222. [Google Scholar]
- Arimura, G.; Ozawa, R.; Horiuchi, J.; Nishioka, T.; Takabayashi, J. Plant–plant interactions mediated by volatiles emitted from plants infested by spider mites. Biochem. Syst. Ecol 2001, 29, 1049–1061. [Google Scholar]
- Whalen, M.C.; Innes, R.W.; Bent, A.F.; Staskawicz, B.J. Identification of Pseudomonas syringae pathogens of Arabidopsis and a bacterial locus determining avirulence on both Arabidopsis and soybean. Plant Cell 1991, 3, 49–59. [Google Scholar]
- Rico, A.; Preston, G.M. Pseudomonas syringae pv. tomato DC3000 uses constitutive and apoplast-induced nutrient assimilation pathways to catabolize nutrients that are abundant in the tomato apoplast. Mol. Plant Microbe Interact 2008, 21, 269–282. [Google Scholar]
- Hamilton-Kemp, T. Effects of some natural volatile compounds on the pathogenic fungiAlternaria alternata andBotrytis cinerea. J. Chem. Ecol 1992, 18, 1083–1091. [Google Scholar]
- Nakamura, S.; Hatanaka, A. Green-leaf-derived C6-aroma compounds with potent antibacterial action that act on both Gram-negative and Gram-positive bacteria. J. Agric. Food 2002, 50, 7639–7644. [Google Scholar]
- Hirano, S.S.; Upper, C.D. Bacteria in the leaf ecosystem with emphasis on Pseudomonas syringae-a pathogen, ice nucleus, and epiphyte. Microbiol. Mol. Biol. Rev 2000, 64, 624–653. [Google Scholar]
- Block, A.; Schmelz, E.; Jones, J.B.; Klee, H.J. Coronatine and salicylic acid: The battle between Arabidopsis and Pseudomonas for phytohormone control. Mol. Plant Pathol 2005, 6, 79–83. [Google Scholar]
- Brooks, D.M.D.; Bender, C.L.C.; Kunkel, B.N.B. The Pseudomonas syringae phytotoxin coronatine promotes virulence by overcoming salicylic acid-dependent defences in Arabidopsis thaliana. Mol. Plant Pathol 2005, 6, 629–639. [Google Scholar]
- Mei, C.; Qi, M.; Sheng, G.; Yang, Y. Inducible overexpression of a rice allene oxide synthase gene increases the endogenous jasmonic acid level, PR gene expression, and host resistance to fungal infection. Mol. Plant Microbe Interact. MPMI 2006, 19, 1127–1137. [Google Scholar]
- Yuan, Y.; Zhong, S.; Li, Q.; Zhu, Z.; Lou, Y.; Wang, L.; Wang, J.; Wang, M.; Li, Q.; Yang, D.; et al. Functional analysis of rice NPR1-like genes reveals that OsNPR1/NH1 is the rice orthologue conferring disease resistance with enhanced herbivore susceptibility. Plant Biotechnol. J 2007, 5, 313–324. [Google Scholar]
- Yang, D.-L.; Li, Q.; Deng, Y.-W.; Lou, Y.-G.; Wang, M.-Y.; Zhou, G.-X.; Zhang, Y.-Y.; He, Z.-H. Altered disease development in the eui mutants and Eui overexpressors indicates that gibberellins negatively regulate rice basal disease resistance. Mol. Plant 2008, 1, 528–537. [Google Scholar]
- Piesik, D.; Lemńczyk, G.; Skoczek, A.; Lamparski, R.; Bocianowski, J.; Kotwica, K.; Delaney, K.J. Fusarium infection in maize: volatile induction of infected and neighboring uninfected plants has the potential to attract a pest cereal leaf beetle, Oulema melanopus. J. Plant Physiol 2011, 168, 1534–1542. [Google Scholar]
- Ongena, M.; Duby, F.; Rossignol, F.; Fauconnier, M.-L.; Dommes, J.; Thonart, P. Stimulation of the lipoxygenase pathway is associated with systemic resistance induced in bean by a nonpathogenic Pseudomonas strain. Mol. Plant Microbe Interact 2004, 17, 1009–1018. [Google Scholar]
- Major, T.; Marchini, P.; Sproston, T. Isolation from Ginkgo of Fungus biloba L. of an Inhibitor Growth. J. Biol. Chem 1960, 235, 3298–3299. [Google Scholar]
- Zeringue, H.J.; McCormick, S.P. Relationships between cotton leaf-derived volatiles and growth of aspergiflus flavus. JAOCS 1989, 66, 581–585. [Google Scholar]
