Proteomic Analysis of the Follicular Fluid of Tianzhu White Yak during Diestrus

Abstract
:1. Introduction
2. Results and Discussion
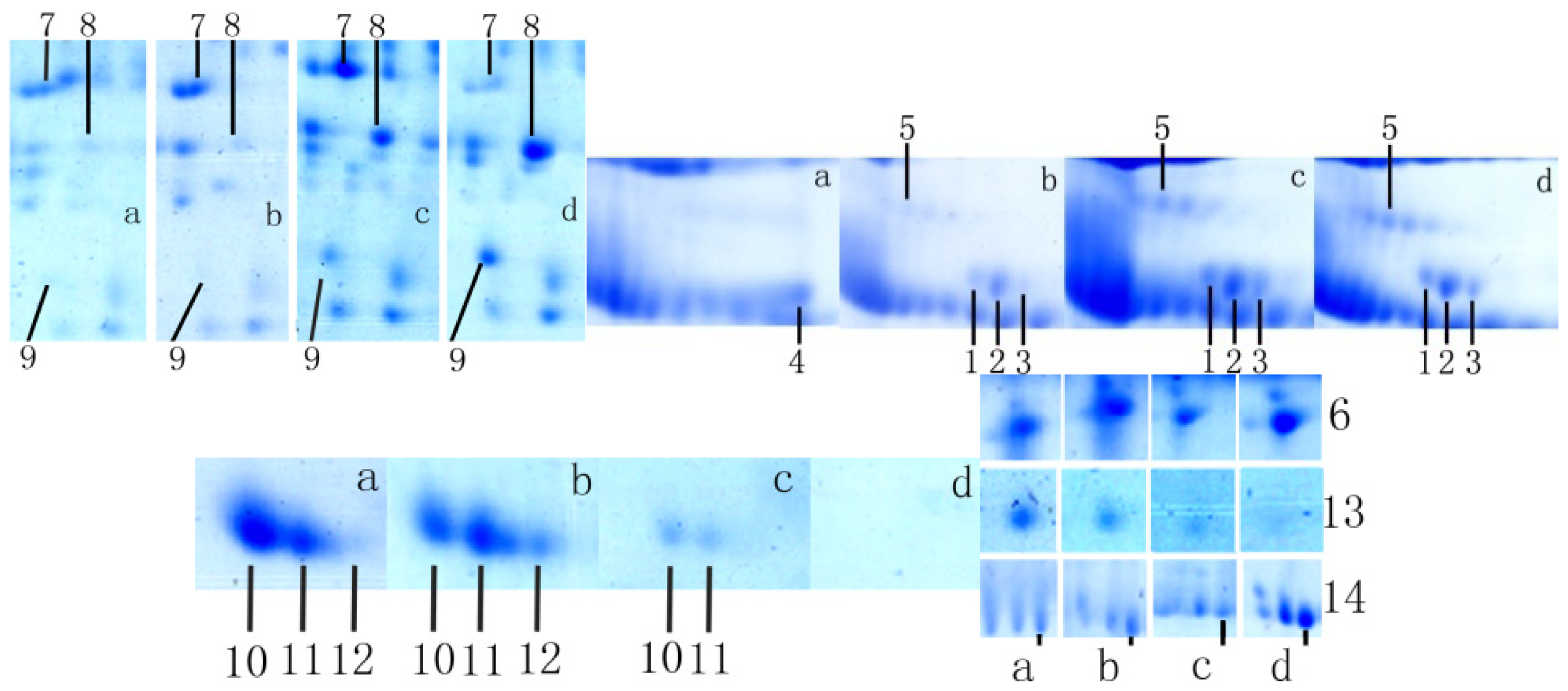
2.1. Analysis of Differentially Expressed Protein Spots
2.2. Identification and Analysis of Differentially Expressed Proteins
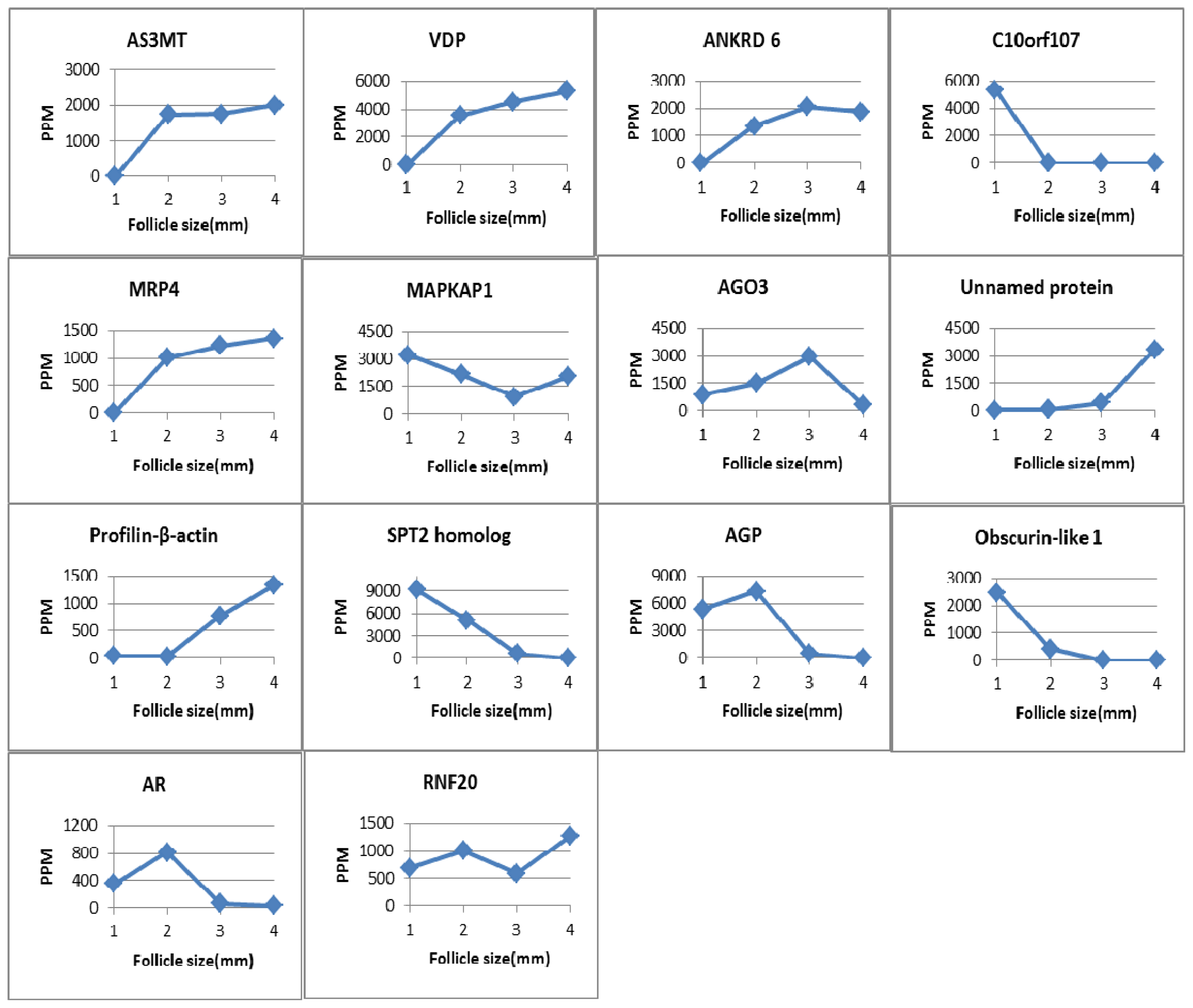
2.2.1. Arsenic (+3) Methyltransferase
2.2.2. Vitamin D-Binding Protein
2.2.3. Mitogen Activated Protein Kinase 2-Associated Protein-1
2.2.4. Argonaute-3
2.2.5. Aldose Reductase (AR)
2.2.6. E3 Ubiquitin-Protein Ligase BRE1A (RNF20)
3. Materials and Methods
3.1. Collection of Follicular Fluid
3.2. Two-Dimensional Gel Electrophoresis
3.3. Staining and Image Analysis
3.4. Peptide Identification
3.5. Statistical Analysis
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Yu, S.J.; Ju, X.H.; Wang, L.B.; Fan, J.F. Successful embryo transfer in tianzhu white yak using standard protocol. Sci. China Ser. C 2007, 50, 655–659. [Google Scholar]
- Zhang, C.J.; Ma, F.L.; Li, L.; Wang, W.F. Investigation on polymorphism of blood proteins in huzhu white yaks. Qinghai J. Anim. Sci. Vet. Med. 1996, 1, 14–16. [Google Scholar]
- Wang, S.L.; Fan, J.F.; Li, T.Q.; Yu, S.J. Ovarian change of the white yak during superovulation using b-mode ultrasonic scanner. J. Gansu Agric. Univ. 2008, 3, 25–28. [Google Scholar]
- Huo, S.D.; Chen, A.; Long, L.; Ju, X.H.; Yu, S.J. Characterization of blood biochemical parameters in tianzhu white yak. J. Northwest Univ. Natl. (Nat. Sci.) 2008, 3, 46–48. [Google Scholar]
- Yan, P.; Guo, X.; Xu, B.Z.; En, R.; Zen, Y.F. White yak oocyte maturation in vitro. China Herbiv. 2006, 4, 7–9. [Google Scholar]
- Zhang, J.; Yu, S.J. Investigation with breeding and reproduction body condition of Yak at the plateau. Anim. Husb. Vet. Med. 2005, 8, 21–22. [Google Scholar]
- Yang, Z.M. Reproductive Biology; Science Press: Beijing, China, 2005. [Google Scholar]
- Bao, B.; Garverick, H.A. Expression of steroidogenic enzyme and gonadotropin receptor genes in bovine follicles during ovarian follicular waves: A review. J. Anim. Sci. 1998, 76, 1903–1921. [Google Scholar]
- Leibfried-Rutledge, M.; Critser, E.; Parrish, J.; First, N. In vitro maturation and fertilization of bovine oocytes. Theriogenology 1989, 31, 61–74. [Google Scholar]
