Increased Circulatory Asymmetric Dimethylarginine and Multiple Organ Failure: Bile Duct Ligation in Rat as a Model



{kind=link}
Abstract
:1. Introduction
2. Asymmetric Dimethylarginine (ADMA) Metabolism
3. ADMA Regulation in Normal Liver Function
4. Increased Circulatory and Hepatic ADMA Concentrations in the Context of Liver Dysfunction
5. The Role of Increased Circulatory ADMA in Multiple Organ Failure in Critical Illness
6. Pathogenic Mechanisms of ADMA in Cell and Organ Metabolism
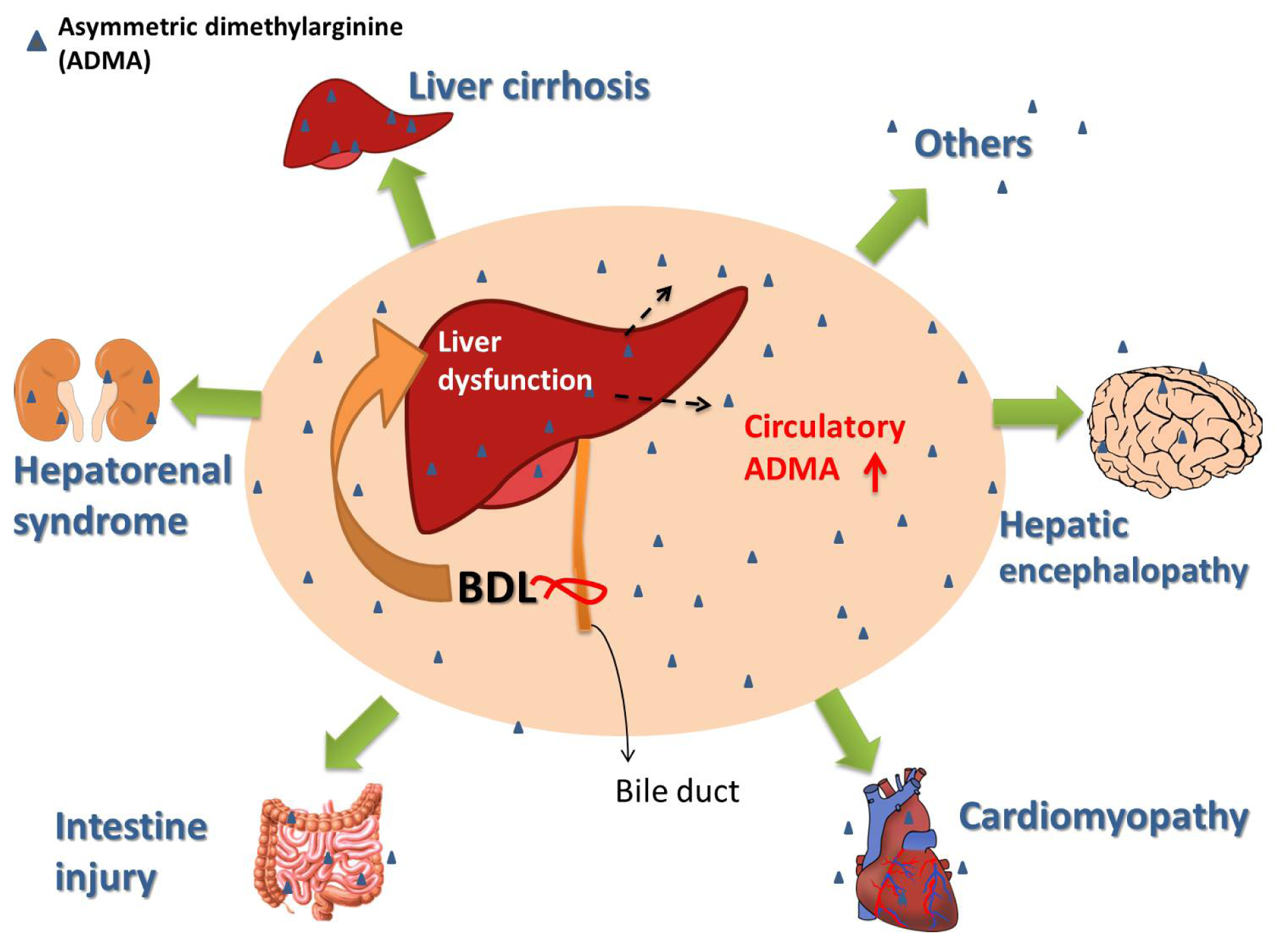
7. Bile Duct Ligation (BDL)-Induced Liver Damages in Rat
8. Plasma ADMA, Symmetric Dimethylarginine (SDMA) and l-Arginine Concentrations in the BDL Rat
9. Possible Role of Increased Circulatory ADMA in the Multiple Organ Damages Observed in the BDL Rat
9.1. ADMA and Nitric Oxide (NO) Dysregulation in the BDL Rat Liver
9.2. ADMA and NO Dysregulation in the BDL Rat Kidney
9.3. ADMA and NO Dysregulation in the BDL Rat Brain
9.4. Other Major Organ Involvement in the Multiple Organ Failure (MOF) Model and BDL in Terms of ADMA and NO Dysregulation
10. Conclusions
Acknowledgments
Conflicts of Interest
Abbreviations
| ADMA | asymmetric dimethylarginine |
| BH4 | tetrahydrobiopterin |
| BDL | bile duct ligation |
| CAT | cationic amino acid transporter |
| cGMP | cyclic guanosine monophosphate |
| DDAH | dimethylarginine dimethylaminohydrolase |
| eNOS | endothelial NOS |
| HE | hepatic encephalopathy |
| HRS | hepatorenal syndrome |
| iNOS | inducible NOS |
| MELD | Model for End-Stage Liver Disease |
| MOF | multiple organ failure |
| NO | nitric oxide |
| NOS | nitric oxide synthase |
| PRMT | protein arginine methyltransferase |
| SDMA | symmetric dimethylarginine |
| TIPS | transjugular intrahepatic portosystemic shunt |
References
- Vallance, P.; Leone, A.; Calver, A.; Collier, J.; Moncada, S. Endogenous dimethylarginine as an inhibitor of nitric oxide synthesis. J. Cardiovasc. Pharmacol 1992, 20, S60–S62. [Google Scholar]
- Kurose, I.; Wolf, R.; Grisham, M.B.; Granger, D.N. Effects of an endogenous inhibitor of nitric oxide synthesis on postcapillary venules. Am. J. Physiol 1995, 268, H2224–H2231. [Google Scholar]
- Faraci, F.M.; Brian, J.E., Jr; Heistad, D.D. Response of cerebral blood vessels to an endogenous inhibitor of nitric oxide synthase. Am. J. Physiol 1995, 269, H1522–H1527. [Google Scholar]
- Segarra, G.; Medina, P.; Ballester, R.M.; Lluch, P.; Aldasoro, M.; Vila, J.M.; Lluch, S.; Pelligrino, D.A. Effects of some guanidino compounds on human cerebral arteries. Stroke 1999, 30, 2206–2210. [Google Scholar]
- Vida, G.; Sulyok, E.; Ertl, T.; Martens-Lobenhoffer, J.; Bode-Boger, S.M. Plasma asymmetric dimethylarginine concentration during the perinatal period. Neonatology 2007, 92, 8–13. [Google Scholar]
- Tsikas, D. A critical review and discussion of analytical methods in the l-arginine/nitric oxide area of basic and clinical research. Anal. Biochem 2008, 379, 139–163. [Google Scholar]
- Lücke, T.; Kanzelmeyer, N.; Kemper, M.J.; Tsikas, D.; Das, A.M. Developmental changes in the l-arginine/nitric oxide pathway from infancy to adulthood: Plasma asymmetric dimethylarginine levels decrease with age. Clin. Chem. Lab. Med 2007, 45, 1525–1530. [Google Scholar]
- Vida, G.; Sulyok, E.; Ertl, T.; Martens-Lobenhoffer, J.; Bode-Böger, S.M. Birth by cesarean section is associated with elevated neonatal plasma levels of dimethylarginines. Pediatr. Int 2012, 54, 476–479. [Google Scholar]
- Bode-Böger, S.M.; Scalera, F.; Ignarro, L.J. The l-arginine paradox: Importance of the l-arginine/asymmetrical dimethylarginine ratio. Pharmacol. Ther 2007, 114, 295–306. [Google Scholar]
- Bode-Böger, S.M.; Muke, J.; Surdacki, A.; Brabant, G.; Böger, R.H.; Frölich, J.C. Oral l-arginine improves endothelial function in healthy individuals older than 70 years. Vasc. Med 2003, 8, 77–81. [Google Scholar]
- Tain, Y.L.; Huang, L.T. Asymmetric dimethylarginine: Clinical applications in pediatric medicine. J. Formos. Med. Assoc 2011, 110, 70–77. [Google Scholar]
