Effects of Lipoic Acid on Immune Function, the Antioxidant Defense System, and Inflammation-Related Genes Expression of Broiler Chickens Fed Aflatoxin Contaminated Diets


 ,
, 
Abstract
:1. Introduction
2. Results
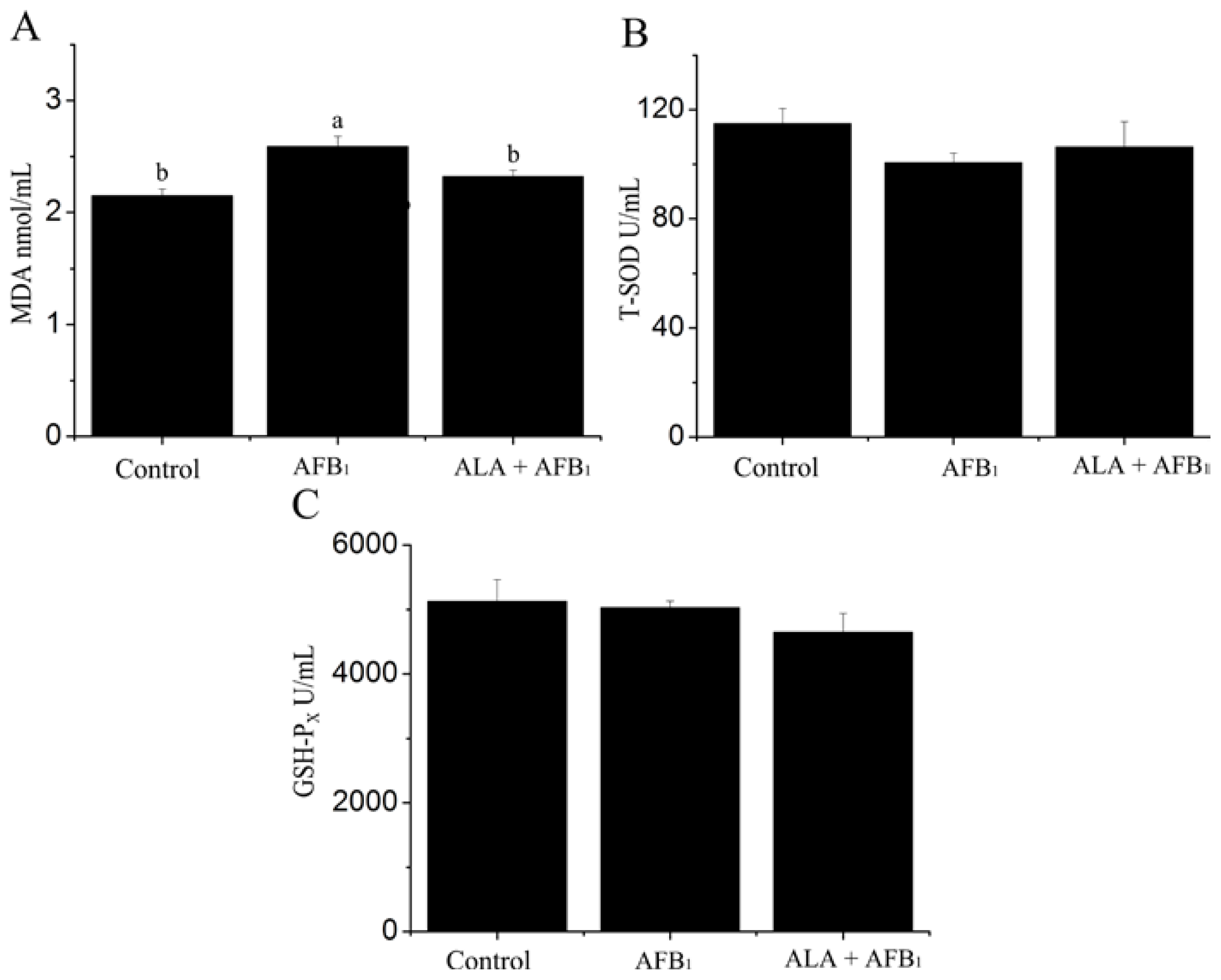
2.1. Effect on Serum Oxidant and Antioxidant Status
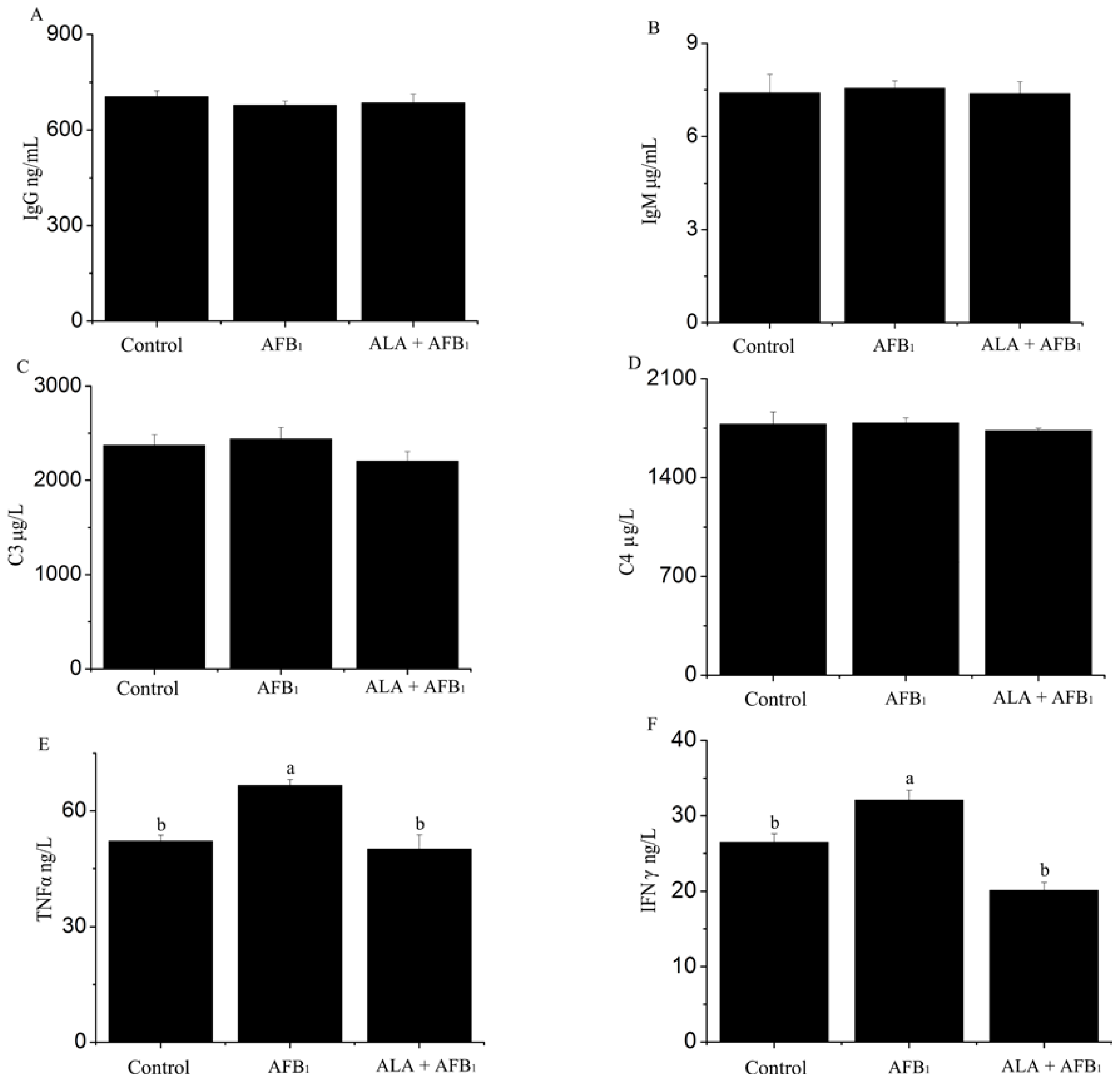