- Zhao, X.X. Veterinary Obstetrics, 4th ed; China Agriculture Press: Beijing, China, 2009. [Google Scholar]
- Revelli, A.; Delle Piane, L.; Casano, S.; Molinari, E.; Massobrio, M.; Rinaudo, P. Follicular fluid content and oocyte quality: From single biochemical markers to metabolomics. Reprod. Biol. Endocrinol. 2009, 7, 4330–4337. [Google Scholar]
- Fahiminiya, S.; Labas, V.; Roche, S.; Dacheux, J.-L.G.; érard, N. Proteomic analysis of mare follicular fluid during late follicle development. Proteome Sci. 2011, 9, 1–19. [Google Scholar]
- Richards, J.S. Hormonal control of gene expression in the ovary. Endocr. Rev. 1994, 15, 725–751. [Google Scholar]
- Mayer, D.R.; Kosmus, W.; Pogglitsch, H.; Beyer, W. Essential trace elements in humans. Biol. Trace Elem. Res. 1993, 37, 27–38. [Google Scholar]
- Chatterjee, A.; Chatterji, U. Arsenic abrogates the estrogen-signaling pathway in the rat uterus. Reprod. Biol. Endocrinol. 2010, 8, 80. [Google Scholar]
- Chiou, T.J.; Chu, S.-T.; Tzeng, W.-F.; Huang, Y.-C.; Liao, C.-J. Arsenic trioxide impairs spermatogenesis via reducing gene expression levels in testosterone synthesis pathway. Chem. Res. Toxicol. 2008, 21, 1562–1569. [Google Scholar]
- Lin, G.Z.; Fan, J.F.; Yu, S.J. Morphological observation of ovary and ovarian follicle of pregnant yak. J. Gansu Agric. Univ. 2006, 1, 1–4. [Google Scholar]
- Fan, J.F.; Wang, S.L.; Li, T.Q.; Xu, G.Q.; Wang, L.B.; Yu, S.J. Preliminary research on the follicular atresia during super ovulation in tianzhu white yak. J. Anhui Agric. Sci. 2008, 20, 8612–8614. [Google Scholar]
- Anifandis, G.M.; Dafopoulos, K.; Messini, C.I.; Chalvatzas, N.; Liakos, N.; Pournaras, S.; Messinis, I.E. Prognostic value of follicular fluid 25-OH vitamin d and glucose levels in the IVF outcome. Reprod. Biol. Endocrinol. 2010, 8, 91–95. [Google Scholar]
- Estes, S.J.; Ye, B.; Qiu, W.; Cramer, D.; Hornstein, M.D.; Missmer, S.A. A proteomic analysis of ivf follicular fluid in women <or= 32 years old. Fertil. Steril. 2009, 92, 1569–1578. [Google Scholar]
- Ozkan, S.; Jindal, S.; Greenseid, K.; Shu, J.; Zeitlian, G.; Hickmon, C.; Pal, L. Replete vitamin d stores predict reproductive success following in vitro fertilization. Fertil. Steril. 2010, 94, 1314–1319. [Google Scholar]