- Cooke, J.P. Does ADMA cause endothelial dysfunction? Arterioscler. Thromb. Vasc. Biol 2000, 20, 2032–2037. [Google Scholar]
- Vallance, P.; Leiper, J. Cardiovascular biology of the asymmetric dimethylarginine: Dimethylarginine dimethylaminohydrolase pathway. Arterioscler. Thromb. Vasc. Biol 2004, 24, 1023–1030. [Google Scholar]
- Baylis, C. Arginine arginine analogs and nitric oxide production in chronic kidney disease. Nat. Clin. Pract. Nephrol 2006, 2, 209–220. [Google Scholar]
- Cardounel, A.J.; Cui, H.; Samouilov, A.; Johnson, W.; Kearns, P.; Tsai, A.L.; Berka, V.; Zweier, J.L. Evidence for the pathophysiological role of endogenous methylarginines in regulation of endothelial NO production and vascular function. J. Biol. Chem 2007, 282, 879–887. [Google Scholar]
- Böger, R.H. Asymmetric dimethylarginine (ADMA): A novel risk marker in cardiovascular medicine and beyond. Ann. Med 2006, 38, 126–136. [Google Scholar]
- Cooke, J.P. ADMA: Its role in vascular disease. Vasc. Med 2005, 10, S11–S17. [Google Scholar]
- Laleman, W.; Omasta, A.; van de Casteele, M.; Zeegers, M.; Vander, E.; Ivan Landeghem, L.; Severi, T.; van Pelt, J.; Roskams, T.; Fevery, J.; et al. A role for asymmetric dimethylarginine in the pathophysiology of portal hypertension in rats with biliary cirrhosis. Hepatology 2005, 42, 1382–1390. [Google Scholar]
- Wang, D.; Strandgaard, S.; Iversen, J.; Wilcox, C.S. Asymmetric dimethylarginine oxidative stress and vascular nitric oxide synthase in essential hypertension. Am. J. Physiol. Regul. Integr. Comp. Physiol 2009, 296, R195–R200. [Google Scholar]
- Perticone, F.; Sciacqua, A.; Maio, R.; Perticone, M.; Galiano, L.G.; Bruni, R.; di Cello, S.; Pascale, A.; Talarico, G.; Greco, L.; et al. Endothelial dysfunction, ADMA and insulin resistance in essential hypertension. Int. J. Cardiol. 2010, 142, 236–241. [Google Scholar]
- Najbauer, J.; Johnson, B.A.; Young, A.L.; Aswad, D.W. Peptides with sequences similar to glycine arginine-rich motifs in proteins interacting with RNA are efficiently recognized by methyltransferase(s) modifying arginine in numerous proteins. J. Biol. Chem 1993, 268, 10501–10509. [Google Scholar]
- Bedford, M.T.; Clarke, S.G. Protein arginine methylation in mammals: Who what and why. Mol. Cell 2009, 33, 1–13. [Google Scholar]
- Closs, E.I.; Basha, F.Z.; Habermeier, A.; Förstermann, U. Interference of l-arginineanalogues with l-arginine transport mediated by the y+ carrier hCAT-2B. Nitric Oxide 1997, 1, 65–73. [Google Scholar]
- Strobel, J.; Mieth, M.; Endress, B.; Auge, D.; König, J.; Fromm, M.F.; Maas, R. Interaction of the cardiovascular risk marker asymmetric dimethylarginine (ADMA) with the human cationic amino acid transporter 1 (CAT1). J. Mol. Cell. Cardiol 2012, 53, 392–400. [Google Scholar]
- Teerlink, T.; Luo, Z.; Palm, F.; Wilcox, C.S. Cellular ADMA: Regulation and action. Pharmacol. Res 2009, 60, 448–460. [Google Scholar]
- Wilcken, D.E.; Sim, A.S.; Wang, J.; Wang, X.L. Asymmetric dimethylarginine (ADMA) in vascular renal and hepatic disease and the regulatory role of l-arginine on its metabolism. Mol. Genet. Metab 2007, 91, 309–317. [Google Scholar]
- Bulau, P.; Zakrzewicz, D.; Kitowska, K.; Leiper, J.; Gunther, A.; Grimminger, F.; Eickelberg, O. Analysis of methylarginine metabolism in the cardiovascular system identifies the lung as a major source of ADMA. Am. J. Physiol. Lung Cell. Mol. Physiol. 2007, 292, L18–L24. [Google Scholar]
- Wang, J.; Sim, A.S.; Wang, X.L.; Wilcken, D.E. l-Arginine regulates asymmetric dimethylarginine metabolism by inhibiting dimethylarginine dimethylaminohydrolase activity in hepatic (HepG2) cells. Cell. Mol. Life Sci. 2006, 63, 2838–2846. [Google Scholar]
- Aldámiz-Echevarría, L.; Andrade, F. Asymmetric dimethylarginine endothelial dysfunction and renal disease. Int. J. Mol. Sci 2012, 13, 11288–11311. [Google Scholar]
- Carnegie, P.R.; Fellows, F.C.; Symington, G.R. Urinary excretion of methylarginine in human disease. Metabolism 1977, 26, 531–537. [Google Scholar]
