A Preliminary X-ray Study of Murine Tnfaip8/Oxi-α

Abstract
:1. Introduction
2. Results and Discussion
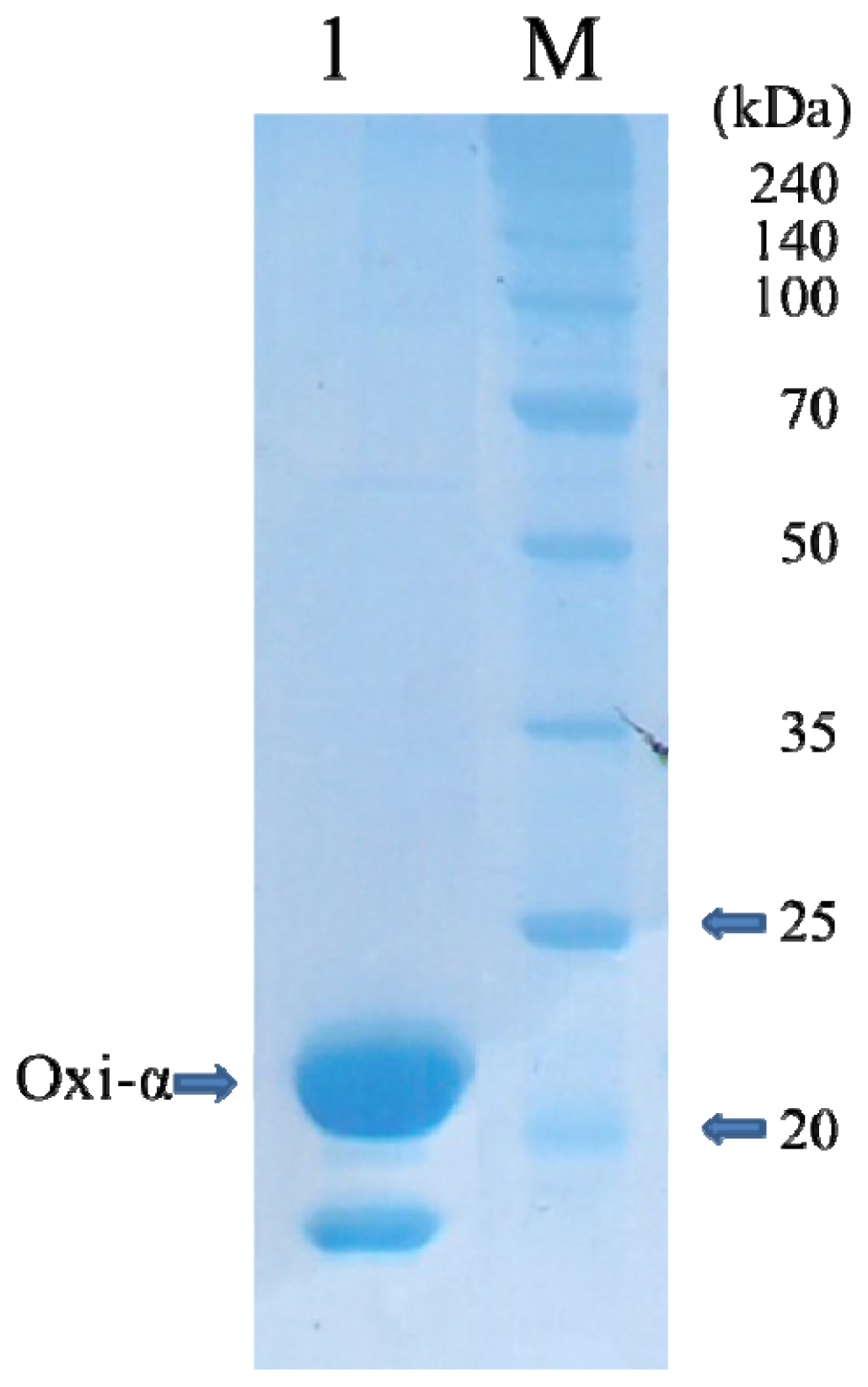
2.1. Protein Expression and Purification
2.2. Protein Crystallization
2.3. X-ray Diffraction
3. Experimental Section
3.1. Expression and Purification of Tnfaip8/Oxi-α
3.2. Growth of Cultures
3.3. Protein Purification
3.4. Site-Directed Mutagenesis
3.5. Crystallization of Mutant Tnfaip8/Oxi-α
3.6. Data Collection and Reduction
4. Conclusions
Acknowledgments
Conflicts of Interest
- Author ContributionsDaeun Lee and Mi-Sun Kim, crystallographic studies and manuscript preparation; Jimin Park, biochemical studies; Gil-Ja Jhon and Jin H. Son, project supervisor, manuscript preparation; Dong Hae Shin, project supervisor, manuscript preparation, principal manuscript author.