- Potashnik, G.; Lunenfeld, E.; Levitas, E.; Itskovitz, J.; Albutiano, S.; Yankowitz, N.; Sonin, Y.; Levy, J.; Glezerman, M.; Shany, S. The relationship between endogenous oestradiol and vitamin d3 metabolites in serum and follicular fluid during ovarian stimulation for in vitro fertilization and embryo transfer. Hum. Reprod. 1992, 7, 1357–1360. [Google Scholar]
- Nielsen, H.K.; Brixen, K.; Bouillon, R.; Mosekilde, L. Changes in biochemical markers of osteoblastic activity during the menstrual cycle. J. Clin. Endocrinol. Metab. 1990, 70, 1431–1437. [Google Scholar]
- Kinuta, K.; Tanaka, H.; Moriwake, T.; Aya, K.; Kato, S.; Seino, Y. Vitamin d is an important factor in estrogen biosynthesis of both female and male gonads. Endocrinology 2000, 141, 1317–1324. [Google Scholar]
- Brain, H.; Bano, G.; Brincat, M.; Galea, R.; Pellatt, L.; Nussey, S.; Mason, H. 125-dihydroxyvitamin d3 has a direct effect on steroid production from human theca cells. Endocri. Abstr. 2003, 5, 211–217. [Google Scholar]
- Jacinto, E.; Facchinetti, V.; Liu, D.; Soto, N.; Wei, S.; Jung, S.Y.; Huang, Q.; Qin, J.; Su, B. Sin1/mip1 maintains rictor-mtor complex integrity and regulates akt phosphorylation and substrate specificity. Cell 2006, 127, 125–138. [Google Scholar]
- Schroder, W.; Cloonan, N.; Bushell, G.; Sculley, T. Alternative polyadenylation and splicing of mrnas transcribed from the human sin1 gene. Gene 2004, 339, 17–23. [Google Scholar]
- Cavaliere, V.; Donati, A.; Hsouna, A.; Hsu, T.; Gargiulo, G. Dakt kinase controls follicle cell size during drosophila oogenesis. Dev. Dyn. 2005, 232, 845–854. [Google Scholar]
- Yu, J.; Yaba, A.; Kasiman, C.; Thomson, T.; Johnson, J. mTor controls ovarian follicle growth by regulating granulosa cell proliferation. PLoS One 2011, 6, e21415. [Google Scholar]
- Hutvagner, G.; Simard, M.J. Argonaute proteins: Key players in RNA silencing. Nat. Rev. Mol. Cell Biol. 2008, 9, 22–32. [Google Scholar]
- Reuter, M.; Chuma, S.; Tanaka, T.; Franz, T.; Stark, A.; Pillai, R.S. Loss of the mili-interacting tudor domain–containing protein-1 activates transposons and alters the mili-associated small rna profile. Nat. Struct. Mol. Biol. 2009, 16, 639–646. [Google Scholar]
- Carmell, M.A.; Xuan, Z.; Zhang, M.Q.; Hannon, G.J. The argonaute family: Tentacles that reach into rnai developmental control stem cell maintenance and tumorigenesis. Genes Dev. 2002, 16, 2733–2742. [Google Scholar]
