Differential Polarization Nonlinear Optical Microscopy with Adaptive Optics Controlled Multiplexed Beams

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
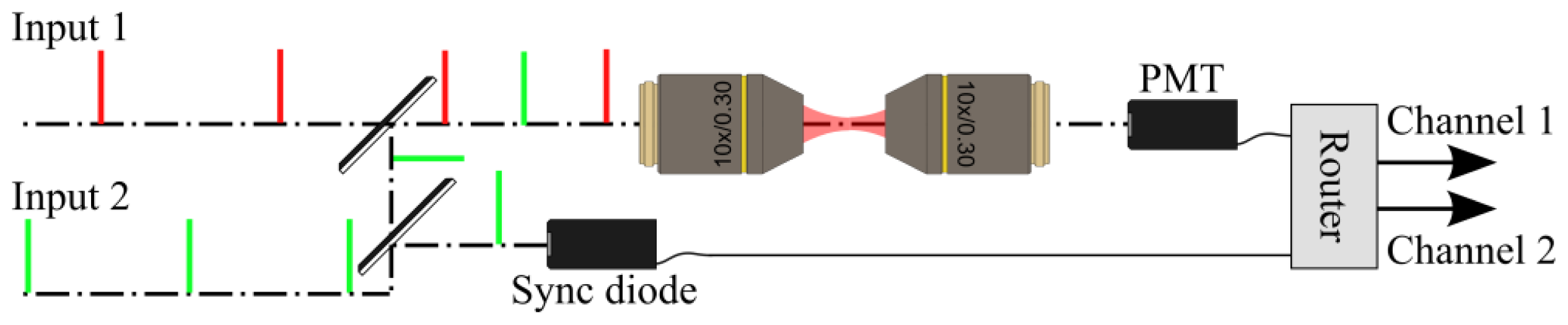
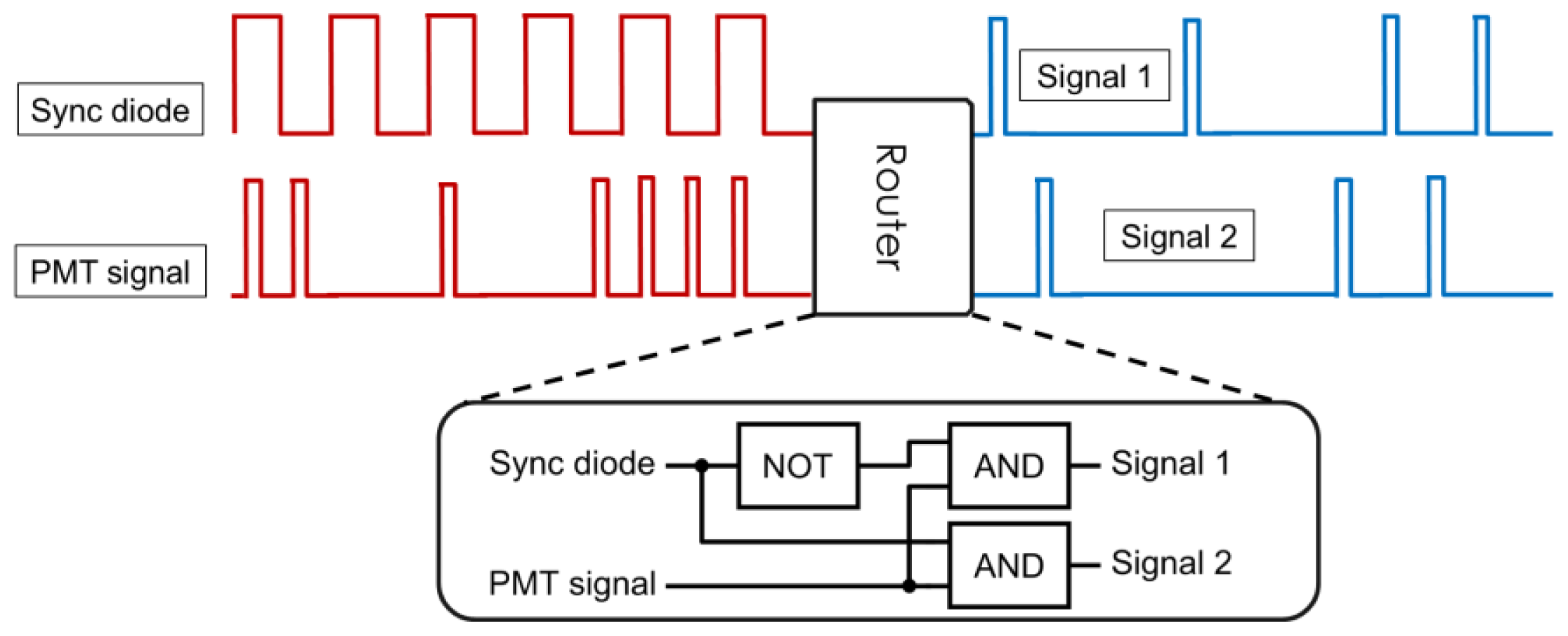
2. Differential Microscopy Setups
2.1. Principles of Differential Microscopy
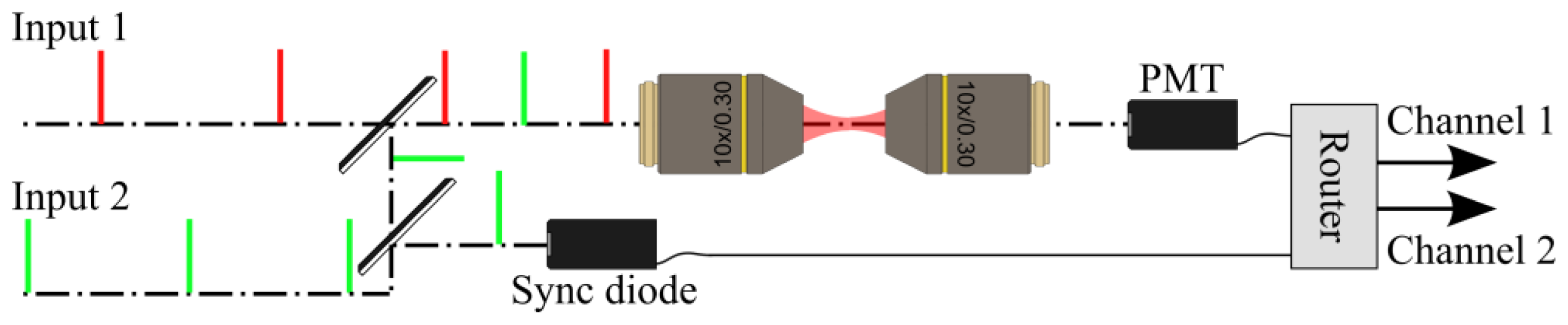
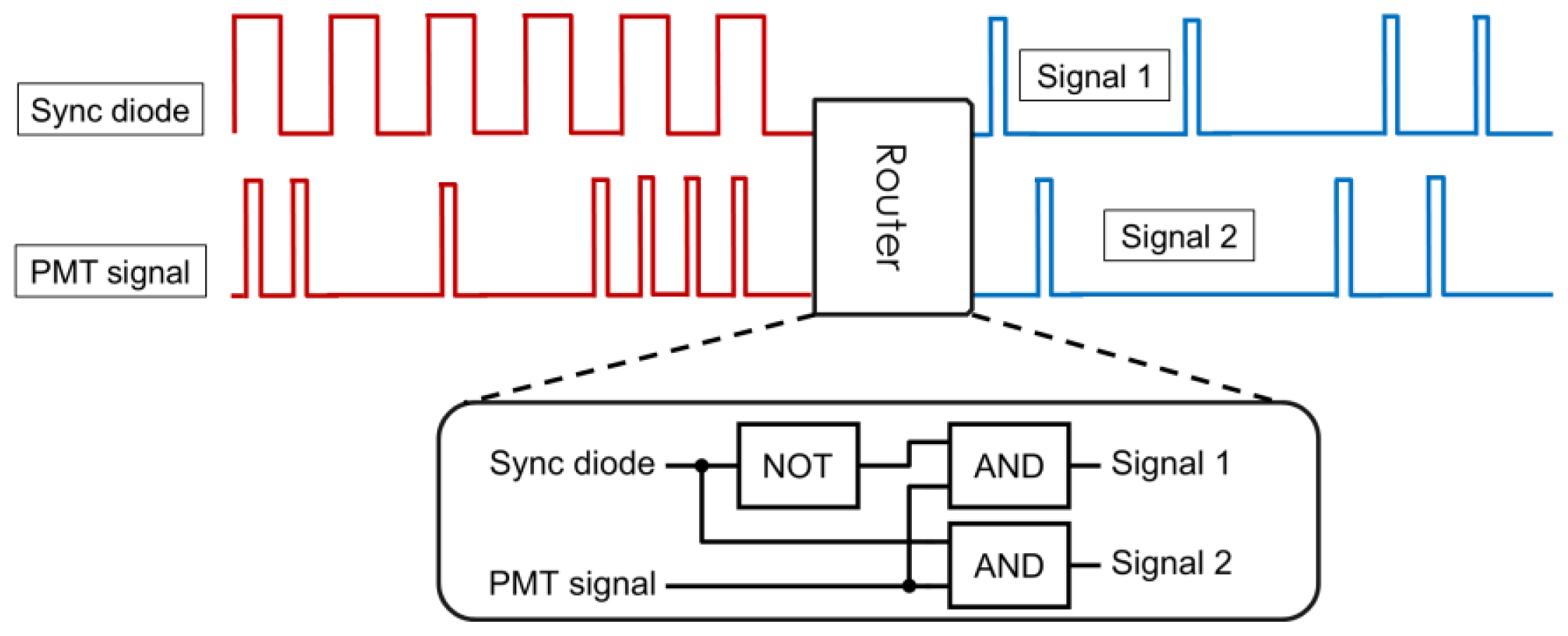
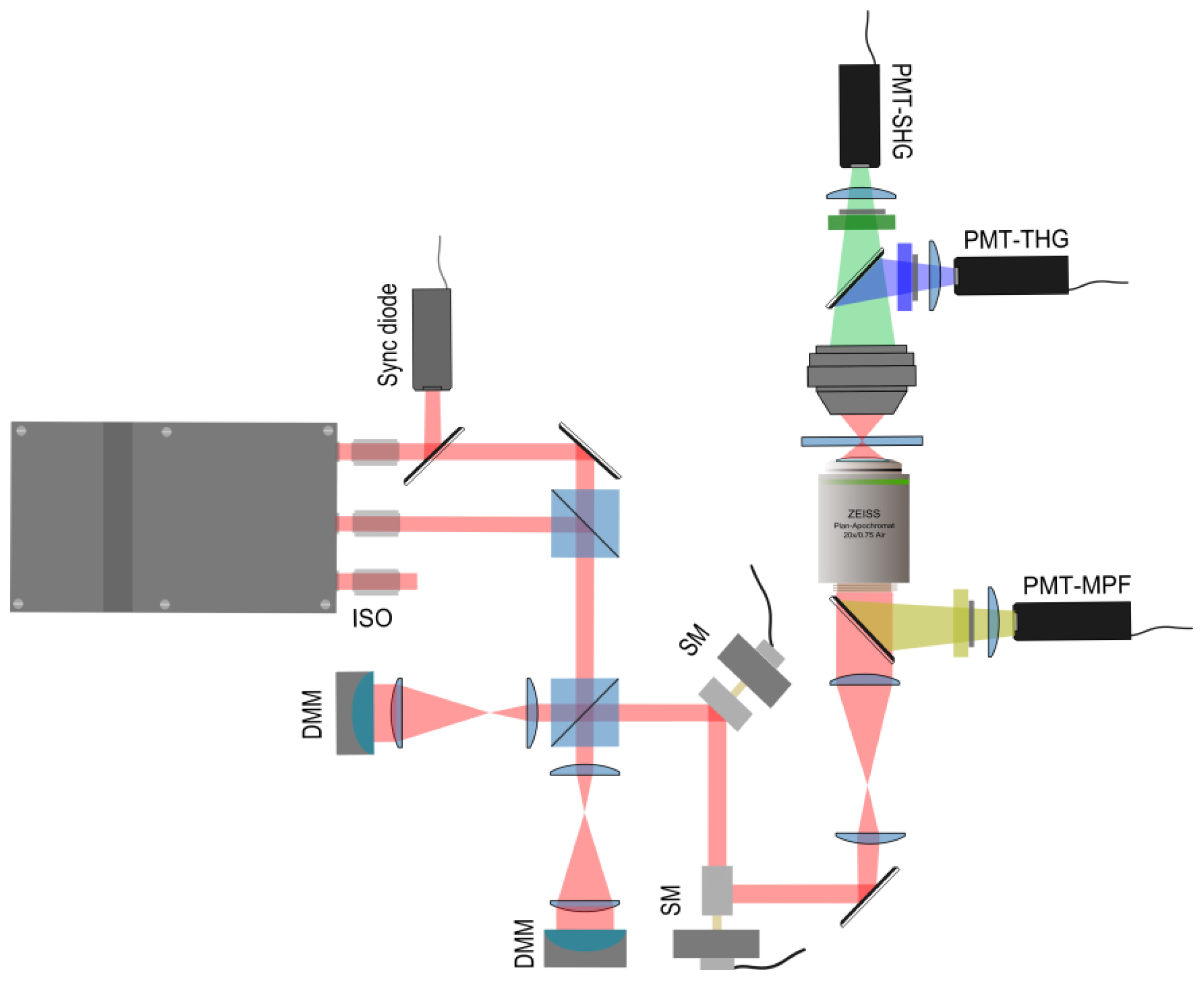
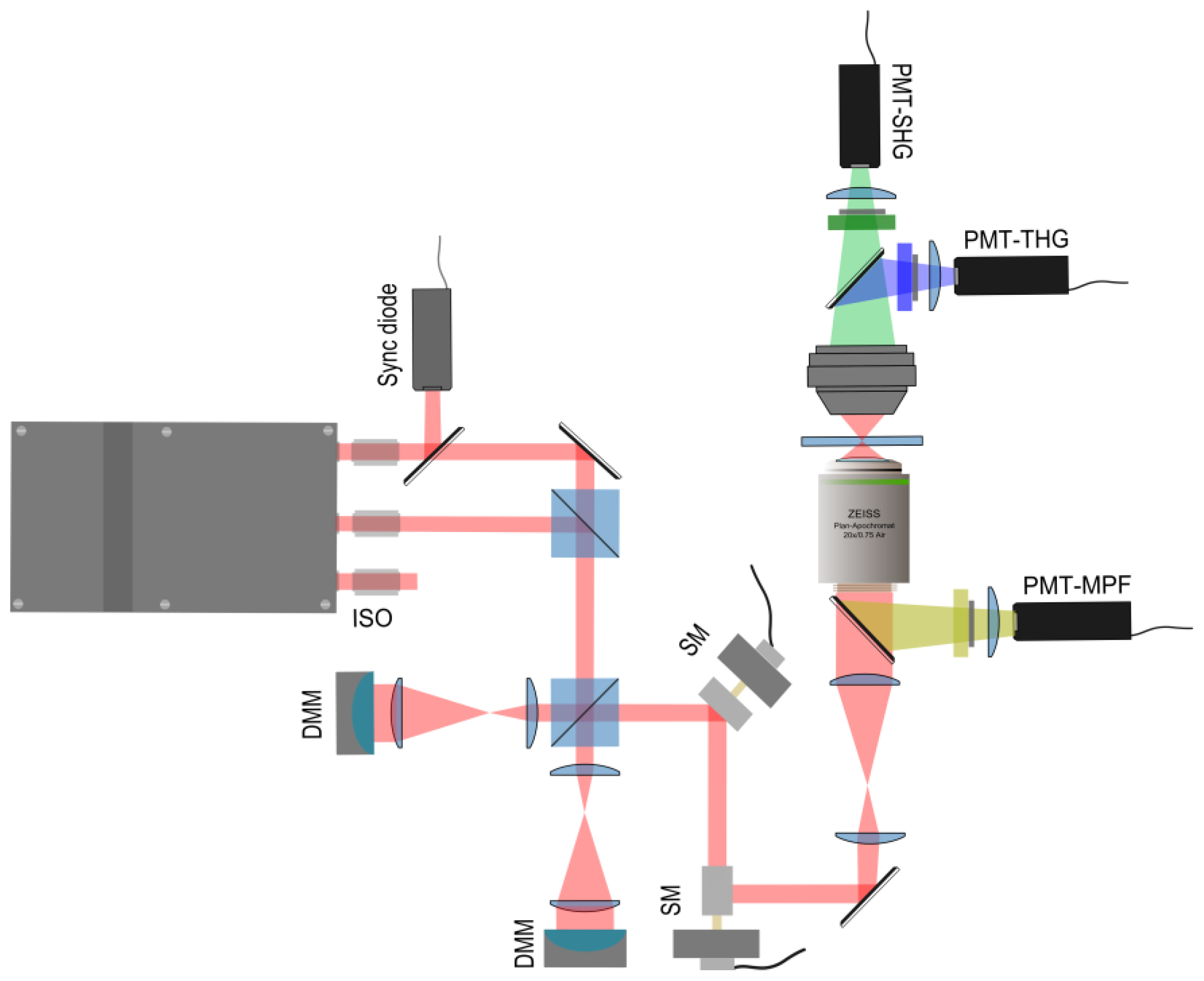
2.2. Implementation of the Differential Polarization Microscope
2.3. Adaptive Optics for Differential Microscopy
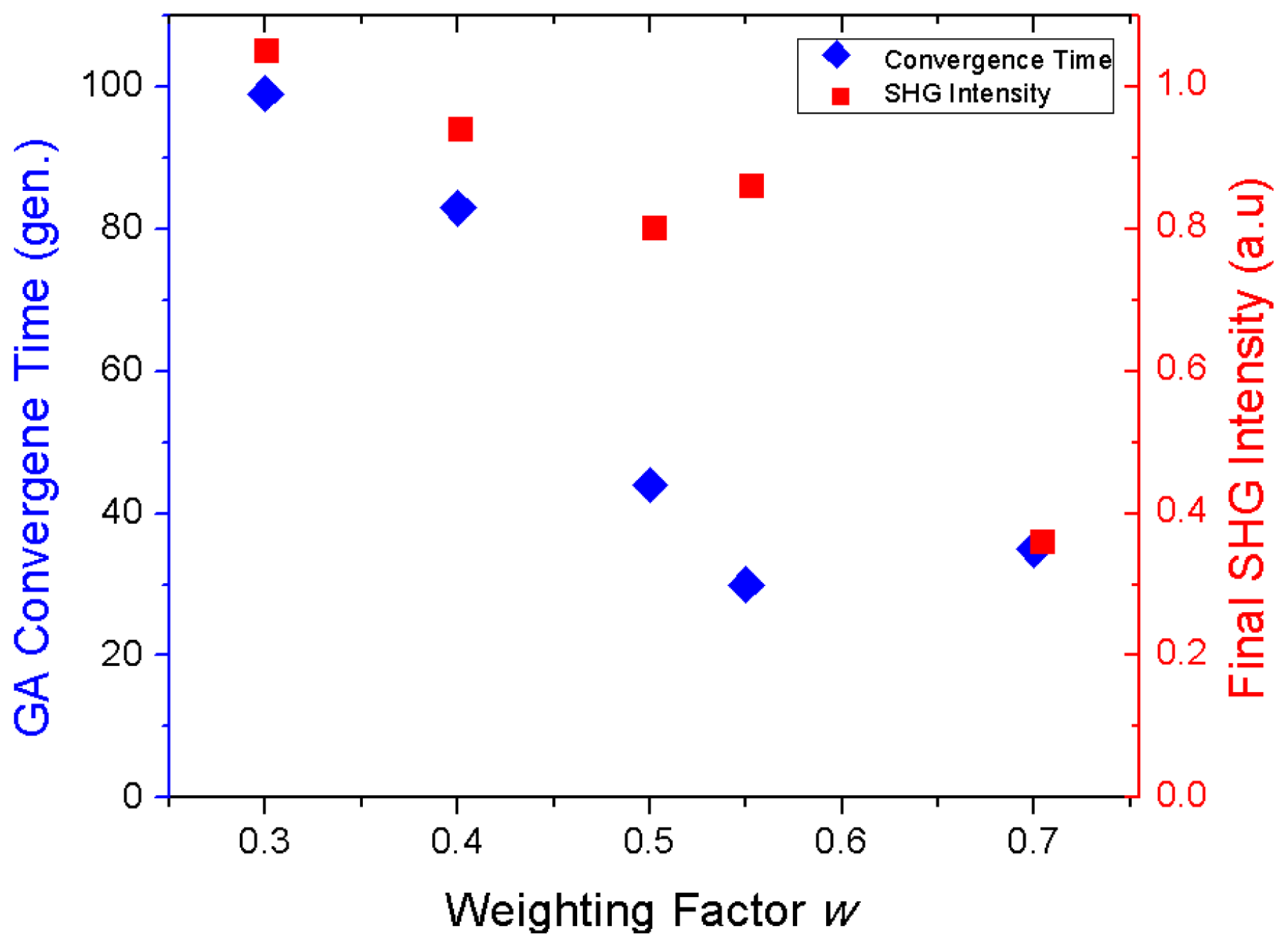
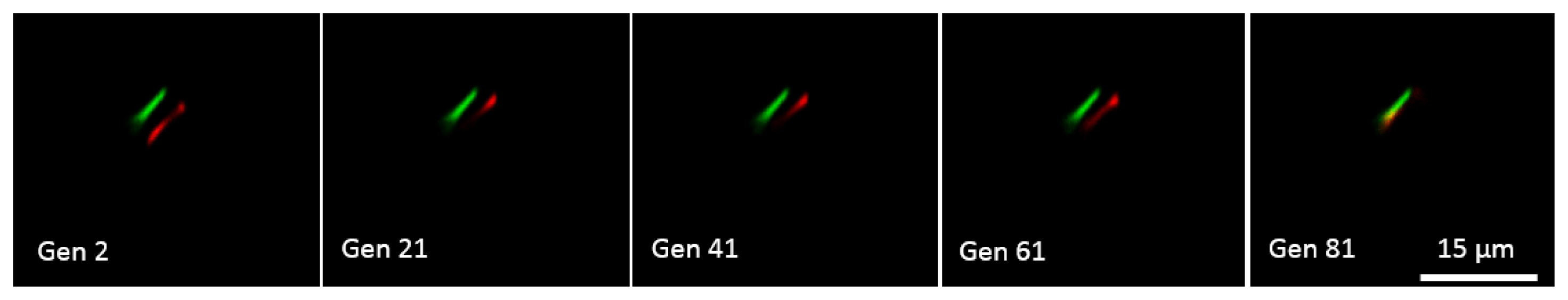