2.2. Effect on Serum Immune Response
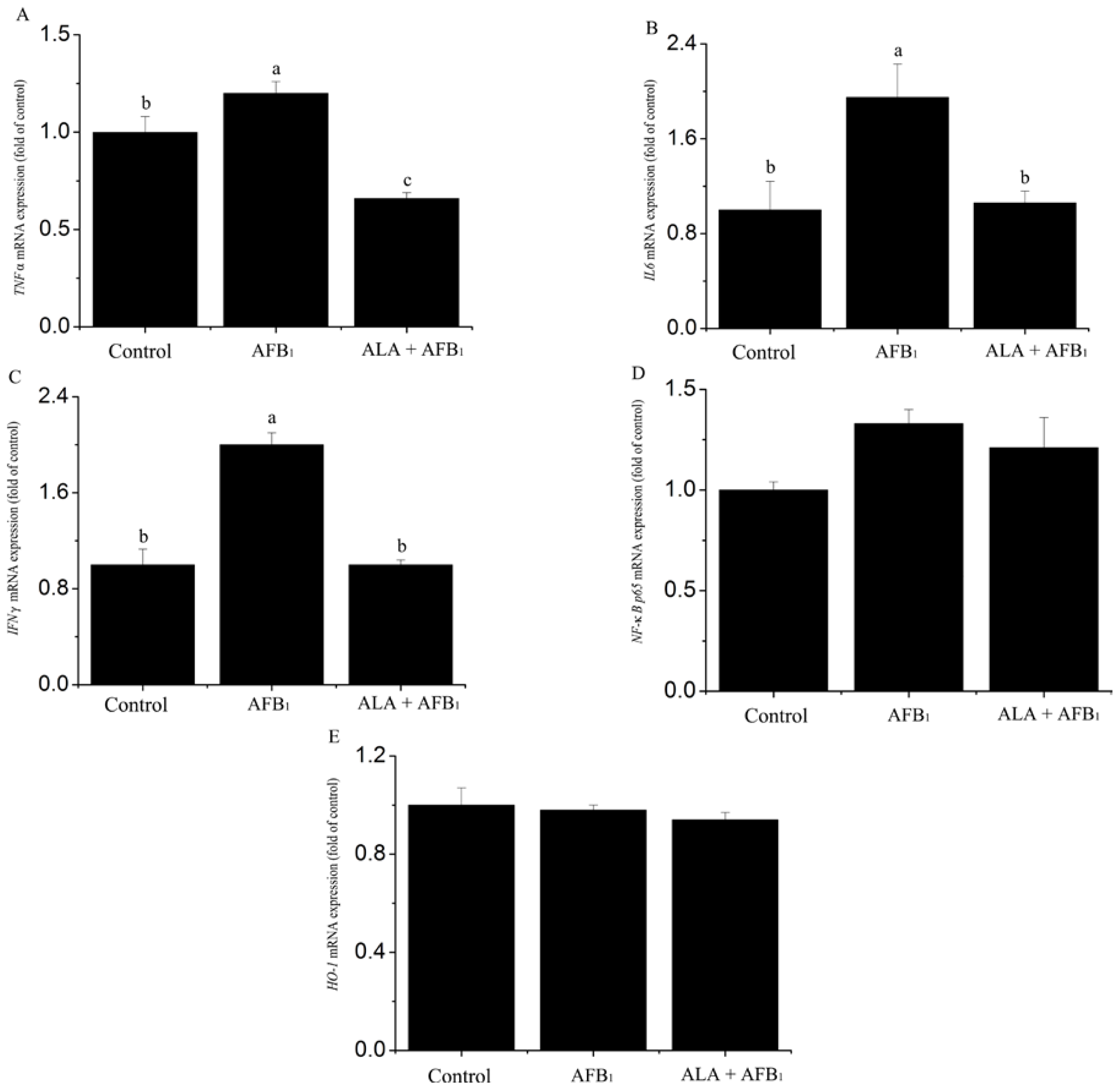
2.3. Effect on Gene mRNA Expression
3. Discussion
4. Experimental Section
4.1. Collection of Feed Ingredients Contaminated with Aflatoxin B1
4.2. Animals
4.3. Experimental Design
4.4. Parameter Analysis
4.5. Gene Expression Analyses
4.6. Statistical Analyses
5. Conclusions
Acknowledgments
Conflicts of Interest
- Author ContributionsY.L., Q.-G.M. and C.J. designed the experiments. Y.L., L.-H.Z. and G.-X.D. performed the experiments. Y.L. analyzed the data. Y.L., L.-H.Z., G.-X.D., Q.-G.M. and C.J. contributed reagents/materials/analysis tools. Y.L., H.W. and C.J. wrote the paper.