- Brennecke, J.; Aravin, A.A.; Stark, A.; Dus, M.; Kellis, M.; Sachidanandam, R.; Hannon, G.J. Discrete small rna-generating loci as master regulators of transposon activity in drosophila. Cell 2007, 128, 1089–1103. [Google Scholar]
- Azzam, G.; Smibert, P.; Lai, E.C.; Liu, J.L. Drosophila argonaute 1 and its mirna biogenesis partners are required for oocyte formation and germline cell division. Dev. Biol. 2012, 365, 384–394. [Google Scholar]
- Colton, S.A.; Downs, S.M. Potential role for the sorbitol pathway in the meiotic dysfunction exhibited by oocytes from diabetic mice. J. Exp. Zool. A 2004, 301, 439–448. [Google Scholar]
- Ramana, K.V.; Reddy, A.; Tammali, R.; Srivastava, S.K. Aldose reductase mediates endotoxin-induced production of nitric oxide and cytotoxicity in murine macrophages. Free Radic. Biol. Med. 2007, 42, 1290–1302. [Google Scholar]
- Iwata, N.; Hara, S.; Nishimura, C.; Takahashi, M.; Mukaii, T.; Takayama, M.; Endo, T. Hormonal regulation of aldose reductase in rat ovary during the estrous cycle. Eur. J. Biochem. 1996, 235, 444–448. [Google Scholar]
- Bhojwani, M.; Rudolph, E.; Kanitz, W.; Zuehlke, H.; Schneider, F.; Tomek, W. Molecular analysis of maturation processes by protein and phosphoprotein profiling during in vitro maturation of bovine oocytes: A proteomic approach. Cloning Stem Cells 2006, 8, 259–274. [Google Scholar]
- Jääskeläinen, T.; Makkonen, H.; Visakorpi, T.; Kim, J.; Roeder, R.G.; Palvimo, J.J. Histone h2b ubiquitin ligases rnf20 and rnf40 in androgen signaling and prostate cancer cell growth. Mol. Cell. Endocrinol. 2012, 350, 87–98. [Google Scholar]
- Dai, A.; Yan, G.; He, Q.; Jiang, Y.; Zhang, Q.; Fang, T.; Ding, L.; Sun, J.; Sun, H.; Hu, Y. Orphan nuclear receptor nur77 regulates androgen receptor gene expression in mouse ovary. PLoS One 2012, 7, e39950. [Google Scholar]
- Bradford, M.M. Arapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar]
- Görg, A.; Obermaier, C.; Boguth, G.; Weiss, W. Recent developments in two-dimensional gel electrophoresis with immobilized ph gradients: Wide ph gradients up to ph 12 longer separation distances and simplified procedures. Electrophoresis 1999, 20, 712–717. [Google Scholar]



{kind=link}
{kind=link}
| No. | (NCBI) Accession | Protein Name | PI/MW (kDa) | Peptide No. | Coverage (%) | MASCOT Score |
|---|---|---|---|---|---|---|