- Nijveldt, R.J.; Teerlink, T.; Siroen, M.P.; van Lambalgen, A.A.; Rauwerda, J.A.; van Leeuwen, P.A. The liver is an important organ in the metabolism of asymmetrical dimethylarginine (ADMA). Clin. Nutr. 2003, 22, 17–22. [Google Scholar]
- Nijveldt, R.J.; Siroen, M.P.; van der Hoven, B.; Teerlink, T.; Prins, H.A.; Girbes, A.R.; van Leeuwen, P.A. High plasma arginine concentrations in critically ill patients suffering from hepatic failure. Eur. J. Clin. Nutr 2004, 58, 587–593. [Google Scholar] [Green Version]
- Teerlink, T. ADMA metabolism and clearance. Vasc. Med 2005, 10, S73–S81. [Google Scholar]
- Nijveldt, R.J.; Teerlink, T.; Siroen, M.P.; van der Hoven, B.; Prins, H.A.; Wiezer, M.J.; Meijer, C.; van der Sijp, J.R.; Cuesta, M.A.; Meijer, S.; et al. Elevation of asymmetric dimethylarginine (ADMA) in patients developing hepatic failure after major hepatectomy. J. Parenter. Enter. Nutr 2004, 28, 382–387. [Google Scholar]
- Mookerjee, R.P.; Dalton, R.N.; Davies, N.A.; Hodges, S.J.; Turner, C.; Williams, R.; Jalan, R. Inflammation is an important determinant of levels of the endogenous nitric oxide synthase inhibitor asymmetric dimethylarginine (ADMA) in acute liver failure. Liver Transplant 2007, 13, 400–405. [Google Scholar]
- Lluch, P.; Torondel, B.; Medina, P.; Segarra, G.; del Olmo, J.A.; Serra, M.A.; Rodrigo, J.M. Plasma concentrations of nitric oxide and asymmetric dimethylarginine in human alcoholic cirrhosis. J. Hepatol 2004, 41, 55–59. [Google Scholar]
- Davids, M.; Richir, M.C.; Visser, M.; Ellger, B.; van den Berghe, G.; van Leeuwen, P.A.; Teerlink, T. Role of dimethylarginine dimethylaminohydrolase activity in regulation of tissue and plasma concentrations of asymmetric dimethylarginine in an animal model of prolonged critical illness. Metabolism 2012, 61, 482–490. [Google Scholar]
- Nijveldt, R.J.; Teerlink, T.; van Leeuwen, P.A. The asymmetrical dimethylarginine (ADMA)-multiple organ failure hypothesis. Clin. Nutr 2003, 22, 99–104. [Google Scholar]
- Milstien, S.; Katusic, Z. Oxidation of tetrahydrobiopterin by peroxynitrite: Implications for vascular endothelial function. Biochem. Biophys. Res. Commun 1999, 263, 681–684. [Google Scholar]
- Rochette, L.; Lorin, J.; Zeller, M.; Guilland, J.C.; Lorgis, L.; Cottin, Y.; Vergely, C. Nitric oxide synthase inhibition and oxidative stress in cardiovascular diseases: Possible therapeutic targets? Pharmacol. Ther 2013, 140, 239–257. [Google Scholar]
- Wells, S.M.; Holian, A. Asymmetric dimethylarginine induces oxidative and nitrosative stress in murine lung epithelial cells. Am. J. Respir. Cell Mol. Biol 2007, 36, 520–528. [Google Scholar]
- Sydow, K.; Münzel, T. ADMA and oxidative stress. Atheroscler. Suppl 2003, 4, 41–51. [Google Scholar]
- Lee, D.Y.; Teyssier, C.; Strahl, B.D.; Stallcup, M.R. Role of protein methylation in regulation of transcription. Endocr. Rev 2005, 26, 147–170. [Google Scholar]
- Cerwinka, W.H.; Cooper, D.; Krieglstein, C.F.; Feelisch, M.; Granger, D.N. Nitric oxide modulates endotoxin-induced platelet-endothelial cell adhesion in intestinal venules. Am. J. Physiol. Heart Circ. Physiol 2002, 282, H1111–H1117. [Google Scholar]
- Jeremy, J.Y.; Rowe, D.; Emsley, A.M.; Newby, A.C. Nitric oxide and the proliferation of vascular smooth muscle cells. Cardiovasc. Res 1999, 43, 580–594. [Google Scholar]
- Fursule, R.A.; Patil, S.D. Hepatoprotective and antioxidant activity of Phaseolus trilobus Ait on bile duct ligation induced liver fibrosis in rats. J. Ethnopharmacol 2010, 129, 416–419. [Google Scholar]
- Canbakan, B.; Akin, H.; Tahan, G.; Tarcin, O.; Eren, F.; Atug, O.; Tahan, V.; Imeryuz, N.; Yapicier, O.; Avsar, E.; et al. The effects of pegylated interferon α 2b on bile-duct ligation induced liver fibrosis in rats. Ann. Hepatol 2009, 8, 234–240. [Google Scholar]
- Coban, S.; Yildiz, F.; Terzi, A.; Al, B.; Aksoy, N.; Bitiren, M.; Celik, H. The effects of Nigella sativa on bile duct ligation induced-liver injury in rats. Cell Biochem. Funct 2010, 28, 83–88. [Google Scholar]