- Zeringue, H.J.; Brown, R.L.; Neucere, J.N.; Cleveland, T.E. Relationships between C6–C12 alkanal and alkenal volatile contents and resistance of maize genotypes to Aspergillus flavus and aflatoxin production. J. Agric. Food Chem 1996, 44, 403–407. [Google Scholar]
- Baker, A.G.; Wiesler, D.; Novotny, M.V. Tandem mass spectrometry of model peptides modified with trans-2-hexenal, a product of. J. Am. Soc. Mass. Spectrom 1999, 10, 613–624. [Google Scholar]
- Myung, K. Interaction with and effects on the profile of proteins of Botrytis cinerea by C6 aldehydes. J. Agric 2007, 55, 2182–2188. [Google Scholar]
- Kishimoto, K.; Matsui, K.; Ozawa, R.; Takabayashi, J. Components of C6-aldehyde-induced resistance in Arabidopsis thaliana against a necrotrophic fungal pathogen, Botrytis cinerea. Plant Sci 2006, 170, 715–723. [Google Scholar]
- Glawischnig, E. Camalexin. Phytochemistry 2007, 68, 401–406. [Google Scholar]
- Ferrari, S.; Plotnikova, J.M.; de Lorenzo, G.; Ausubel, F.M. Arabidopsis local resistance to Botrytis cinerea involves salicylic acid and camalexin and requires EDS4 and PAD2, but not SID2, EDS5 or PAD4. Plant J 2003, 35, 193–205. [Google Scholar]
- Devoto, A.; Ellis, C.; Magusin, A.; Chang, H.-S.; Chilcott, C.; Zhu, T.; Turner, J.G. Expression profiling reveals COI1 to be a key regulator of genes involved in wound- and methyl jasmonate-induced secondary metabolism, defence, and hormone interactions. Plant Mol. Biol 2005, 58, 497–513. [Google Scholar]
- Bell, E.; Creelman, R.A.; Mullet, J.E. A chloroplast lipoxygenase is required for wound-induced jasmonic acid accumulation in Arabidopsis. Proc. Natl. Acad. Sci. USA 1995, 92, 8675–8679. [Google Scholar]
- Thomma, B.P.; Eggermont, K.; Penninckx, I.A.; Mauch-Mani, B.; Vogelsang, R.; Cammue, B.P.; Broekaert, W.F. Separate jasmonate-dependent and salicylate-dependent defense-response pathways in Arabidopsis are essential for resistance to distinct microbial pathogens. Proc. Natl. Acad. Sci. USA 1998, 95, 15107–15111. [Google Scholar]
- Ton, J.; van Pelt, J.A.; van Loon, L.C.; Pieterse, C.M.J. Differential effectiveness of salicylate-dependent and jasmonate/ethylene-dependent induced resistance in Arabidopsis. Mol. Plant Microbe Interact 2002, 15, 27–34. [Google Scholar]
- Pieterse, C.; Ton, J.; Loon, L. Van Cross-talk between plant defence signalling pathways: Boost or burden? AgBiotechNet 2001, 3, 1–8. [Google Scholar]
- Yedidia, I.; Shoresh, M.; Kerem, Z.; Kapulnik, Y.; Chet, I.; Benhamou, N. Concomitant induction of systemic resistance to pseudomonas syringae pv. lachrymans in cucumber by trichoderma asperellum (T-203) and accumulation of phytoalexins concomitant induction of systemic resistance to pseudomonas syringae pv. lachrymans in C. Appl. Environ. Microbiol 2003, 69, 7343–7353. [Google Scholar]
- McMenemy, L.S.; Hartley, S.E.; MacFarlane, S.A.; Karley, A.J.; Shepherd, T.; Johnson, S.N. Raspberry viruses manipulate the behaviour of their insect vectors. Entomol. Exp. Appl 2012, 144, 56–68. [Google Scholar]
- Dicke, M.; van Loon, J.J.A.; Soler, R. Chemical complexity of volatiles from plants induced by multiple attack. Nat. Chem. Biol 2009, 5, 317–324. [Google Scholar]
- Mumm, R.; Dicke, M. Variation in natural plant products and the attraction of bodyguards involved in indirect plant defenseThe present review is one in the special series of reviews on animal–plant interactions. Can. J. Zool 2010, 88, 628–667. [Google Scholar]
- Dicke, M.; Sabelis, M. Infochemical terminology: Based on cost-benefit analysis rather than origin of compounds? Funct. Ecol 1988, 2, 131–139. [Google Scholar]
- Heil, M.; Ton, J. Long-distance signalling in plant defence. Trends Plant Sci 2008, 13, 264–272. [Google Scholar]