References
- Bennett, J.W.; Klich, M. Mycotoxins. Clin. Microbiol. Rev 2003, 16, 497–516. [Google Scholar]
- Wogan, G.N. Aflatoxin carcinogenesis. In Methods in Cancer Research; Bush, H., Ed.; Academic Press: New York, NY, USA, 1973; pp. 309–344. [Google Scholar]
- Meissonnier, G.M.; Pinton, P.; Laffitte, J.; Cossalter, A.-M.; Gong, Y.Y.; Wild, C.P.; Bertin, G.; Galtier, P.; Oswald, I.P. Immunotoxicity of aflatoxin B1: Impairment of the cell-mediated response to vaccine antigen and modulation of cytokine expression. Toxicol. Appl. Pharmacol 2008, 231, 142–149. [Google Scholar]
- Richard, J.L. Some major mycotoxins and their mycotoxicoses—An overview. Int. J. Food Microbiol 2007, 119, 3–10. [Google Scholar]
- Mary, V.S.; Theumer, M.G.; Arias, S.L.; Rubinstein, H.R. Reactive oxygen species sources and biomolecular oxidative damage induced by aflatoxin B1 and fumonisin b1 in rat spleen mononuclear cells. Toxicology 2012, 302, 299–307. [Google Scholar]
- Wagacha, J.; Muthomi, J. Mycotoxin problem in Africa: Current status, implications to food safety and health and possible management strategies. Int. J. Food Microbiol 2008, 124, 1–12. [Google Scholar]
- Ames, B.N. Dietary carcinogens and anticarcinogens oxygen radicals and degenerative diseases. Science 1983, 221, 1256–1264. [Google Scholar]
- Packer, L.; Witt, E.H.; Tritschler, H.J. Alpha-lipoic acid as a biological antioxidant. Free Radic. Biol. Med 1995, 19, 227–250. [Google Scholar]
- Shay, K.P.; Moreau, R.F.; Smith, E.J.; Smith, A.R.; Hagen, T.M. Alpha-lipoic acid as a dietary supplement: Molecular mechanisms and therapeutic potential. Biochim. Biophys. Acta 2009, 1790, 1149–1160. [Google Scholar]
- Wong, A.; Dukic-Stefanovic, S.; Gasic-Milenkovic, J.; Schinzel, R.; Wiesinger, H.; Riederer, P.; Münch, G. Anti-inflammatory antioxidants attenuate the expression of inducible nitric oxide synthase mediated by advanced glycation endproducts in murine microglia. Eur. J. Neurosci 2001, 14, 1961–1967. [Google Scholar]
- Sola, S.; Mir, M.Q.; Cheema, F.A.; Khan-Merchant, N.; Menon, R.G.; Parthasarathy, S.; Khan, B.V. Irbesartan and lipoic acid improve endothelial function and reduce markers of inflammation in the metabolic syndrome results of the irbesartan and lipoic acid in endothelial dysfunction (island) study. Circulation 2005, 111, 343–348. [Google Scholar]
- Miazzo, R.; Peralta, M.F.; Magnoli, C.; Salvano, M.; Ferrero, S.; Chiacchiera, S.M.; Carvalho, E.C.; Rosa, C.A.; Dalcero, A. Efficacy of sodium bentonite as a detoxifier of broiler feed contaminated with aflatoxin and fumonisin. Poult. Sci 2005, 84, 1–8. [Google Scholar]