References
- McGeer, P.L.; Itagaki, S.; Boyes, B.E.; McGeer, E.G. Reactive microglia are positive for HLA-DR in the substantia nigra of Parkinson’s and Alzheimer’s disease brains. Neurology 1988, 38, 1285–1291. [Google Scholar]
- Hwang, O. Role of oxidative stress in Parkinson’s disease. Exp. Neurobiol. 2013, 22, 11–17. [Google Scholar]
- Yoo, M.S.; Kawamata, H.; Kim, D.J.; Chun, H.S.; Son, J.H. Experimental strategy to identify genes susceptible to oxidative stress in nigral dopaminergic neurons. Neurochem. Res. 2004, 29, 1223–1234. [Google Scholar]
- Choi, K.C.; Kim, S.H.; Ha, J.Y.; Kim, S.T.; Son, J.H. A novel mTOR activating protein protects dopamine neurons against oxidative stress by repressing autophagy related cell death. J. Neurochem. 2010, 112, 366–376. [Google Scholar]
- Zhang, X.; Wang, J.; Fan, C.; Li, H.; Sun, H.; Gong, S.; Chen, Y.H.; Shi, Y. Crystal structure of TIPE2 provides insights into immune homeostasis. Nat. Struct. Mol. Biol. 2009, 16, 89–90. [Google Scholar]
- Andreeva, A.; Howorth, D.; Chandonia, J.M.; Brenner, S.E.; Hubbard, T.J.; Chothia, C.; Murzin, A.G. Data growth and its impact on the SCOP database: New developments. Nucleic Acids Res. 2008, 36, D419–D425. [Google Scholar]
- Sun, H.; Gong, S.; Carmody, R.J.; Hilliard, A.; Li, L.; Sun, J.; Kong, L.; Xu, L.; Hilliard, B.; Hu, S.; et al. TIPE2 a negative regulator of innate and adaptive immunity that maintains immune homeostasis. Cell 2008, 133, 415–426. [Google Scholar]
- Kim, M.S.; Jeong, J.; Jeong, J.; Shin, D.H.; Lee, K.J. Structure of Nm23-H1 under oxidative conditions. Acta Cryst. 2013, D69, 669–680. [Google Scholar]
- Matthews, B.W. Solvent content of protein crystals. J. Mol. Biol. 1968, 33, 491–497. [Google Scholar]
- McCoy, A.J.; Grosse-Kunstleve, R.W.; Adams, P.D.; Winn, M.D.; Storoni, L.C.; Read, R.J. Phaser crystallographic software. J. Appl. Cryst. 2007, 40, 658–674. [Google Scholar]
- Adams, P.D.; Afonine, P.V.; Bunkóczi, G.; Chen, V.B.; Davis, I.W.; Echols, N.; Headd, J.J.; Hung, L.W.; Kapral, G.J.; Grosse-Kunstleve, R.W.; et al. PHENIX: A comprehensive Python-based system for macromolecular structure solution. Acta Cryst. 2010, D66, 213–221. [Google Scholar]
- Terwilliger, T.C.; Read, R.J.; Adams, P.D.; Brünger, A.T.; Afonine, P.V.; Hung, L.W. Model morphing and sequence assignment after molecular replacement. Acta Cryst. 2013, D69, 2244–2250. [Google Scholar]
- Terwilliger, T.C.; Grosse-Kunstleve, R.W.; Afonine, P.V.; Moriarty, N.W.; Zwart, P.H.; Hung, L.W.; Read, R.J.; Adams, P.D. Iterative model building structure refinement and density modification with the PHENIX AutoBuild wizard. Acta Cryst. 2008, D64, 61–69. [Google Scholar]
- Kim, S.H.; Shin, D.H.; Liu, J.; Oganesyan, V.; Chen, S.; Xu, Q.S.; Kim, J.S.; Das, D.; Schulze-Gahmen, U.; Holbrook, S.R.; et al. Structural genomics of minimal organisms and protein fold space. J. Struct. Funct. Genomics 2005, 6, 63–70. [Google Scholar]
- Kim, R.; Sandler, S.J.; Goldman, S.; Yokota, H.; Clark, A.J.; Kim, S.-H. Overexpression of archaeal proteins in Escherichia coli. Biotechnol. Lett. 1998, 20, 207–210. [Google Scholar]
- Otwinowski, Z.; Minor, W. Processing of X-ray diffraction data collected in oscillation mode. Methods Enzymol. 1997, 276, 307–326. [Google Scholar]


{kind=link}
{kind=link}
| Family members | Tnfaip8/Oxi-α | Tnfaip8l1/Oxi-β | Tnfaip8l2/Oxi-γ | Tnfaip8l3/Oxi-δ |
|---|---|---|---|---|
| Sequence identity with Tnfaip8/Oxi-α | 100% | 54.4% | 53.9% | 54.4% |
| X-ray source | Pohang Accelerator Laboratory (PAL) |
|---|---|
| X-ray wavelength (Å) | 0.97933 |
| Temperature (K) | 100 |
| Space group | P21212 |
| Unit cell parameter | |
| -a (Å) | 66.9 |
| -b (Å) | 72.3 |
| -c (Å) | 93.5 |
| Volume fraction of solvent (%) | 47.1 |
| Vm (Å3/Dalton) | 2.33 |
| Resolution range (Å) | 50.0–1.8 (1.83–1.80) |
| Total number of observed reflections | 181684 |
| Multiplicity | 4.4 (4.5) |
| Unique reflections | 41052 (2090) |
| Rsym † (%) | 7.7 (46.1) |
| Data completeness (%) | 95.7 (99.1) |
| Average I/σ (I) | 16.4 (2.9) |
© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Lee, D.; Kim, M.-S.; Park, J.; Jhon, G.-J.; Son, J.H.; Shin, D.H. A Preliminary X-ray Study of Murine Tnfaip8/Oxi-α. Int. J. Mol. Sci. 2014, 15, 4523-4530. https://doi.org/10.3390/ijms15034523
Lee D, Kim M-S, Park J, Jhon G-J, Son JH, Shin DH. A Preliminary X-ray Study of Murine Tnfaip8/Oxi-α. International Journal of Molecular Sciences. 2014; 15(3):4523-4530. https://doi.org/10.3390/ijms15034523
Chicago/Turabian StyleLee, Daeun, Mi-Sun Kim, Jimin Park, Gil-Ja Jhon, Jin H. Son, and Dong Hae Shin. 2014. "A Preliminary X-ray Study of Murine Tnfaip8/Oxi-α" International Journal of Molecular Sciences 15, no. 3: 4523-4530. https://doi.org/10.3390/ijms15034523