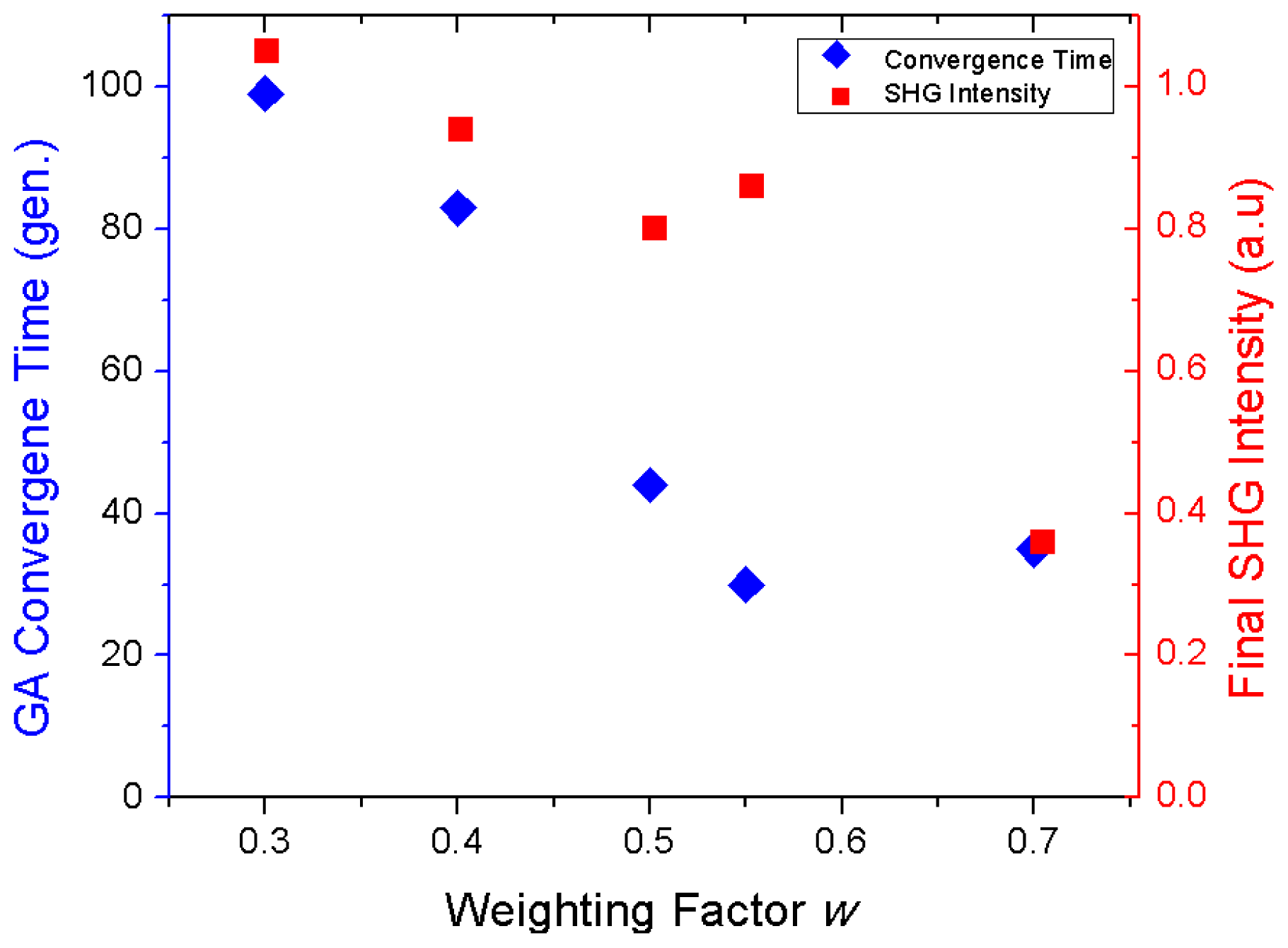
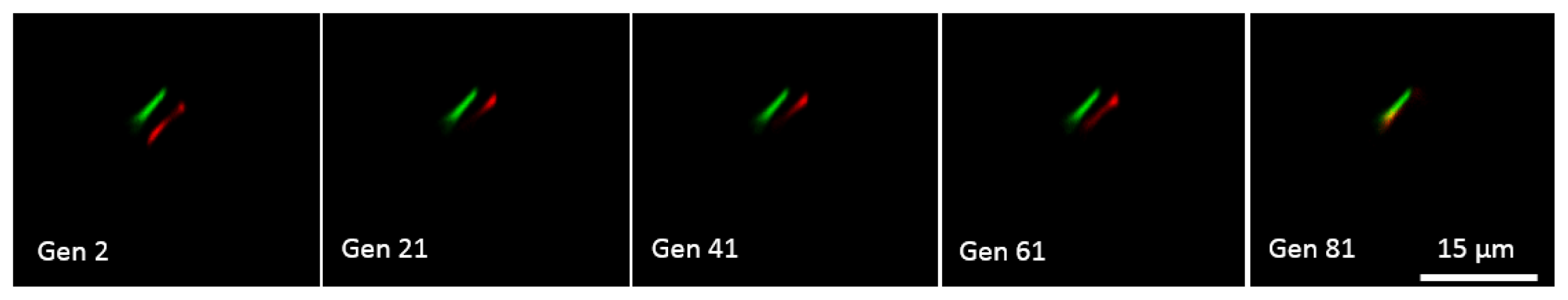
Genetic Algorithm for Multibeam Control in Differential Microscopy
3. Applications
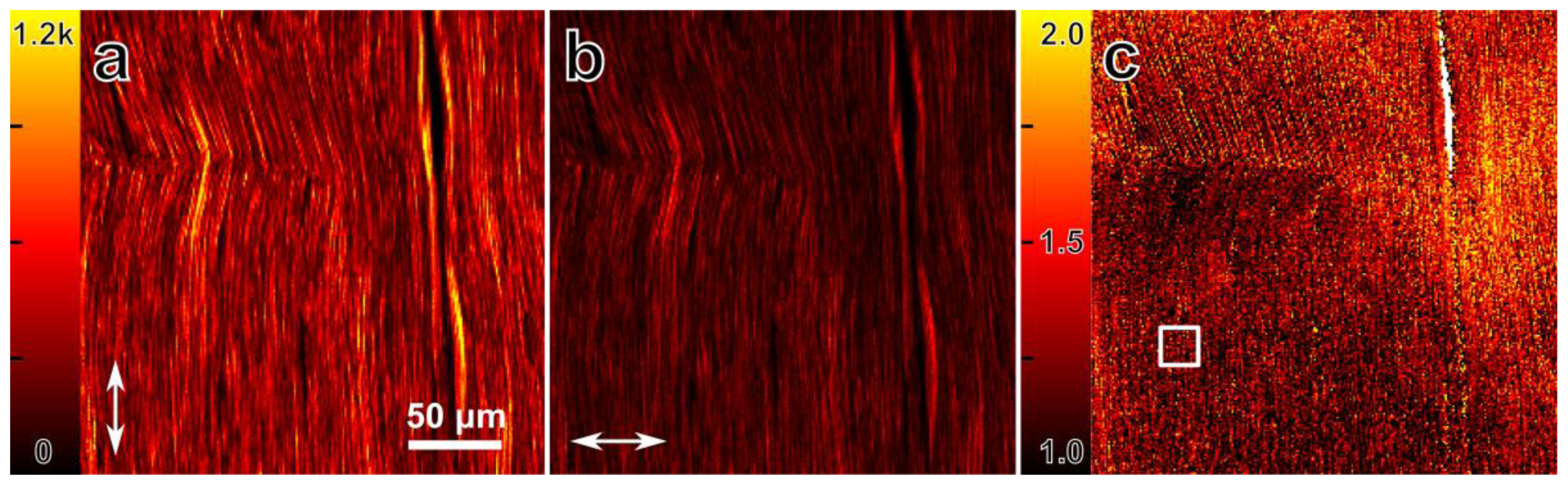
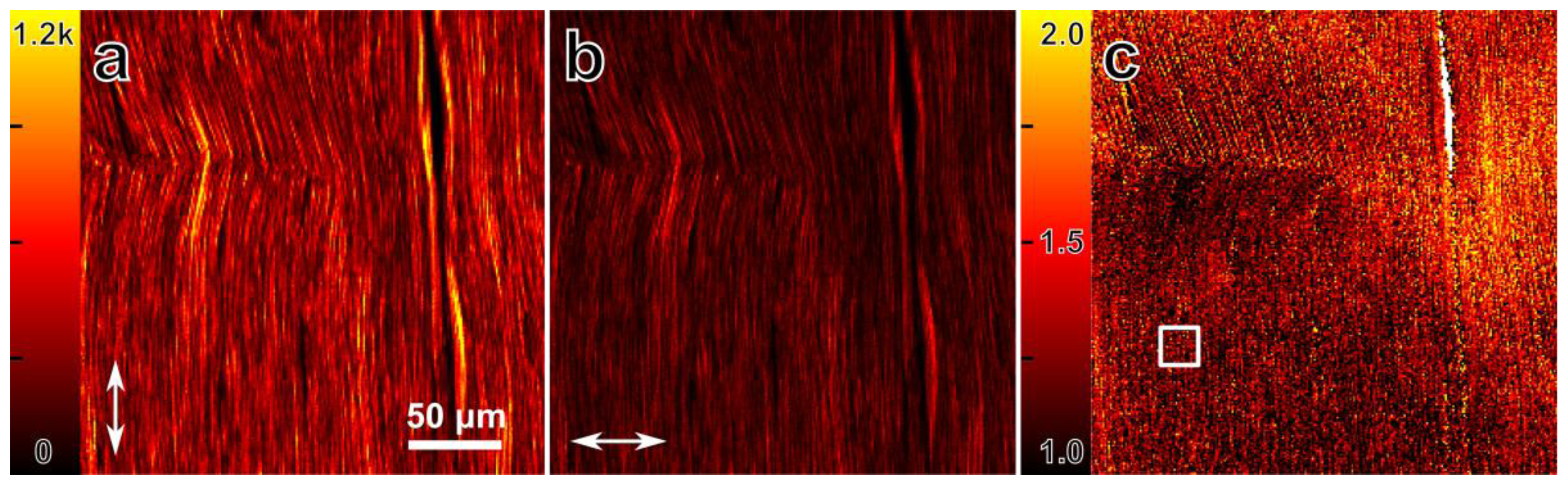
3.1. Differential Polarization SHG Microscopy of Collagen Fibers
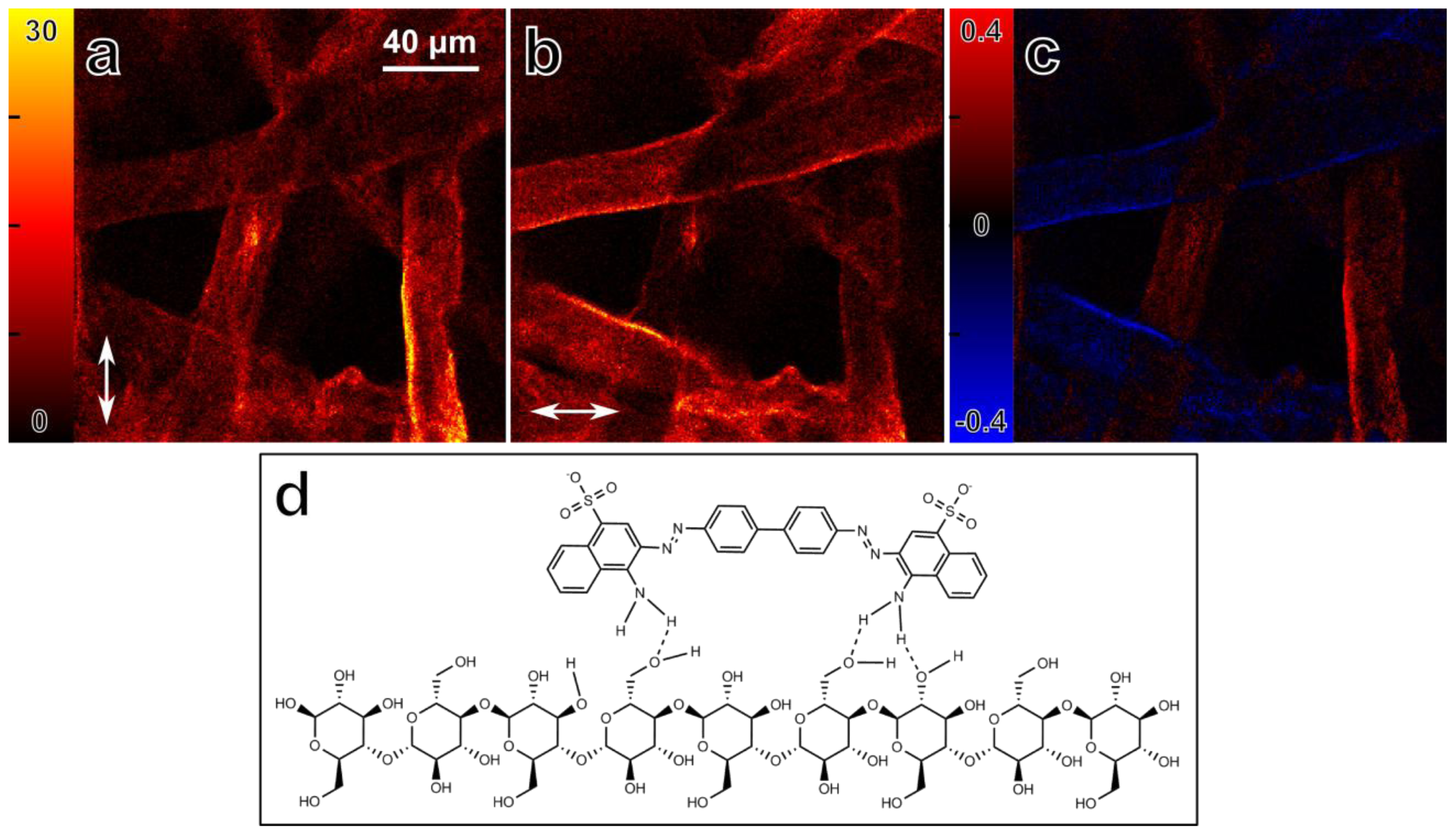
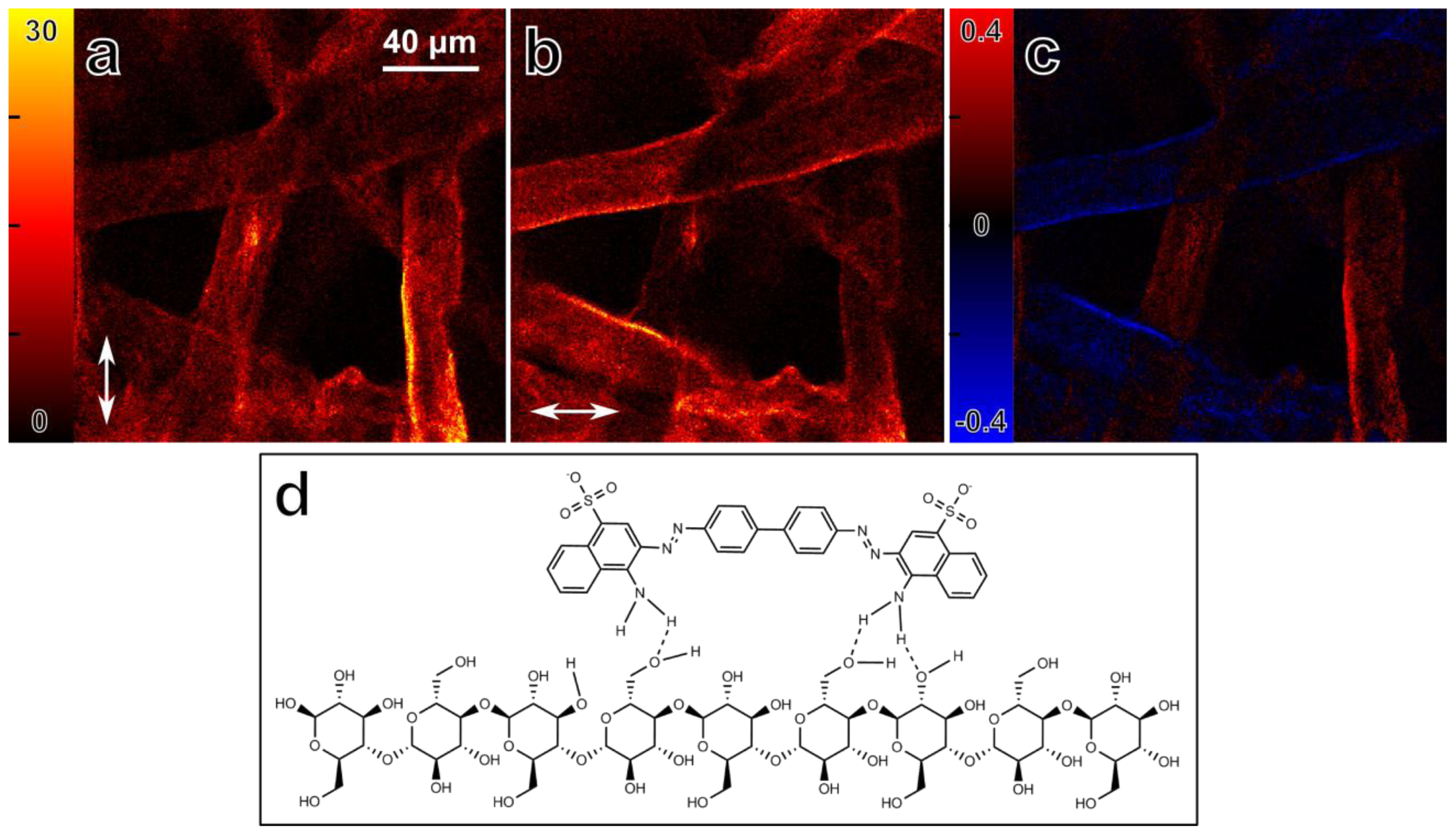
3.2. Absorption Anisotropy of Congo Red Stained Cellulose
3.3. Multicontrast Differential Polarization Microscopy
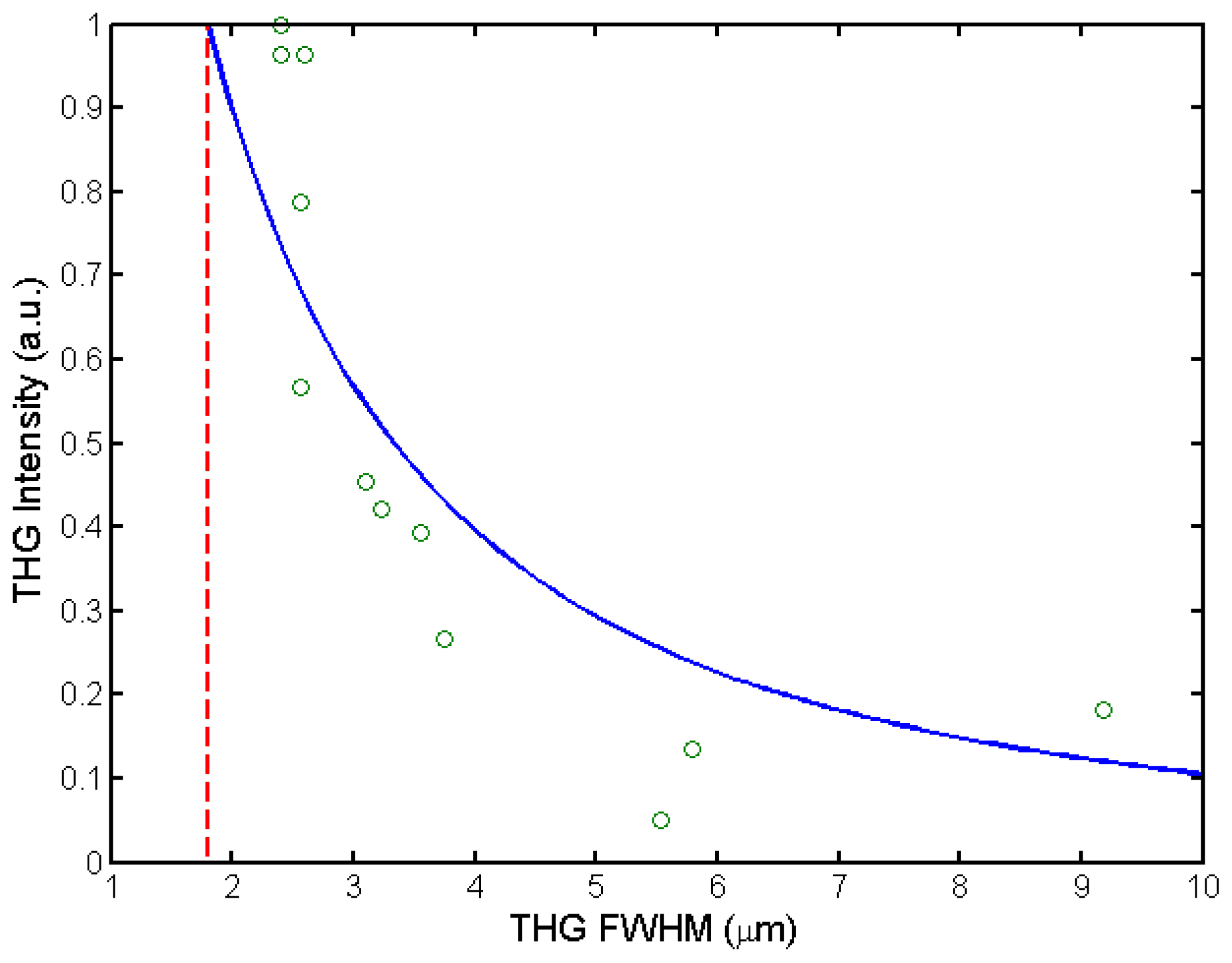
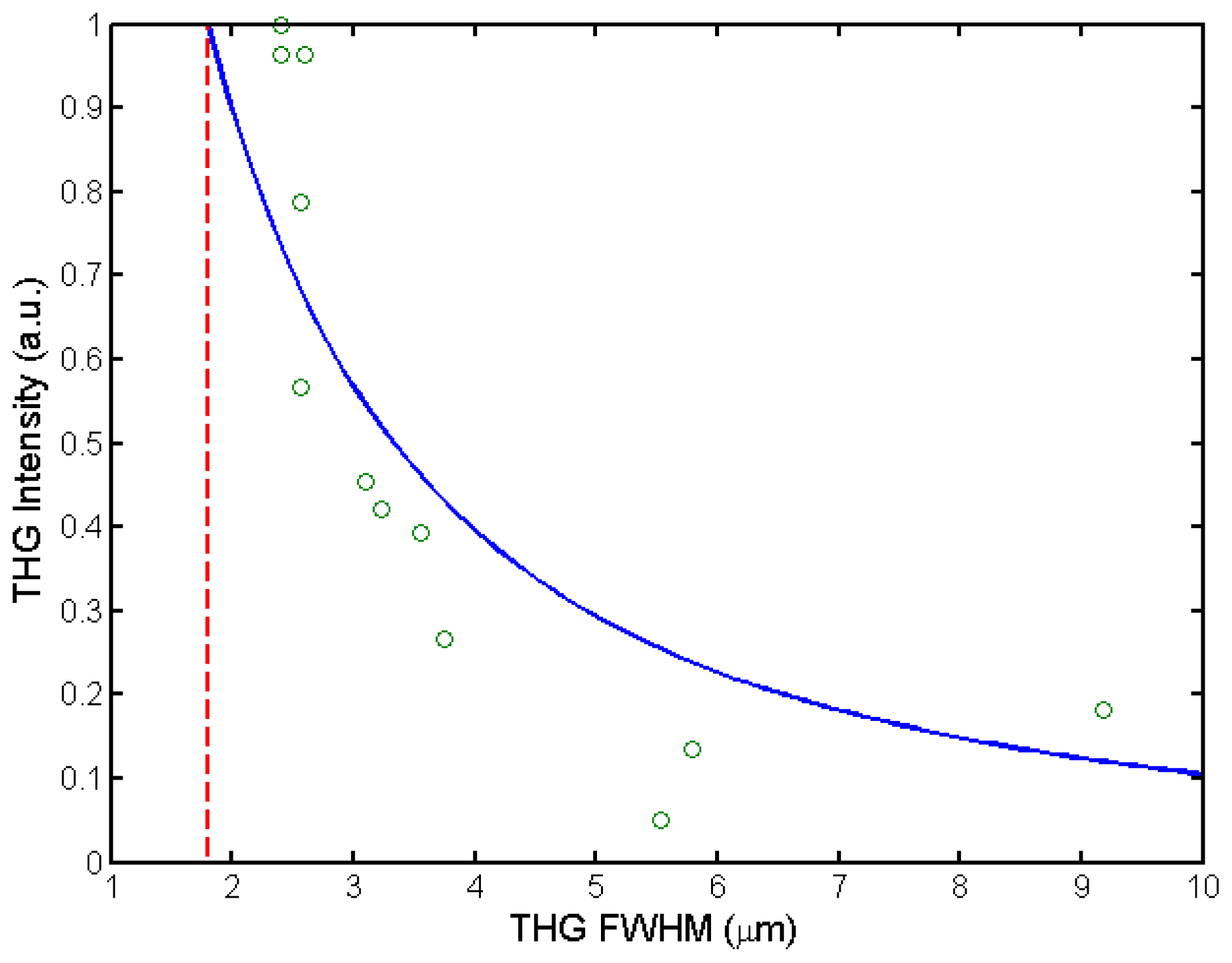
3.4. High Accuracy Beam Overlapping for Differential Polarization Microscopy
