Baicalin Inhibits Hypoxia-Induced Pulmonary Artery Smooth Muscle Cell Proliferation via the AKT/HIF-1α/p27-Associated Pathway

Abstract
:1. Introduction
2. Results and Discussion
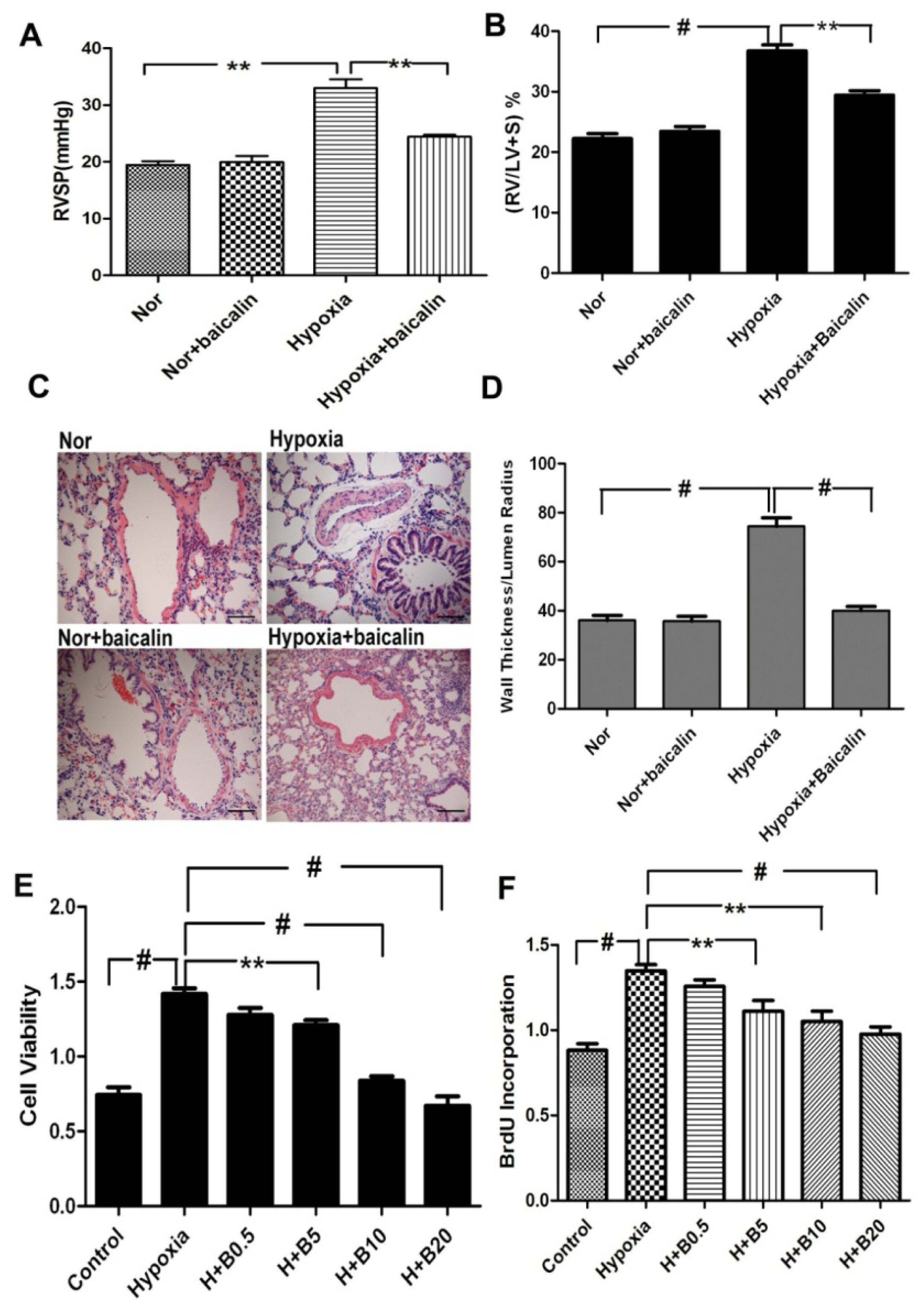
2.1. Baicalin Attenuates Chronic Hypoxia-Induced Pulmonary Hypertension and Pulmonary Vascular Remodeling
2.2. Baicalin Inhibited Hypoxia-Induced Pulmonary Artery Smooth Muscle Cell (PASMCs) Proliferation