- Modirsanei, M.; Mansoori, B.; Khosravi, A.R.; Kiaei, M.M.; Khazraeinia, P.; Farkhoy, M.; Masoumi, Z. Effect of diatomaceous earth on the performance and blood variables of broiler chicks during experimental aflatoxicosis. J. Sci. Food Agric 2008, 88, 626–632. [Google Scholar]
- Chaytor, A.C.; See, M.T.; Hansen, J.A.; de Souza, A.L.; Middleton, T.F.; Kim, S.W. Effects of chronic exposure of diets with reduced concentrations of aflatoxin and deoxynivalenol on growth and immune status of pigs. J. Anim. Sci 2011, 89, 124–135. [Google Scholar]
- Oğuz, H.; Kurtoglu, V.; Coskun, B. Preventive efficacy of clinoptilolite in broilers during chronic aflatoxin (50 and 100 ppb) exposure. Res. Vet. Sci 2000, 69, 197–201. [Google Scholar]
- Magnoli, A.P.; Monge, M.P.; Miazzo, R.D.; Cavaglieri, L.R.; Magnoli, C.E.; Merkis, C.I.; Cristofolini, A.L.; Dalcero, A.M.; Chiacchiera, S.M. Effect of low levels of aflatoxin B1 on performance, biochemical parameters, and aflatoxin B1 in broiler liver tissues in the presence of monensin and sodium bentonite. Poult. Sci 2011, 90, 48–58. [Google Scholar]
- Ozen, H.; Karaman, M.; Cifremio, Y.; Tuzcu, M.; Ozcan, K.; Erdaf, D. Effectiveness of melatonin on aflatoxicosis in chicks. Res. Vet. Sci 2009, 86, 485–489. [Google Scholar]
- Niki, E. Lipid peroxidation: Physiological levels and dual biological effects. Free Radic. Biol. Med 2009, 47, 469–484. [Google Scholar]
- Eraslan, G.; Akdogan, M.; Yarsan, E.; Sahindokuyucu, F.; Essiz, D.; Altintas, L. The effects of aflatoxins on oxidative stress in broiler chickens. Turk. J. Vet. Anim. Sci 2005, 29, 701–707. [Google Scholar]
- Matur, E.; Ergul, E.; Akyazi, I.; Eraslan, E.; Inal, G.; Bilgic, S.; Demircan, H. Effects of saccharomyces cerevisiae extract on haematological parameters, immune function and the antioxidant defence system in breeder hens fed aflatoxin contaminated diets. Br. Poult. Sci 2011, 52, 541–550. [Google Scholar]
- Sies, H. Physiological society symposium: Impaired endothelial and smooth muscle cell function in oxidative stress. Exp. Physiol 1997, 82, 291–295. [Google Scholar]
- Oswald, I.; Comera, C. Immunotoxicity of mycotoxins. Rev. Med. Vet 1998, 149, 585–590. [Google Scholar]
- Bondy, G.S.; Pestka, J.J. Immunomodulation by fungal toxins. J. Toxicol. Environ. Health 2000, 3, 109–143. [Google Scholar]
- Marin, D.; Taranu, I.; Bunaciu, R.; Pascale, F.; Tudor, D.; Avram, N.; Sarca, M.; Cureu, I.; Criste, R.; Suta, V. Changes in performance, blood parameters, humoral and cellular immune responses in weanling piglets exposed to low doses of aflatoxin. J. Anim. Sci 2002, 80, 1250–1257. [Google Scholar]
- Oguz, H.; Hadimli, H.; Kurtoglu, V.; Erganis, O. Evaluation of humoral immunity of broilers during chronic aflatoxin (50 and 100 ppb) and clinoptilolite exposure. Rev. Med. Vet 2003, 154, 483–486. [Google Scholar]