- Tahan, G.; Eren, F.; Tarçin, O.; Akin, H.; Tahan, V.; Şahın, H.; Özdoğan, O.; İmeryüz, N.; Çelıkel, Ç; Avşar, E.; et al. Effects of a long-acting somatostatin analogue lanreotide on bile duct ligation-induced liver fibrosis in rats. Turk. J. Gastroenterol 2010, 21, 287–292. [Google Scholar]
- Tain, Y.L.; Hsieh, C.S.; Chen, C.C.; Sheen, J.M.; Lee, C.T.; Huang, L.T. Melatonin prevents increased asymmetric dimethylarginine in young rats with bile duct ligation. J. Pineal Res 2010, 48, 212–221. [Google Scholar]
- Marques, T.G.; Chaib, E.; da Fonseca, J.H.; Lourenço, A.C.; Silva, F.D.; Ribeiro, M.A., Jr; Galvão, F.H.; D’Albuquerque, L.A. Review of experimental models for inducing hepatic cirrhosis by bile duct ligation and carbon tetrachloride injection. Acta Cir. Bras 2012, 27, 589–594. [Google Scholar] [Green Version]
- Ayvaz, S.; Kanter, M.; Aksu, B.; Sahin, S.H.; Uzun, H.; Erboga, M.; Pul, M. The effects of hyperbaric oxygen application against cholestatic oxidative stress and hepatic damage after bile duct ligation in rats. J. Surg. Res 2013, 183, 146–155. [Google Scholar]
- Kim, H.G.; Han, J.M.; Lee, H.W.; Lee, J.S.; Son, S.W.; Choi, M.K.; Lee, D.S.; Wang, J.H.; Son, C.G. CGX a multiple herbal drug improves cholestatic liver fibrosis in a bile duct ligation-induced rat model. J. Ethnopharmacol 2013, 145, 653–662. [Google Scholar]
- Kim, T.W.; Lee, H.K.; Song, I.B.; Lim, J.H.; Cho, E.S.; Son, H.Y.; Jung, J.Y.; Yun, H.I. Platycodin D attenuates bile duct ligation-induced hepatic injury and fibrosis in mice. Food Chem. Toxicol 2013, 51, 364–369. [Google Scholar]
- Han, J.M.; Kim, H.G.; Choi, M.K.; Lee, J.S.; Park, H.J.; Wang, J.H.; Lee, J.S.; Son, S.W.; Hwang, S.Y.; Son, C.G. Aqueous extract of Artemisia iwayomogi Kitamura attenuates cholestatic liver fibrosis in a rat model of bile duct ligation. Food Chem. Toxicol 2012, 50, 3505–3513. [Google Scholar]
- Veidal, S.S.; Vassiliadis, E.; Bay-Jensen, A.C.; Tougas, G.; Vainer, B.; Karsdal, M.A. Procollagen type I N-terminal propeptide (PINP) is a marker for fibrogenesis in bile duct ligation-induced fibrosis in rats. Fibrogenesis Tissue Repair 2010, 3, 5:1–5:12. [Google Scholar]
- Sheen, J.M.; Huang, L.T.; Hsieh, C.S.; Chen, C.C.; Wang, J.Y.; Tain, Y.L. Bile duct ligation in developing rats: Temporal progression of liver kidney and brain damage. J. Pediatr. Surg 2010, 45, 1650–1658. [Google Scholar]
- Yang, Y.Y.; Lee, T.Y.; Huang, Y.T.; Chan, C.C.; Yeh, Y.C.; Lee, F.Y.; Lee, S.D.; Lin, H.C. Asymmetric dimethylarginine (ADMA) determines the improvement of hepatic endothelial dysfunction by vitamin E in cirrhotic rats. Liver Int 2012, 32, 48–57. [Google Scholar]
- Huang, L.T.; Hung, J.F.; Chen, C.C.; Hsieh, C.S.; Yu, H.R.; Hsu, C.N.; Tain, Y.L. Endotoxemia exacerbates kidney injury and increases asymmetric dimethylarginine in young bile duct-ligated rats. Shock 2012, 37, 441–448. [Google Scholar]
- Schwedhelm, E.; Maas, R.; Freese, R.; Jung, D.; Lukacs, Z.; Jambrecina, A.; Spickler, W.; Schulze, F.; Böger, R.H. Pharmacokinetic and pharmacodynamic properties of oral l-citrulline and l-arginine: Impact on nitric oxide metabolism. Br. J. Clin. Pharmacol 2008, 65, 51–59. [Google Scholar]
- Ljubuncic, P.; Tanne, Z.; Bomzon, A. Evidence of a systemic phenomenon for oxidative stress in cholestatic liver disease. Gut 2000, 47, 710–716. [Google Scholar]
- Assimakopoulos, S.F.; Mavrakis, A.G.; Grintzalis, K.; Papapostolou, I.; Zervoudakis, G.; Konstantinou, D.; Chroni, E.; Vagianos, C.E.; Georgiou, C. Superoxide radical formation in diverse organs of rats with experimentally induced obstructive jaundice. Redox. Rep 2008, 13, 179–184. [Google Scholar]
- Grintzalis, K.; Papapostolou, I.; Assimakopoulos, S.F.; Mavrakis, A.; Faropoulos, K.; Karageorgos, N.; Georgiou, C.; Chroni, E.; Konstantinou, D. Time-related alterations of superoxide radical levels in diverse organs of bile duct-ligated rats. Free Radic. Res 2009, 43, 803–808. [Google Scholar]
- Huang, L.T.; Tiao, M.M.; Tain, Y.L.; Chen, C.C.; Hsieh, C.S. Melatonin ameliorates bile duct ligation-induced systemic oxidative stress and spatial memory deficits in developing rats. Pediatr. Res 2009, 65, 176–180. [Google Scholar]