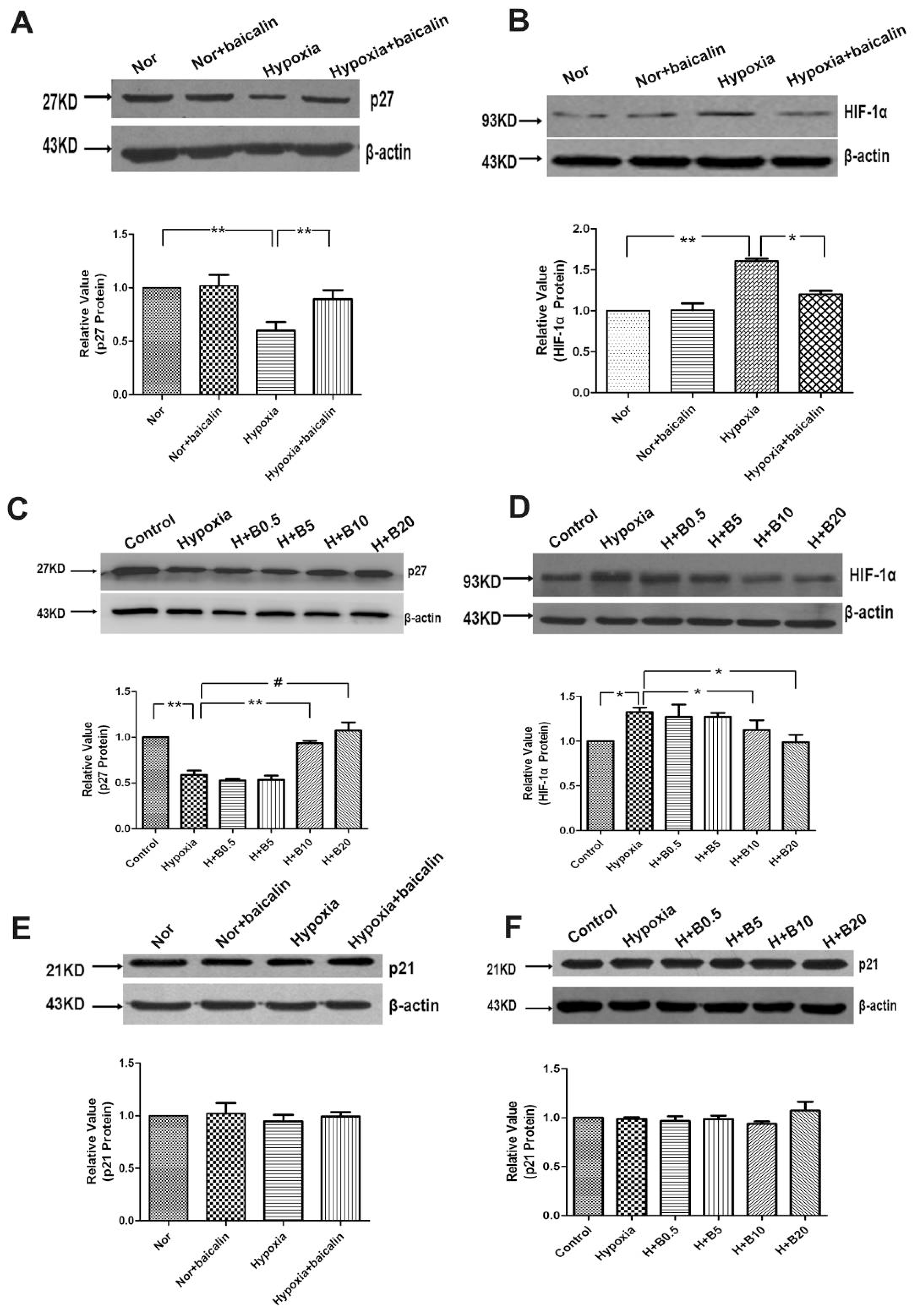
2.3. Baicalin Effected the Protein Expression of p27 and Hypoxia-Inducible Factor-α (HIF-α) in Rat Lung Tissue and Cultured PASMCs
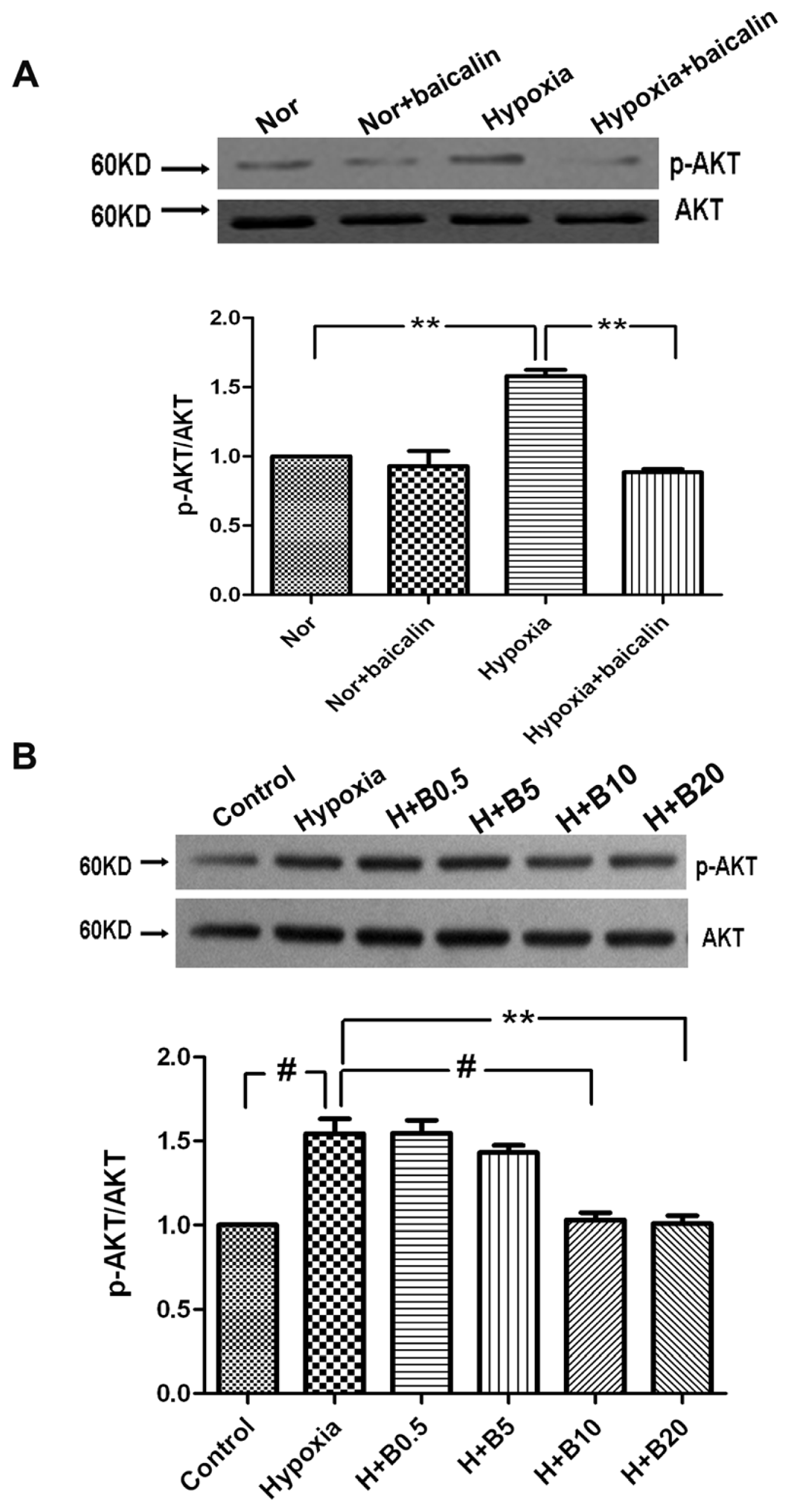
2.4. Protein Expression of p-AKT in Rat Lung Tissue and Cultured PASMCs
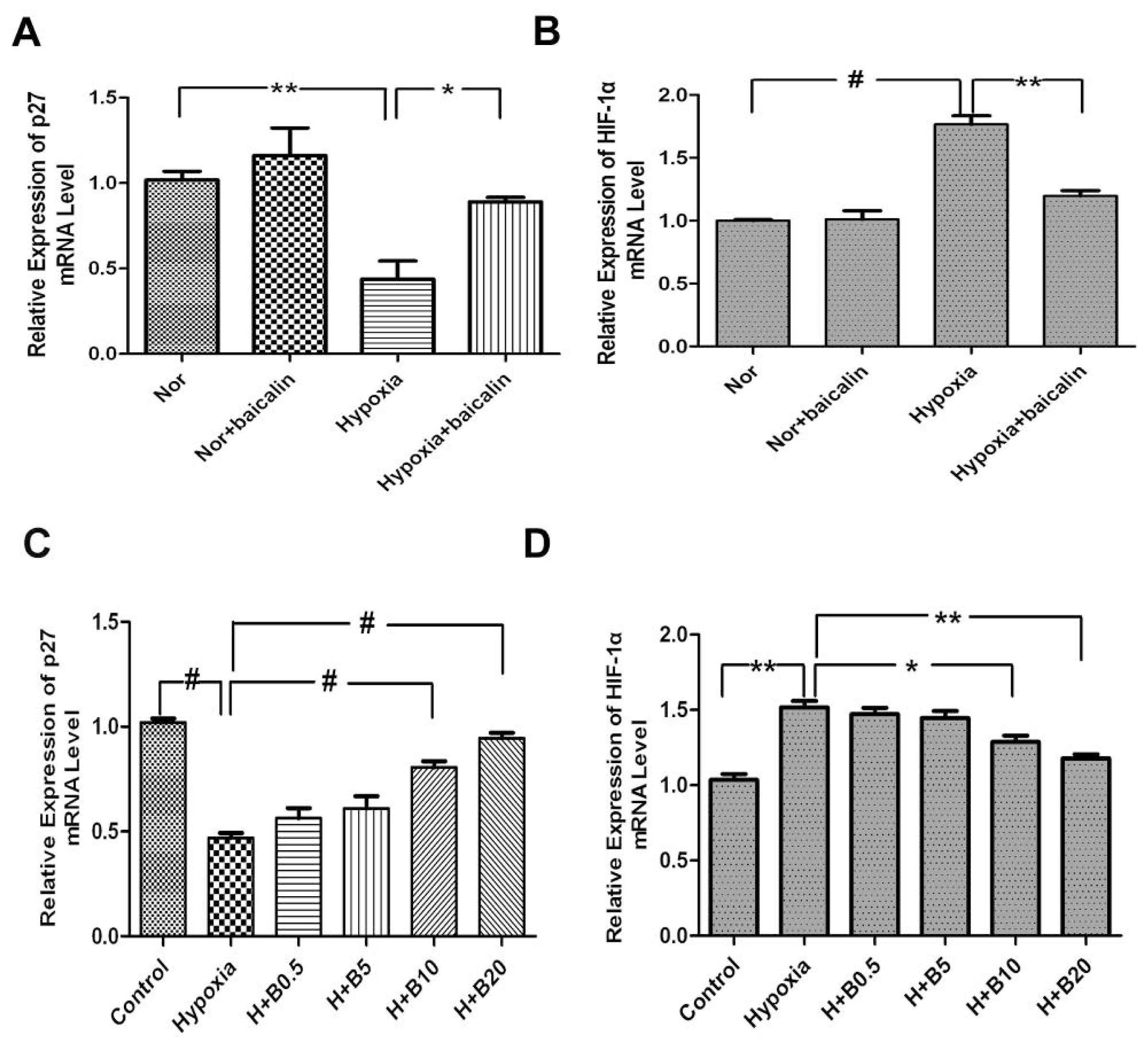
2.5. Changes in mRNA Levels of p27 and HIF-1α in Rat Lung Tissue and Cultured PASMCs
2.6. Discussion
3. Experimental Section
3.1. Reagents and Antibodies
3.2. Animals and Lung Tissue Preparation
3.3. Hemodynamic Experiments
3.4. Morphological Investigation
3.5. Cell Culture
3.6. 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium Bromide (MTT) Assay
3.7. DNA Bromodeoxyuridine (BrdU) Incorporation Assay
3.8. Real-Time Reverse Transcription-Quantitative PCR (RT-qPCR)
3.9. Western Blotting Analysis
3.10. Statistical Analysis
4. Conclusions
Acknowledgments
Conflicts of Interest
- Author ContributionsJ.W. was involved in the conception and hypothesis delineation; L.Z. and Z.P. designed the experiments, conducted the western blotting and quantitative real-time PCR experiments, and wrote the article; J.W. and Z.Z. performed hemodynamic experiments; D.H. designed and conducted the animal studies.