4. Conclusions and Outlook
Acknowledgments
Conflicts of Interest
References
- Carpenter, W.B.; Smith, F.G. The Microscope: And Its Revelations; Blanchard and Lea: Philadelphia, PA, USA, 1856. [Google Scholar]
- Hogg, J. The microscope: Its history, construction, and applications; George Routledge and Sons: London, UK, 1867. [Google Scholar]
- Juang, C.B.; Finzi, L.; Bustamante, C.J. Design and application of a computer-controlled confocal scanning differential polarization microscope. Rev. Sci. Instrum 1988, 59, 2399–2408. [Google Scholar]
- Finzi, L.; Bustamante, C.; Garab, G.; Juang, C.B. Direct observation of large chiral domains in chloroplast thylakoid membranes by differential polarization microscopy. Proc. Natl. Acad. Sci. USA 1989, 86, 8748–8752. [Google Scholar]
- Gorjanacz, M.; Torok, I.; Pomozi, I.; Garab, G.; Szlanka, T.; Kiss, I.; Mechler, B.M. Domains of importin-alpha 2 required for ring canal assembly during drosophila oogenesis. J. Struct. Biol 2006, 154, 27–41. [Google Scholar]
- Steinbach, G.; Pomozi, I.; Zsiros, O.; Menczel, L.; Garab, G. Imaging anisotropy using differential polarization laser scanning confocal microscopy. Acta Histochem 2009, 111, 317–326. [Google Scholar]