- James, D.G. Further field evaluation of synthetic herbivore-induced plan volatiles as attractants for beneficial insects. J. Chem. Ecol 2005, 31, 481–495. [Google Scholar]
- Shiojiri, K.; Ozawa, R.; Matsui, K.; Kishimoto, K.; Kugimiya, S.; Takabayashi, J. Role of the lipoxygenase/lyase pathway of host-food plants in the host searching behavior of two parasitoid species, Cotesia glomerata and Cotesia plutellae. J. Chem. Ecol 2006, 32, 969–979. [Google Scholar]
- Shimoda, T. A key volatile infochemical that elicits a strong olfactory response of the predatory mite Neoseiulus californicus, an important natural enemy of the two-spotted spider mite Tetranychus urticae. Exp.Appl. Acarol 2010, 50, 9–22. [Google Scholar]
- Halitschke, R.; Stenberg, J.A.; Kessler, D.; Kessler, A.; Baldwin, I.T. Shared signals - “alarm calls” from plants increase apparency to herbivores and their enemies in nature. Ecol. Lett 2008, 11, 24–34. [Google Scholar]
- Schuman, M.C.; Barthel, K.; Baldwin, I.T. Herbivory-induced volatiles function as defenses increasing fitness of the native plant Nicotiana attenuata in nature. eLife 2012, 1, e00007. [Google Scholar]
- Dicke, M.; van Loon, J.J.A. Multitrophic effects of herbivore-induced plant volatiles in an evolutionary context. Entomol. Exp. et Appl 2000, 97, 237–249. [Google Scholar]
- Dicke, M.; Baldwin, I.T. The evolutionary context for herbivore-induced plant volatiles: Beyond the “cry for help”. Trends Plant Sci 2010, 15, 167–175. [Google Scholar]
- Meldau, S.; Wu, J.Q.; Baldwin, I.T. Silencing two herbivory-activated MAP kinases, SIPK and WIPK, does not increase Nicotiana attenuata’s susceptibility to herbivores in the glasshouse and in nature. New Phytol 2009, 181, 161–173. [Google Scholar]
- Han, B.; Zhang, Q.-H.; Byers, J.A. Attraction of the tea aphid, Toxoptera aurantii, to combinations of volatiles and colors related to tea plants. Entomol. Exp. Appl 2012, 144, 258–269. [Google Scholar]
- Reinecke, A.; Ruther, J.; Tolasch, T.; Francke, W.; Hilker, M. Alcoholism in cockchafers: Orientation of male Melolontha melolontha towards green leaf alcohols. Naturwissenschaften 2002, 89, 265–269. [Google Scholar]
- Ruther, J.; Reinecke, A.; Hilker, M. Plant volatiles in the sexual communication of Melolontha hippocastani: response towards time-dependent bouquets and novel function of (Z)-3-hexen-1-ol as a sexual kairomone. Ecol. Entomol 2002, 27, 76–83. [Google Scholar]
- Ruther, J. Male-biassed response of garden chafer, Phyllopertha horticola L., to leaf alcohol and attraction of both sexes to floral plant volatiles. Chemoecology 2004, 14, 187–192. [Google Scholar]
- Reddy, G.V.P.; Guerrero, A. Interactions of insect pheromones and plant semiochemicals. Trends Plant Sci 2004, 9, 253–261. [Google Scholar]
- Hildebrand, D.F.; Brown, G.C.; Jackson, D.M.; Hamilton-Kemp, T.R. Effects of some leaf-emitted volatile compounds on aphid population increase. J. Chem. Ecol 1993, 19, 1875–1887. [Google Scholar]
- Vancanneyt, G.; Sanz, C.; Farmaki, T.; Paneque, M.; Ortego, F.; Castañera, P.; Sánchez-Serrano, J.J. Hydroperoxide lyase depletion in transgenic potato plants leads to an increase in aphid performance. Proc. Natl. Acad. Sci. USA 2001, 98, 8139–8144. [Google Scholar]
- Visser, J.; Avé, D. General green leaf volatiles in the olfactory orientation of the Colorado beetle, Leptinotarsa decemlineata. Entomol. Exper. Appl 1978, 24, 738–749. [Google Scholar]
- Allmann, S.; Späthe, A.; Bisch-Knaden, S.; Kallenbach, M.; Reinecke, A.; Sachse, S.; Baldwin, I.T.; Hansson, B.S. Feeding-induced rearrangement of green leaf volatiles reduces moth oviposition. ELife 2013. [Google Scholar] [CrossRef]