- Tain, Y.L.; Kao, Y.H.; Hsieh, C.S.; Chen, C.C.; Sheen, J.M.; Lin, I.C.; Huang, L.T. Melatonin blocks oxidative stress-induced increased asymmetric dimethylarginine. Free Radic. Biol. Med 2010, 49, 1088–1098. [Google Scholar]
- Pinzani, M.; Rosselli, M.; Zuckermann, M. Liver cirrhosis. Best Pract. Res. Clin. Gastroenterol 2011, 25, 281–290. [Google Scholar]
- D’Amico, G.; Garcia-Tsao, G.; Pagliaro, L. Natural history and prognostic indicators of survival in cirrhosis A systematic review of 118 studies. J. Hepatol 2006, 44, 217–231. [Google Scholar]
- Iwakiri, Y.; Groszmann, R.J. Vascular endothelial dysfunction in cirrhosis. J. Hepatol 2007, 46, 927–934. [Google Scholar]
- Iwakiri, Y. Endothelial dysfunction in the regulation of cirrhosis and portal hypertension. Liver Int 2012, 32, 199–213. [Google Scholar]
- Wiest, R. Splanchnic and systemic vasodilation: The experimental models. J. Clin. Gastroenterol 2007, 41, S272–S287. [Google Scholar]
- Sarela, A.I.; Mihaimeed, F.M.; Batten, J.J.; Davidson, B.R.; Mathie, R.T. Hepatic and splanchnic nitric oxide activity in patients with cirrhosis. Gut 1999, 44, 749–753. [Google Scholar]
- Wei, C.L.; Khoo, H.E.; Lee, K.H.; Hon, W.M. Differential expression and localization of nitric oxide synthases in cirrhotic livers of bile duct-ligated rats. Nitric Oxide 2002, 7, 91–102. [Google Scholar]
- Mohammed, N.A.; El-Aleem, S.A.; Appleton, I.; Maklouf, M.M.; Said, M.; McMahon, R.F. Expression of nitric oxide synthase isoforms in human liver cirrhosis. J. Pathol 2003, 200, 647–655. [Google Scholar]
- Serna, E.; Mauricio, M.D.; Lluch, P.; Segarra, G.; Cortina, B.; Lluch, S.; Medina, P. Basal release of nitric oxide in the mesenteric artery in portal hypertension and cirrhosis: Role of dimethylarginine dimethylaminohydrolase. J. Gastroenterol. Hepatol 2013, 28, 880–886. [Google Scholar]
- Tain, Y.L.; Chen, C.C.; Lee, C.T.; Kao, Y.H.; Sheen, J.M.; Yu, H.R.; Huang, L.T. Melatonin regulates l-arginine transport and NADPH oxidase in young rats with bile duct ligation: Role of protein kinase C. Pediatr. Res 2013, 73, 395–401. [Google Scholar]
- Chang, K.A.; Lin, I.C.; Sheen, J.M.; Chen, Y.C.; Chen, C.C.; Tain, Y.L.; Hsieh, C.S.; Huang, L.T. Sex differences of oxidative stress to cholestatic liver and kidney injury in young rats. Pediatr. Neonatol 2013, 54, 95–101. [Google Scholar]
- Vizzutti, F.; Romanelli, R.G.; Arena, U.; Rega, L.; Brogi, M.; Calabresi, C.; Masini, E.; Tarquini, R.; Zipoli, M.; Boddi, V.; et al. ADMA correlates with portal pressure in patients with compensated cirrhosis. Eur. J. Clin. Investig 2007, 37, 509–515. [Google Scholar]
- Sciacqua, A.; Perticone, M.; Miceli, S.; Laino, I.; Tassone, E.J.; Grembiale, R.D.; Andreozzi, F.; Sesti, G.; Perticone, F. Endothelial dysfunction and non-alcoholic liver steatosis in hypertensive patients. Nutr. Metab. Cardiovasc. Dis 2011, 21, 485–491. [Google Scholar]
- Child, C.G.; Turcotte, J.G. Surgery and Portal Hypertension. In The Liver and Portal Hypertension; Child, C.G., Ed.; Saunders: Philadelphia, PA, USA, 1964; pp. 50–64. [Google Scholar]
- Eisinger, K.; Krautbauer, S.; Wiest, R.; Karrasch, T.; Hader, Y.; Scherer, M.N.; Farkas, S.; Aslanidis, C.; Buechler, C. Portal vein omentin is increased in patients with liver cirrhosis but is not associated with complications of portal hypertension. Eur. J. Clin. Investig 2013, 43, 926–932. [Google Scholar]
- Malinchoc, M.; Kamath, P.S.; Gordon, F.D.; Peine, C.J.; Rank, J.; ter Borg, P.C. A model to predict poor survival in patients undergoing transjugular intrahepatic portosystemic shunts. Hepatology 2000, 31, 864–871. [Google Scholar]
- Bajaj, J.S.; Ahluwalia, V.; Wade, J.B.; Sanyal, A.J.; White, M.B.; Noble, N.A.; Monteith, P.; Fuchs, M.; Sterling, R.K.; Luketic, V.; et al. Asymmetric dimethylarginine is strongly associated with cognitive dysfunction and brain MR spectroscopic abnormalities in cirrhosis. J. Hepatol 2013, 58, 38–44. [Google Scholar]