- Choi, K.-C.; Lee, B.-S.; Chung, W.-T.; Choi, M.-S.; Lee, J.-C. Protective effects of apigenin and quercetin on aflatoxin B1-induced immunotoxicity in mice. Food Sci. Biotechnol 2010, 19, 987–992. [Google Scholar]
- Van Heugten, E.; Spears, J.; Coffey, M.; Kegley, E.; Qureshi, M. The effect of methionine and aflatoxin on immune function in weanling pigs. J. Anim. Sci 1994, 72, 658–664. [Google Scholar]
- Panangala, V.; Giambrone, J.; Diener, U.; Davis, N.; Hoerr, F.; Mitra, A.; Schultz, R.; Wilt, G. Effects of aflatoxin on the growth performance and immune responses of weanling swine. Am. J. Vet. Res 1986, 47, 2062–2067. [Google Scholar]
- Dwivedi, N.; Flora, G.; Kushwaha, P.; Flora, S.J. Alpha-lipoic acid protects oxidative stress, changes in cholinergic system and tissue histopathology during co-exposure to arsenic-dichlorvos in rats. Environ. Toxicol. Pharmacol 2013, 37, 7–23. [Google Scholar]
- Meissonnier, G.; Marin, D.; Galtier, P.; Bertin, G.; Taranu, I.; Oswald, I.; Mengheri, E. Modulation of the immune response by a group of fungal food contaminant, the aflatoxins. In Nutrition and Immunity; Mengheri, E., Roselli, M., Britti, M.S., Finamore, A., Eds.; Research Signpost: Kerala, India, 2006; pp. 147–166. [Google Scholar]
- Jakab, G.J.; Hmieleski, R.R.; Zarba, A.; Hemenway, D.R.; Groopman, J.D. Respiratory aflatoxicosis: Suppression of pulmonary and systemic host defenses in rats and mice. Toxicol. Appl. Pharmacol 1994, 125, 198–205. [Google Scholar]
- Dugyala, R.R.; Sharma, R.P. The effect of aflatoxin B1 on cytokine mrna and corresponding protein levels in peritoneal macrophages and splenic lymphocytes. Int. J. Immunopharmacol 1996, 18, 599–608. [Google Scholar]
- Yarru, L.P.; Settivari, R.S.; Gowda, N.K.; Antoniou, E.; Ledoux, D.R.; Rottinghaus, G.E. Effects of turmeric (Curcuma longa) on the expression of hepatic genes associated with biotransformation, antioxidant, and immune systems in broiler chicks fed aflatoxin. Poult. Sci 2009, 88, 2620–2627. [Google Scholar]
- Hinton, D.M.; Myers, M.J.; Raybourne, R.A.; Francke-Carroll, S.; Sotomayor, R.E.; Shaddock, J.; Warbritton, A.; Chou, M.W. Immunotoxicity of aflatoxin B1 in rats: Effects on lymphocytes and the inflammatory response in a chronic intermittent dosing study. Toxicol. Sci 2003, 73, 362–377. [Google Scholar]
- Moon, E.-Y.; Rhee, D.-K.; Pyo, S. In vitro suppressive effect of aflatoxin B1 on murine peritoneal macrophage functions. Toxicology 1999, 133, 171–179. [Google Scholar]
- OLÁH, I.; VERVELDE, L. Structure of the avian lymphoid system. In Avian Immunology; Davison, F., Kaspers, B., Schat, K.A., Eds.; Academic Press: London, UK, 2008; pp. 13–50. [Google Scholar]