- Sánchez-Hernández, C.; López, M.G.; Delano-Frier, J.P. Reduced levels of volatile emissions in jasmonate-deficient spr2 tomato mutants favour oviposition by insect herbivores. Plant Cell Environ 2006, 29, 546–557. [Google Scholar]
- Bruce, T.J.A.; Wadhams, L.J.; Woodcock, C.M. Insect host location: a volatile situation. Trends Plant Sci 2005, 10, 269–274. [Google Scholar]
- Furuichi, T.; Mori, I.C.; Takahashi, K.; Muto, S. Sugar-induced increase in cytosolic Ca(2+) in Arabidopsis thaliana whole plants. Plant Cell Physiol 2001, 42, 1149–1155. [Google Scholar]
- Asai, N.; Nishioka, T. Plant volatiles regulate the activities of Ca2+-permeable channels and promote cytoplasmic calcium transients in Arabidopsis leaf cells. Plant Signal. Behav 2009, 4, 294–300. [Google Scholar]
- Mori, I.C.; Schroeder, J.I. Reactive oxygen species activation of plant Ca2+ channels. A signaling mechanism in polar growth, hormone transduction, stress signaling, and hypothetically mechanotransduction 1. Plant Physiol 2004, 135, 702–708. [Google Scholar]
- Zebelo, S.A.; Matsui, K.; Ozawa, R.; Maffei, M.E. Plasma membrane potential depolarization and cytosolic calcium flux are early events involved in tomato (Solanum lycopersicon) plant-to-plant communication. Plant Sci 2012, 196, 93–100. [Google Scholar]
- Shiojiri, K.; Ozawa, R.; Matsui, K.; Sabelis, M.W.; Takabayashi, J. Intermittent exposure to traces of green leaf volatiles triggers a plant response. Sci. Rep. 2012, 2. [Google Scholar] [CrossRef]
- Arimura, G.-I.; Matsui, K.; Takabayashi, J. Chemical and molecular ecology of herbivore-induced plant volatiles: proximate factors and their ultimate functions. Plant Cell Physiol 2009, 50, 911–923. [Google Scholar]
- Heil, M.; Adame-Álvarez, R.M. Short signalling distances make plant communication a soliloquy. Biol. Lett 2010, 6, 843–845. [Google Scholar]
- Choh, Y.; Kugimiya, S.; Takabayashi, J. Induced production of extrafloral nectar in intact lima bean plants in response to volatiles from spider mite-infested conspecific plants as a possible indirect defense against spider mites. Oecologia 2006, 147, 455–460. [Google Scholar]
- Heil, M.; Silva Bueno, J.C. Within-plant signaling by volatiles leads to induction and priming of an indirect plant defense in nature. Proc. Natl. Acad. Sci. USA 2007, 104, 5467–5472. [Google Scholar]
- Kost, C.; Heil, M. Herbivore-induced plant volatiles induce an indirect defence in neighbouring plants. J. Ecol 2006, 94, 619–628. [Google Scholar]
- Frost, C.J.; Appel, H.M.; Carlson, J.E.; de Moraes, C.M.; Mescher, M.C.; Schultz, J.C. Within-plant signalling via volatiles overcomes vascular constraints on systemic signalling and primes responses against herbivores. Ecol. Lett 2007, 10, 490–498. [Google Scholar]
- Rodriguez-Saona, C.R.; Rodriguez-Saona, L.E.; Frost, C.J. Herbivore-induced volatiles in the perennial shrub, Vaccinium corymbosum, and their role in inter-branch signaling. J. Chem. Ecol 2009, 35, 163–175. [Google Scholar]
- Karban, R.; Maron, J.; Felton, G.; Ervin, G.; Eichenseer, H. Herbivore damage to sagebrush induces resistance in wild tobacco: evidence for eavesdropping between plants. Oikos 2003, 2, 325–332. [Google Scholar]
- Zimmerli, L.; Jakab, G.; Metraux, J.P.; Mauch-Mani, B. Potentiation of pathogen-specific defense mechanisms in Arabidopsis by beta -aminobutyric acid. Proc. Natl. Acad. Sci. USA 2000, 97, 12920–12925. [Google Scholar]
- Zimmerli, L.; Métraux, J.P.; Mauch-Mani, B. Beta-Aminobutyric acid-induced protection of Arabidopsis against the necrotrophic fungus Botrytis cinerea. Plant Physiol 2001, 126, 517–23. [Google Scholar]
- Yi, H.-S.; Heil, M.; Adame-Alvarez, R.M.; Ballhorn, D.J.; Ryu, C.-M. Airborne induction and priming of plant defenses against a bacterial pathogen. Plant Physiol 2009, 151, 2152–2161. [Google Scholar]