- Maddrey, W.C.; Boitnott, J.K.; Bedine, M.S.; Weber, F.L., Jr; Mezey, E.; White, R.I., Jr. Corticosteroid therapy of alcoholic hepatitis. Gastroenterology 1978, 75, 193–199. [Google Scholar]
- Mookerjee, R.P.; Malaki, M.; Davies, N.A.; Hodges, S.J.; Dalton, R.N.; Turner, C.; Sen, S.; Williams, R.; Leiper, J.; Vallance, P.; et al. Increasing dimethylarginine levels are associated with adverse clinical outcome in severe alcoholic hepatitis. Hepatology 2007, 45, 62–71. [Google Scholar]
- Siroen, M.P.; Wiest, R.; Richir, M.C.; Teerlink, T.; Rauwerda, J.A.; Drescher, F.T.; Zorger, N.; van Leeuwen, P.A. Transjugular intrahepatic portosystemic shunt-placement increases arginine/asymmetric dimethylarginine ratio in cirrhotic patients. World J. Gastroenterol 2008, 14, 7214–7219. [Google Scholar]
- Orellana, M.; Rodrigo, R.; Thielemann, L.; Guajardo, V. Bile duct ligation and oxidative stress in the rat: Effects in liver and kidney. Comp. Biochem. Physiol. Toxicol. Pharmacol 2000, 126, 105–111. [Google Scholar]
- Pereira, R.M.; dos Santos, R.A.; Oliveira, E.A.; Leite, V.H.; Dias, F.L.; Rezende, A.S.; Costa, L.P.; Barcelos, L.S.; Teixeira, M.M.; Simoes e Silva, A.C. Development of hepatorenal syndrome in bile duct ligated rats. World J. Gastroenterol 2008, 14, 4505–4511. [Google Scholar]
- Betrosian, A.P.; Agarwal, B.; Douzinas, E.E. Acute renal dysfunction in liver diseases. World J. Gastroenterol 2007, 13, 5552–5559. [Google Scholar]
- Assimakopoulos, S.F.; Vagianos, C.E. Bile duct ligation in rats: A reliable model of hepatorenal syndrome? World J. Gastroenterol 2009, 15, 121–123. [Google Scholar]
- Nijveldt, R.J.; Teerlink, T.; van Guldener, C.; Prins, H.A.; van Lambalgen, A.A.; Stehouwer, C.D.; Rauwerda, J.A.; van Leeuwen, P.A. Handling of asymmetrical dimethylarginine and symmetrical dimethylarginine by the rat kidney under basal conditions and during endotoxaemia. Nephrol. Dial. Transplant 2003, 18, 2542–2550. [Google Scholar]
- Lluch, P.; Mauricio, M.D.; Vila, J.M.; Segarra, G.; Medina, P.; del Olmo, J.A.; Rodrigo, J.M.; Serra, M.A. Accumulation of symmetric dimethylarginine in hepatorenal syndrome. Exp. Biol. Med. 2006, 231, 70–75. [Google Scholar]
- Nicković, V.; Nikolić, J.; Djindjić, N.; Ilić, M.; Nicković, J.; Mladenović, D.; Krstić, N. Diagnostical significance of dimethylarginine in the development of hepatorenal syndrome in patients with alcoholic liver cirrhosis. Vojnosanit. Pregl. 2012, 69, 686–691. [Google Scholar]
- Sesti, G.; Mannino, G.C.; de Lorenzo, C.; Greco, A.; Sciacqua, A.; Marini, M.A.; Andreozzi, F.; Perticone, F. A functional variant of the dimethylarginine dimethylaminohydrolase-2 gene is associated with chronic kidney disease. Atherosclerosis 2013, 231, 141–144. [Google Scholar]
- Andreozzi, F.; Presta, I.; Mannino, G.C.; Scarpelli, D.; di Silvestre, S.; di Pietro, N.; Succurro, E.; Sciacqua, A.; Pandolfi, A.; Consoli, A.; et al. A functional variant of the dimethylarginine dimethylaminohydrolase-2 gene is associated with insulin sensitivity. PLoS One 2012, 7, e36224. [Google Scholar]
- Huang, L.T.; Chen, C.C.; Sheen, J.M.; Chen, Y.J.; Hsieh, C.S.; Tain, Y.L. The interaction between high ammonia diet and bile duct ligation in developing rats: Assessment by spatial memory and asymmetric dimethylarginine. Int. J. Dev. Neurosci 2010, 28, 169–174. [Google Scholar]
- Leke, R.; Oliveira, D.L.; Forgiarini, L.F.; Escobar, T.D.; Hammes, T.O.; Meyer, F.S.; Keiding, S.; Silveira, T.R.; Schousboe, A. Impairment of short term memory in rats with hepatic encephalopathy due to bile duct ligation. Metab. Brain Dis 2013, 28, 187–192. [Google Scholar]
- Dhanda, S.; Kaur, S.; Sandhir, R. Preventive effect of N-acetyl-l-cysteine on oxidative stress and cognitive impairment in hepatic encephalopathy following bile duct ligation. Free Radic. Biol. Med 2013, 56, 204–215. [Google Scholar]
- Javadi-Paydar, M.; Ghiassy, B.; Ebadian, S.; Rahimi, N.; Norouzi, A.; Dehpour, A.R. Nitric oxide mediates the beneficial effect of chronic naltrexone on cholestasis-induced memory impairment in male rats. Behav. Pharmacol 2013, 24, 195–206. [Google Scholar]