- Steinbach, G.; Pomozi, I.; Zsiros, O.; Pay, A.; Horvath, G.V.; Garab, G. Imaging fluorescence detected linear dichroism of plant cell walls in laser scanning confocal microscope. Cytom. Part A 2008, 73, 202–208. [Google Scholar]
- Denk, W.; Strickler, J.; Webb, W. Two-photon laser scanning fluorescence microscopy. Science 1990, 248, 73–76. [Google Scholar]
- Sheppard, C.J.R.; Wilson, T. Depth of field in scanning microscope. Opt. Lett 1978, 3, 115–117. [Google Scholar]
- Barad, Y.; Eisenberg, H.; Horowitz, M.; Silberberg, Y. Nonlinear scanning laser microscopy by third harmonic generation. Appl. Phys. Lett 1997, 70, 922–924. [Google Scholar]
- Tuer, A.E.; Krouglov, S.; Prent, N.; Cisek, R.; Sandkuijl, D.; Yasufuku, K.; Wilson, B.C.; Barzda, V. Nonlinear optical properties of type i collagen fibers studied by polarization dependent second harmonic generation microscopy. J. Phys. Chem. B 2011, 115, 12759–12769. [Google Scholar]
- Carriles, R.; Sheetz, K.E.; Hoover, E.E.; Squier, J.A.; Barzda, V. Simultaneous multifocal, multiphoton, photon counting microscopy. Opt. Express 2008, 16, 10364–10371. [Google Scholar]
- Chandler, E.; Hoover, E.; Field, J.; Sheetz, K.; Amir, W.; Carriles, R.; Ding, S.Y.; Squier, J. High-resolution mosaic imaging with multifocal, multiphoton photon-counting microscopy. Appl. Opt 2009, 48, 2067–2077. [Google Scholar]
- Field, J.J.; Sheetz, K.E.; Chandler, E.V.; Hoover, E.E.; Young, M.D.; Ding, S.Y.; Sylvester, A.W.; Kleinfeld, D.; Squier, J.A. Differential multiphoton laser scanning microscopy. IEEE J. Sel. Top. Quant 2012, 18, 14–28. [Google Scholar]
- Sheetz, K.E.; Squier, J. Ultrafast optics: Imaging and manipulating biological systems. J. Appl. Phys 2009, 105, 051101–051101. [Google Scholar]
- Squier, J.A.; Amir, W.; Carriles, R.; Chandler, E.; Field, J.J.; Hoover, E.E.; Schafer, D.; Sheetz, K.E. Multifocal, Multi-Modal, Photon Counting, Multiphoton Microscopy. Proceedings of Conference on Lasers and Electro-Optics and Quantum Electronics and Laser Science Conference, Baltimore, MD, USA, June 2009; pp. 371–372.