- Ellinger-Ziegelbauer, H.; Stuart, B.; Wahle, B.; Bomann, W.; Ahr, H.-J. Characteristic expression profiles induced by genotoxic carcinogens in rat liver. Toxicol. Sci 2004, 77, 19–34. [Google Scholar]
- Abdel-Zaher, A.O.; Abdel-Hady, R.H.; Mahmoud, M.M.; Farrag, M.M.Y. The potential protective role of α-lipoic acid against acetaminophen-induced hepatic and renal damage. Toxicology 2008, 243, 261–270. [Google Scholar]
- Zalejska-Fiolka, J.; Wielkoszyński, T.; Kasperczyk, S.; Kasperczyk, A.; Birkner, E. Effects of oxidized cooking oil and α-lipoic acid on blood antioxidants: Enzyme activities and lipid peroxidation in rats fed a high-fat diet. Biol. Trace Elem. Res 2012, 145, 217–221. [Google Scholar]
- Chen, P.; Ma, Q.-G.; Ji, C.; Zhang, J.-Y.; Zhao, L.-H.; Zhang, Y.; Jie, Y.-Z. Dietary lipoic acid influences antioxidant capability and oxidative status of broilers. Int. J. Mol. Sci 2011, 12, 8476–8488. [Google Scholar]
- Şehirli, Ö.; Tatlıdede, E.; Yüksel, M.; Erzik, C.; Çetinel, S.; Yeğen, B.Ç.; Şener, G. Antioxidant effect of α-lipoic acid against ethanol-induced gastric mucosal erosion in rats. Pharmacology 2007, 81, 173–180. [Google Scholar]
- Maczurek, A.; Hager, K.; Kenklies, M.; Sharman, M.; Martins, R.; Engel, J.; Carlson, D.A.; Munch, G. Lipoic acid as an anti-inflammatory and neuroprotective treatment for Alzheimer’s disease. Adv. Drug Deliv. Rev 2008, 60, 1463–1470. [Google Scholar]
- Zhang, Y.; Han, P.; Wu, N.; He, B.; Lu, Y.; Li, S.; Liu, Y.; Zhao, S.; Liu, L.; Li, Y. Amelioration of lipid abnormalities by α-lipoic acid through antioxidative and anti-inflammatory effects. Obesity 2011, 19, 1647–1653. [Google Scholar]
- Kang, K.P.; Kim, D.H.; Jung, Y.J.; Lee, A.S.; Lee, S.; Lee, S.Y.; Jang, K.Y.; Sung, M.J.; Park, S.K.; Kim, W. α-Lipoic acid attenuates cisplatin-induced acute kidney injury in mice by suppressing renal inflammation. Nephrol. Dial. Transplant 2009, 24, 3012–3020. [Google Scholar]
- Zhang, W.-J.; Wei, H.; Hagen, T.; Frei, B. α-Lipoic acid attenuates LPS-induced inflammatory responses by activating the phosphoinositide 3-kinase/Akt signaling pathway. Proc. Natl. Acad. Sci. USA 2007, 104, 4077–4082. [Google Scholar]
- Chou, T.-C. The protective effect of α-lipoic acid in lipopolysaccharide-induced acute lung injury is mediated by heme oxygenase-1. Evid. Based Complement. Altern. Med 2013, 2013. [Google Scholar] [CrossRef]
- Trucksess, M.W.; Stack, M.E.; Nesheim, S.; Albert, R.H.; Romer, T.R. Multifunctional column coupled with liquid chromatography for determination of aflatoxins B1, B2, G1, and G2 in corn, almonds, brazil nuts, peanuts, and pistachio nuts: Collaborative study. J. AOAC Int 1994, 77, 1512–1521. [Google Scholar]
- National Research Council, Nutrient Requirements of Poultry, 9th rev. ed.; National Academies Press: Washington, DC, USA, 1994.