- Häussinger, D.; Sies, H. Hepatic encephalopathy: Clinical aspects and pathogenetic concept. Arch. Biochem. Biophys 2013, 536, 97–100. [Google Scholar]
- Córdoba, J.; Mínguez, B. Hepatic encephalopathy. Semin. Liver Dis 2008, 28, 70–80. [Google Scholar]
- Montoliu, C.; Rodrigo, R.; Monfor, P.; Llansola, M.; Cauli, O.; Boix, J.; Elmlili, N.; Agusti, A.; Felipo, V. Cyclic GMP pathways in hepatic encephalopathy Neurological and therapeutic implications. Metab. Brain Dis. 2010, 25, 39–48. [Google Scholar]Int. J. Mol. Sci. 2014, 15, 4005.
- Balasubramaniyan, V.; Wright, G.; Sharma, V.; Davies, N.A.; Sharifi, Y.; Habtesion, A.; Mookerjee, R.P.; Jalan, R. Ammonia reduction with ornithine phenylacetate restores brain eNOS activity via the DDAH-ADMA pathway in bile duct-ligated cirrhotic rats. Am. J. Physiol. Gastrointest. Liver Physiol 2012, 302, G145–G152. [Google Scholar]
- Corbalán, R.; Montoliu, C.; Miñana, M.D.; del Olmo, J.A.; Serra, M.A.; Aparisi, L.; Rodrigo, J.M.; Felipo, V. Altered modulation of soluble guanylatecyclase by nitric oxide in patients with liver disease. Metab. Brain Dis 2002, 17, 295–301. [Google Scholar]
- Boulton, C.L.; Southam, E.; Garthwaite, J. Nitric oxide-dependent long-term potentiation is blocked by a specific inhibitor of soluble guanylylcyclase. Neuroscience 1995, 69, 699–703. [Google Scholar]
- Hawkins, R.D. NO honey, I don’t remember. Neuron 1996, 16, 465–467. [Google Scholar]
- Kielstein, J.T.; Donnerstag, F.; Gasper, S.; Menne, J.; Kielstein, A.; Martens-Lobenhoffer, J.; Scalera, F.; Cooke, J.P.; Fliser, D.; Bode-Böger, S.M. ADMA increases arterial stiffness and decreases cerebral blood flow in humans. Stroke 2006, 37, 2024–2029. [Google Scholar]
- De la Torre, J.C.; Aliev, G. Inhibition of vascular nitric oxide after rat chronic brain hypoperfusion: Spatial memory and immunocytochemical changes. J. Cereb. Blood Flow Metab 2005, 25, 663–672. [Google Scholar]
- Notsu, Y.; Nabika, T.; Bokura, H.; Suyama, Y.; Kobayashi, S.; Yamaguchi, S.; Masuda, J. Evaluation of asymmetric dimethylarginine and homocysteine in microangiopathy-related cerebral damage. Am. J. Hypertens 2009, 22, 257–262. [Google Scholar]
- Mallamaci, F.; Tripepi, G.; Cutrupi, S.; Malatino, L.S.; Zoccali, C. Prognostic value of combined use of biomarkers of inflammation endothelial dysfunction and myocardiopathy in patients with ESRD. Kidney Int 2005, 67, 2330–2337. [Google Scholar]
- Kielstein, J.T.; Kielstein, A. ADMA and the brain: An unfolding story. Am. J. Hypertens. 2009. [Google Scholar] [CrossRef]
- Richir, M.C.; van Lambalgen, A.A.; Teerlink, T.; Wisselink, W.; Bloemena, E.; Prins, H.A.; de Vries, T.P.; van Leeuwen, P.A. Low arginine/asymmetric dimethylarginine ratio deteriorates systemic hemodynamics and organ blood flow in a rat model. Crit. Care Med 2009, 37, 2010–2017. [Google Scholar]
- Perticone, F.; Sciacqua, A.; Maio, R.; Perticone, M.; Maas, R.; Boger, R.H.; Tripepi, G.; Sesti, G.; Zoccali, C. Asymmetric dimethylarginine l-arginine and endothelial dysfunction in essential hypertension. J. Am. Coll. Cardiol. 2005, 46, 518–523. [Google Scholar]
- Sciacqua, A.; Grillo, N.; Quero, M.; Sesti, G.; Perticone, F. Asymmetric dimethylarginine plasma levels and endothelial function in newly diagnosed type 2 diabetic patients. Int. J. Mol. Sci 2012, 13, 13804–13815. [Google Scholar]
- Visser, M.; Vermeulen, M.A.; Richir, M.C.; Teerlink, T.; Houdijk, A.P.; Kostense, P.J.; Wisselink, W.; de Mol, B.A.; van Leeuwen, P.A.; Oudemans-van Straaten, H.M. Imbalance of arginine and asymmetric dimethylarginine is associated with markers of circulatory failure organ failure and mortality in shock patients. Br. J. Nutr. 2012, 107, 1458–1465. [Google Scholar]
- Koch, A.; Weiskirchen, R.; Kunze, J.; Dückers, H.; Bruensing, J.; Buendgens, L.; Matthes, M.; Luedde, T.; Trautwein, C.; Tacke, F. Elevated asymmetric dimethylarginine levels predict short-and long-term mortality risk in critically ill patients. J. Crit. Care 2013, 28, 947–953. [Google Scholar]