| 1 | gi|78042514 | Arsenic (+3) methyltransferase [Bos taurus] | 5.36/42.51 | 7 | 26 | 65 |
| 2 | gi|85701291 | Vitamin D-binding protein [Bos taurus] | 5.36/54.90 | 16 | 34 | 102 |
| 3 | NP_001192406.1 | Ankyrin repeat domain-containing protein 6 [Bos taurus] | 9.42/80.17 | 4 | 7 | 35 |
| 4 | gi|157278953 | C10orf107 protein [Bos taurus] | 6.92/21.19 | 5 | 4 | 50 |
| 5 | XP_001256333.2 | Multidrug resistance-associated protein 4-like [Bos taurus] | 9.53/22.29 | 3 | 20 | 35 |
| 6 | gi|187611460 | Mitogen activated protein kinase 2-associated protein-1 | 7.24/59.54 | 4 | 12 | 66 |
| 7 | gi|229544700 | Protein argonaute-3 [Bos taurus] | 9.22/98.38 | 8 | 15 | 66 |
| 8 | gi|21538351 | Unnamed protein [Bos taurus] | 6.36/34.24 | 11 | 41 | 79 |
| 9 | gi|157881403 | Crystalline profilin-beta-actin [Bos taurus] | 5.29/41.92 | 9 | 29 | 63 |
| 10 | gi|329664588 | Protein SPT2 homolog [Bos taurus] | 9.79/75.44 | 8 | 11 | 48 |
| 11 | gi|121957959 | Alpha-1-acid glycoprotein | 5.62/23.45 | 7 | 35 | 65 |
| 12 | gi|111305443 | Obscurin-like-1 [Bos taurus] | 5.15/68.96 | 7 | 13 | 67 |
| 13 | gi|162652 | Aldose reductase (EC 1.1.1.21), partial [Bos taurus] | 5.68/34.35 | 6 | 19 | 72 |
| 14 | gi|182627578 | E3 ubiquitin-protein ligase BRE1A [Bos taurus] | 5.70/114.27 | 11 | 14 | 64 |
© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Tao, J.; Zhao, G.; Zhao, X.; Li, F.; Wu, X.; Hu, J.; Zhang, Y. Proteomic Analysis of the Follicular Fluid of Tianzhu White Yak during Diestrus. Int. J. Mol. Sci. 2014, 15, 4481-4491. https://doi.org/10.3390/ijms15034481
Tao J, Zhao G, Zhao X, Li F, Wu X, Hu J, Zhang Y. Proteomic Analysis of the Follicular Fluid of Tianzhu White Yak during Diestrus. International Journal of Molecular Sciences. 2014; 15(3):4481-4491. https://doi.org/10.3390/ijms15034481
Chicago/Turabian StyleTao, Jinzhong, Guoshun Zhao, Xingxu Zhao, Fadi Li, Xiaohu Wu, Junjie Hu, and Yong Zhang. 2014. "Proteomic Analysis of the Follicular Fluid of Tianzhu White Yak during Diestrus" International Journal of Molecular Sciences 15, no. 3: 4481-4491. https://doi.org/10.3390/ijms15034481
APA StyleTao, J., Zhao, G., Zhao, X., Li, F., Wu, X., Hu, J., & Zhang, Y. (2014). Proteomic Analysis of the Follicular Fluid of Tianzhu White Yak during Diestrus. International Journal of Molecular Sciences, 15(3), 4481-4491. https://doi.org/10.3390/ijms15034481