- Cai, Y.; Song, Z.; Zhang, X.; Wang, X.; Jiao, H.; Lin, H. Increased de novo lipogenesis in liver contributes to the augmented fat deposition in dexamethasone exposed broiler chickens (Gallus gallus domesticus). Comp. Biochem. Physiol 2009, 150, 164–169. [Google Scholar]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem 1979, 95, 351–358. [Google Scholar]
- Winterbourn, C.C.; Hawkins, R.; Brian, M.; Carrell, R. The estimation of red cell superoxide dismutase activity. J. Lab. Clin. Med 1975, 85, 337–341. [Google Scholar]
- Lawrence, R.A.; Burk, R.F. Glutathione peroxidase activity in selenium-deficient rat liver. Biochem. Biophys. Res. Commun 1976, 71, 952–958. [Google Scholar]
- Hong, Y.H.; Lillehoj, H.S.; Hyen Lee, S.; Woon Park, D.; Lillehoj, E.P. Molecular cloning and characterization of chicken lipopolysaccharide-induced TNF-α factor (LITAF). Dev. Comp. Immunol 2006, 30, 919–929. [Google Scholar]
- Druyan, S.; Cahaner, A.; Ashwell, C. The expression patterns of hypoxia-inducing factor subunit α-1, heme oxygenase, hypoxia upregulated protein 1, and cardiac troponin T during development of the chicken heart. Poult. Sci 2007, 86, 2384–2389. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt method. Methods 2001, 25, 402–408. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Gene | Genebank number | Primers position | Primers sequences(5′→3′) | Product size |
|---|---|---|---|---|
| β-actin | AW05994 | Forward | tgcgtgacatcaaggagaag | 300 bp |
| Reverse | tgccagggtacattgtggta | |||
| TNFα # | AY765397.1 | Forward | tgtgtatgtgcagcaacccgtagt | 229 bp |
| Reverse | ggcattgcaatttggacagaagt | |||
| IFNγ | NM_205149.1 | Forward | tgagccagattgtttcgatg | 246 bp |
| Reverse | tccttttgaaactcggagga | |||
| IL6 | NM_204628.1 | Forward | agatgtgcaagaagttcacc | 286 bp |
| Reverse | accacttcatcgggatttat | |||
| NF-κB p65 | D13719.1 | Forward | ttgctgctggagttgatgtc | 167 bp |
| Reverse | tgctatgtgaagaggcgttg | |||
| HO-1 * | NM_205344.1 | Forward | ggtcccgaatgaatgcccttg | 137 bp |
| Reverse | accgttctcctggctcttgg |
© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Li, Y.; Ma, Q.-G.; Zhao, L.-H.; Wei, H.; Duan, G.-X.; Zhang, J.-Y.; Ji, C. Effects of Lipoic Acid on Immune Function, the Antioxidant Defense System, and Inflammation-Related Genes Expression of Broiler Chickens Fed Aflatoxin Contaminated Diets. Int. J. Mol. Sci. 2014, 15, 5649-5662. https://doi.org/10.3390/ijms15045649
Li Y, Ma Q-G, Zhao L-H, Wei H, Duan G-X, Zhang J-Y, Ji C. Effects of Lipoic Acid on Immune Function, the Antioxidant Defense System, and Inflammation-Related Genes Expression of Broiler Chickens Fed Aflatoxin Contaminated Diets. International Journal of Molecular Sciences. 2014; 15(4):5649-5662. https://doi.org/10.3390/ijms15045649
Chicago/Turabian StyleLi, Yan, Qiu-Gang Ma, Li-Hong Zhao, Hua Wei, Guo-Xiang Duan, Jian-Yun Zhang, and Cheng Ji. 2014. "Effects of Lipoic Acid on Immune Function, the Antioxidant Defense System, and Inflammation-Related Genes Expression of Broiler Chickens Fed Aflatoxin Contaminated Diets" International Journal of Molecular Sciences 15, no. 4: 5649-5662. https://doi.org/10.3390/ijms15045649
APA StyleLi, Y., Ma, Q.-G., Zhao, L.-H., Wei, H., Duan, G.-X., Zhang, J.-Y., & Ji, C. (2014). Effects of Lipoic Acid on Immune Function, the Antioxidant Defense System, and Inflammation-Related Genes Expression of Broiler Chickens Fed Aflatoxin Contaminated Diets. International Journal of Molecular Sciences, 15(4), 5649-5662. https://doi.org/10.3390/ijms15045649