- O’Dwyer, M.J.; Dempsey, F.; Crowley, V.; Kelleher, D.P.; McManus, R.; Ryan, T. Septic shock is correlated with asymmetrical dimethyl arginine levels which may be influenced by a polymorphism in the dimethylarginine dimethylaminohydrolase-2 gene: A prospective observational study. Crit. Care 2006, 10, R139:1–R139:7. [Google Scholar]
- Leiper, J.; Murray-Rust, J.; McDonald, N.; Vallance, P. S-Nitrosylation of dimethylarginine dimethylaminohydrolase regulates enzyme activity: Further interactions between nitric oxide synthase and dimethylarginine dimethylaminohydrolase. Proc. Natl. Acad. Sci. USA 2002, 99, 13527–13532. [Google Scholar]
- Liu, H.; Ma, Z.; Lee, S.S. Contribution of nitric oxide to the pathogenesis of cirrhotic cardiomyopathy in bile duct-ligated rats. Gastroenterology 2000, 118, 937–944. [Google Scholar]
- Shafaroodi, H.; Ebrahimi, F.; Moezi, L.; Hashemi, M.; Doostar, Y.; Ghasemi, M.; Dehpour, A.R. Cholestasis induces apoptosis in mice cardiac cells: The possible role of nitric oxide and oxidative stress. Liver Int 2010, 30, 898–905. [Google Scholar]
- Fink, M.P.; Delude, R.L. Epithelial barrier dysfunction: A unifying theme to explain the pathogenesis of multiple organ dysfunction at the cellular level. Crit. Care Clin. 2005, 21, 177–196. [Google Scholar]
- Zhang, J.B.; Du, X.G.; Zhang, H.; Li, M.L.; Xiao, G.; Wu, J.; Gan, H. Breakdown of the gut barrier in patients with multiple organ dysfunction syndrome is attenuated by continuous blood purification: Effects on tight junction structural proteins. Int. J. Artif. Organs 2010, 33, 5–14. [Google Scholar]
- Assimakopoulos, S.F.; Vagianos, C.E.; Patsoukis, N.; Georgiou, C.; Nikolopoulou, V.; Scopa, C.D. Evidence for intestinal oxidative stress in obstructive jaundice-induced gut barrier dysfunction in rats. Acta Physiol. Scand. 2004, 180, 177–185. [Google Scholar]
- Assimakopoulos, S.F.; Scopa, C.D.; Zervoudakis, G.; Mylonas, P.G.; Georgiou, C.; Nikolopoulou, V.; Vagianos, C.E. Bombesin and neurotensin reduce endotoxemia intestinal oxidative stress and apoptosis in experimental obstructive jaundice. Ann. Surg 2005, 241, 159–167. [Google Scholar]
- Assimakopoulos, S.F.; Thomopoulos, K.C.; Patsoukis, N.; Georgiou, C.D.; Scopa, C.D.; Nikolopoulou, V.N.; Vagianos, C.E. Evidence for intestinal oxidative stress in patients with obstructive jaundice. Eur. J. Clin. Investig 2006, 36, 181–187. [Google Scholar]
- Assimakopoulos, S.F.; Tsamandas, A.C.; Tsiaoussis, G.I.; Karatza, E.; Zisimopoulos, D.; Maroulis, I.; Kontogeorgou, E.; Georgiou, C.D.; Scopa, C.D.; Thomopoulos, K.C. Intestinal mucosal proliferation apoptosis and oxidative stress in patients with liver cirrhosis. Ann. Hepatol 2013, 12, 301–307. [Google Scholar]
- Hong, J.Y.F.; Sato, E.; Hiramoto, K.; Nishikawa, M.; Inoue, M. Mechanism of liver injury during obstructive jaundice: Role of nitric oxide splenic cytokines and intestinal flora. J. Clin. Biochem. Nutr. 2007, 40, 184–193. [Google Scholar]
- Hong, J.Y.F.; Sato, E.; Nishikawa, T.; Hiramoto, K.; Inoue, M. Effect of obstructive jaundice and nitric oxide on the profiles of intestinal bacterial flora in wild and iNOS mice. J. Clin. Biochem. Nutr. 2009, 44, 160–167. [Google Scholar]

© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Sheen, J.-M.; Chen, Y.-C.; Tain, Y.-L.; Huang, L.-T. Increased Circulatory Asymmetric Dimethylarginine and Multiple Organ Failure: Bile Duct Ligation in Rat as a Model. Int. J. Mol. Sci. 2014, 15, 3989-4006. https://doi.org/10.3390/ijms15033989
Sheen J-M, Chen Y-C, Tain Y-L, Huang L-T. Increased Circulatory Asymmetric Dimethylarginine and Multiple Organ Failure: Bile Duct Ligation in Rat as a Model. International Journal of Molecular Sciences. 2014; 15(3):3989-4006. https://doi.org/10.3390/ijms15033989
Chicago/Turabian StyleSheen, Jiunn-Ming, Yu-Chieh Chen, You-Lin Tain, and Li-Tung Huang. 2014. "Increased Circulatory Asymmetric Dimethylarginine and Multiple Organ Failure: Bile Duct Ligation in Rat as a Model" International Journal of Molecular Sciences 15, no. 3: 3989-4006. https://doi.org/10.3390/ijms15033989