- Conrath, U.; Beckers, G.J.M.; Flors, V.; García-Agustín, P.; Jakab, G.; Mauch, F.; Newman, M.-A.; Pieterse, C.M.J.; Poinssot, B.; Pozo, M.J.; et al. Priming: getting ready for battle. Mol. Plan Microbe Interact 2006, 19, 1062–1071. [Google Scholar]
- Ton, J.; Mauch-Mani, B. Beta-amino-butyric acid-induced resistance against necrotrophic pathogens is based on ABA-dependent priming for callose. Plant J 2004, 38, 119–130. [Google Scholar]
- Hien Dao, T.T.; Puig, R.C.; Kim, H.K.; Erkelens, C.; Lefeber, A.W.M.; Linthorst, H.J.M.; Choi, Y.H.; Verpoorte, R. Effect of benzothiadiazole on the metabolome of Arabidopsis thaliana. Plant Physiol. Biochem 2009, 47, 146–152. [Google Scholar]
- Ross, F. Systemic acquired virus resistance infections induced by localized in plants. Virology 1961, 358, 340–358. [Google Scholar]
- Van Loon, L.C.; Bakker, P.A.H.M.; Pieterse, C.M.J. Systemic resistance induced by rhizosphere bacteria. Annu. Rev. Phytopathol 1998, 36, 453–483. [Google Scholar]
- Conrath, U.; Pieterse, C.M.J.; Mauch-Mani, B. Priming in plant-pathogen interactions. Trends Plant Sci 2002, 7, 210–216. [Google Scholar]
- Babikova, Z.; Gilbert, L.; Bruce, T.J.A.; Birkett, M.; Caulfield, J.C.; Woodcock, C.; Pickett, J.A.; Johnson, D. Underground signals carried through common mycelial networks warn neighbouring plants of aphid attack. Ecol. Lett 2013, 16, 835–843. [Google Scholar]
- Rhoades, D.F. Responses of Alder and Willow to Attack by Tent Caterpillars and Webworms: Evidence for Pheromonal Sensitivity of Willows. In Plant Resistance to Insects; Hedin, P.A., Ed.; American Chemical Society: Washington, DC, USA, 1983; Volume 208, pp. 4–55. [Google Scholar]
- Baldwin, I.T.; Schultz, J.C. Rapid changes in tree leaf chemistry induced by damage: Evidence for communication between plants. Science 1983, 221, 277–279. [Google Scholar]
- Karban, R.; Baldwin, I.T.; Baxter, K.J.; Laue, G.; Felton, G.W. Communication between plants: Induced resistance in wild tobacco plants following clipping of neighboring sagebrush. Oecologia 2000, 125, 66–71. [Google Scholar]
- Kessler, A.; Halitschke, R.; Diezel, C.; Baldwin, I.T. Priming of plant defense responses in nature by airborne signaling between Artemisia tridentata and Nicotiana attenuata. Oecologia 2006, 148, 280–292. [Google Scholar]
- Schütz, S.; Weißbecker, B.; Klein, A.; Hummel, H. Host plant selection of the Colorado potato beetle as influenced by damage induced volatiles of the potato plant. Naturwissenschaften 1997, 217, 212–217. [Google Scholar]
- Savchenko, T.; Pearse, I.S.; Ignatia, L.; Karban, R.; Dehesh, K. Insect herbivores selectively suppress the HPL branch of the oxylipin pathway in host plants. Plant J. Cell Mol. Biol 2013, 73, 653–662. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Scala, A.; Allmann, S.; Mirabella, R.; Haring, M.A.; Schuurink, R.C. Green Leaf Volatiles: A Plant’s Multifunctional Weapon against Herbivores and Pathogens. Int. J. Mol. Sci. 2013, 14, 17781-17811. https://doi.org/10.3390/ijms140917781
Scala A, Allmann S, Mirabella R, Haring MA, Schuurink RC. Green Leaf Volatiles: A Plant’s Multifunctional Weapon against Herbivores and Pathogens. International Journal of Molecular Sciences. 2013; 14(9):17781-17811. https://doi.org/10.3390/ijms140917781
Chicago/Turabian StyleScala, Alessandra, Silke Allmann, Rossana Mirabella, Michel A. Haring, and Robert C. Schuurink. 2013. "Green Leaf Volatiles: A Plant’s Multifunctional Weapon against Herbivores and Pathogens" International Journal of Molecular Sciences 14, no. 9: 17781-17811. https://doi.org/10.3390/ijms140917781