- Albert, O.; Sherman, L.; Mourou, G.; Norris, T.B.; Vdovin, G. Smart microscope: An adaptive optics learning system for aberration correction in multiphoton confocal microscopy. Opt. Lett 2000, 25, 52–54. [Google Scholar]
- Booth, M.J.; Débarre, D.; Jesacher, A. Adaptive optics for biomedical microscopy. Opt. Photon. News 2012, 23, 22–29. [Google Scholar]
- Gould, T.J.; Burke, D.; Bewersdorf, J.; Booth, M.J. Adaptive optics enables 3d sted microscopy in aberrating specimens. Opt. Express 2012, 20, 20998–21009. [Google Scholar]
- Planchon, T.A.; Amir, W.; Field, J.J.; Durfee, C.G.; Squier, J.A.; Rousseau, P.; Albert, O.; Mourou, G. Adaptive correction of a tightly focused, high-intensity laser beam by use of a third-harmonic signal generated at an interface. Opt. Lett 2006, 31, 2214–2216. [Google Scholar]
- Major, A.; Cisek, R.; Sandkuijl, D.; Barzda, V. Femtosecond Yb: KGD(WO4)2 laser with > 100 nJ of pulse energy. Laser Phys. Lett 2009, 6, 272–274. [Google Scholar]
- Sandkuijl, D.; Cisek, R.; Major, A.; Barzda, V. Differential microscopy for fluorescence-detected nonlinear absorption linear anisotropy based on a staggered two-beam femtosecond Yb:KGW oscillator. Biomed Opt Express 2010, 1, 895–901. [Google Scholar]
- Sheetz, K.E.; Hoover, E.E.; Carriles, R.; Kleinfeld, D.; Squier, J.A. Advancing multifocal nonlinear microscopy: Development and application of a novel multibeam Yb:KGD(WO4)2 oscillator. Opt. Express 2008, 16, 17574–17584. [Google Scholar]
- Samim, M.; Cisek, R.; Sandkuijl, D.; Musikhin, S.; Prent, N.; Stewart, B.; Barzda, V. Towards fast 3d microscopy: Multiple-depth focusing for video-rate imaging. Phys. In Can 2010, 66, 183–185. [Google Scholar]
- Squier, J.; Muller, M. High resolution nonlinear microscopy: A review of sources and methods for achieving optimal imaging. Rev. Sci. Instrum 2001, 72, 2855–2867. [Google Scholar]
- Goldberg, D.E. Genetic Algorithms in Search, Optimization and Machine Learning; Addison-Wesley Longman Publishing Co., Inc.: Reading, MA, USA, 1989; p. 372. [Google Scholar]
- Wright, A.J.; Burns, D.; Patterson, B.A.; Poland, S.P.; Valentine, G.J.; Girkin, J.M. Exploration of the optimisation algorithms used in the implementation of adaptive optics in confocal and multiphoton microscopy. Microsc. Res. Tech 2005, 67, 36–44. [Google Scholar]
- Poland, S.P.; Wright, A.J.; Girkin, J.M. Evaluation of fitness parameters used in an iterative approach to aberration correction in optical sectioning microscopy. Appl. Opt 2008, 47, 731–736. [Google Scholar]
- Betzig, E.; Patterson, G.H.; Sougrat, R.; Lindwasser, O.W.; Olenych, S.; Bonifacino, J.S.; Davidson, M.W.; Lippincott-Schwartz, J.; Hess, H.F. Imaging intracellular fluorescent proteins at nanometer resolution. Science 2006, 313, 1642–1645. [Google Scholar]
- Rust, M.J.; Bates, M.; Zhuang, X. Sub-diffraction-limit imaging by stochastic optical reconstruction microscopy (storm). Nat. Meth 2006, 3, 793–796. [Google Scholar]
- Chu, S.W.; Chen, S.Y.; Chern, G.W.; Tsai, T.H.; Chen, Y.C.; Lin, B.L.; Sun, C.K. Studies of χ(2)/χ(3) tensors in submicron-scaled bio-tissues by polarization harmonics optical microscopy. Biophys. J 2004, 86, 3914–3922. [Google Scholar]
- Su, P.J.; Chen, W.L.; Chen, Y.F.; Dong, C.Y. Determination of collagen nanostructure from second-order susceptibility tensor analysis. Biophys. J 2011, 100, 2053–2062. [Google Scholar]
- Freund, I.; Deutsch, M.; Sprecher, A. Connective-tissue polarity. Optical 2nd harmonic microscopy, crossed-beam summation, and small-angle scattering in rat-tail tendon. Biophys. J 1986, 50, 693–712. [Google Scholar]
- Tuer, A.E.; Akens, M.K.; Krouglov, S.; Sandkuijl, D.; Wilson, B.C.; Whyne, C.M.; Barzda, V. Hierarchical model of fibrillar collagen organization for interpreting the second-order susceptibility tensors in biological tissue. Biophys. J 2012, 103, 2093–2105. [Google Scholar]
- Puchtler, H.; Sweat, F.; Levine, M. On binding of congo red by amyloid. J. Histochem. Cytochem 1962, 10, 355. [Google Scholar]
- Taylor, D.L.; Allen, R.D.; Benditt, E.P. Determination of polarization optical-properties of amyloid-congo red complex by phase modulation microspectrophotometry. J. Histochem. Cytochem 1974, 22, 1105–1112. [Google Scholar]
- Brown, R.M.; Millard, A.C.; Campagnola, P.J. Macromolecular structure of cellulose studied by second-harmonic generation imaging microscopy. Opt. Lett 2003, 28, 2207–2209. [Google Scholar]
- Philipose, U.; Sun, P.; Xu, T.; Ruda, H.E.; Yang, L.; Kavanagh, K.L. Structure and photoluminescence of ZnSe nanostructures fabricated by vapor phase growth. J. Appl. Phys 2007, 101, 014326–014325. [Google Scholar]
- Philipose, U.; Xu, T.; Yang, S.; Sun, P.; Ruda, H.E.; Wang, Y.Q.; Kavanagh, K.L. Enhancement of band edge luminescence in ZnSe nanowires. J. Appl. Phys 2006, 100, 084316–084316. [Google Scholar]
- Debarre, D.; Booth, M.J.; Wilson, T. Image based adaptive optics through optimisation of low spatial frequencies. Opt. Express 2007, 15, 8176–8190. [Google Scholar]
- Boulesteix, T.; Beaurepaire, E.; Sauviat, M.P.; Schanne-Klein, M.C. Second-harmonic microscopy of unstained living cardiac myocytes: Measurements of sarcomere length with 20-nm accuracy. Opt. Lett 2004, 29, 2031–2033. [Google Scholar]
- Nucciotti, V.; Stringari, C.; Sacconi, L.; Vanzi, F.; Fusi, L.; Linari, M.; Piazzesi, G.; Lombardi, V.; Pavone, F.S. Probing myosin structural conformation in vivo by second-harmonic generation microscopy. Proc. Natl. Acad. Sci. USA 2010, 107, 7763–7768. [Google Scholar]

Supplementary Files
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Samim, M.; Sandkuijl, D.; Tretyakov, I.; Cisek, R.; Barzda, V. Differential Polarization Nonlinear Optical Microscopy with Adaptive Optics Controlled Multiplexed Beams. Int. J. Mol. Sci. 2013, 14, 18520-18534. https://doi.org/10.3390/ijms140918520
Samim M, Sandkuijl D, Tretyakov I, Cisek R, Barzda V. Differential Polarization Nonlinear Optical Microscopy with Adaptive Optics Controlled Multiplexed Beams. International Journal of Molecular Sciences. 2013; 14(9):18520-18534. https://doi.org/10.3390/ijms140918520
Chicago/Turabian StyleSamim, Masood, Daaf Sandkuijl, Ian Tretyakov, Richard Cisek, and Virginijus Barzda. 2013. "Differential Polarization Nonlinear Optical Microscopy with Adaptive Optics Controlled Multiplexed Beams" International Journal of Molecular Sciences 14, no. 9: 18520-18534. https://doi.org/10.3390/ijms140918520
APA StyleSamim, M., Sandkuijl, D., Tretyakov, I., Cisek, R., & Barzda, V. (2013). Differential Polarization Nonlinear Optical Microscopy with Adaptive Optics Controlled Multiplexed Beams. International Journal of Molecular Sciences, 14(9), 18520-18534. https://doi.org/10.3390/ijms140918520