References
- Stenmark, K.R.; Fagan, K.A.; Frid, M.G. Hypoxia-induced pulmonary vascular remodeling: cellular and molecular mechanisms. Circ. Res 2006, 99, 675–691. [Google Scholar]
- Rubin, L.J. Primary pulmonary hypertension. N. Engl. J. Med 1997, 336, 111–117. [Google Scholar]
- Pak, O.; Aldashev, A.; Welsh, D.; Peacock, A. The effects of hypoxia on the cells of the pulmonary vasculature. Eur. Respir. J 2007, 30, 364–372. [Google Scholar]
- Yu, L.; Quinn, D.A.; Garg, H.G.; Hales, C.A. Gene expression of cyclin-dependent kinase inhibitors and effect of heparin on their expression in mice with hypoxia-induced pulmonary hypertension. Biochem. Biophys. Res. Commun 2006, 345, 1565–1572. [Google Scholar]
- Toyoshima, H.; Hunter, T. p27, a novel inhibitor of G1 cyclin-Cdk protein kinase activity, is related to p21. Cell 1994, 78, 67–74. [Google Scholar]
- Sherr, C.J.; Roberts, J.M. Inhibitors of mammalian G1 cyclin-dependent kinases. Genes. Dev 1995, 9, 1149–1163. [Google Scholar]
- Reed, S.I.; Bailly, E.; Dulic, V.; Hengst, L.; Resnitzky, D.; Slingerland, J. G1 control in mammalian cells. J. Cell Sci. Suppl 1994, 18, 69–73. [Google Scholar]
- Xu, D.Q.; Luo, Y.; Liu, Y.; Wang, J.; Zhang, B.; Xu, M.; Wang, Y.X.; Dong, H.Y.; Dong, M.Q.; Zhao, P.T.; Niu, W.; Liu, M.L.; Gao, Y.Q.; Li, Z.C. β-Estradiol attenuates hypoxic pulmonary hypertension by stabilizing the expression of p27 kip1 in rats. Respir. Res 2010, 11, 182. [Google Scholar]
- Krymskaya, V.P.; Snow, J.; Cesarone, G.; Khavin, I.; Goncharov, D.A.; Lim, P.N.; Veasey, S.C.; Ihida-Stansbury, K.; Jones, P.L.; Goncharova, E.A. mTOR is required for pulmonary arterial vascular smooth muscle cell proliferation under chronic hypoxia. FASEB J 2011, 25, 1922–1933. [Google Scholar]
- Gao, Z.; Huang, K.; Xu, H. Protective effects of flavonoids in the roots of Scutellaria baicalensis Georgi against hydrogen peroxide-induced oxidative stress in HS-SY5Y cells. Pharmacol. Res 2001, 43, 173–178. [Google Scholar]
- Ikezoe, T.; Chen, S.S.; Heber, D.; Taguchi, H.; Koeffler, H.P. Baicalin is a major component of PC-SPES which inhibits the proliferation of human cancer cells via apoptosis and cell cycle arrest. Prostate 2001, 49, 285–292. [Google Scholar]
- Shen, Y.C.; Chiou, W.F.; Chou, Y.C.; Chen, C.F. Mechanisms in mediating the anti-inflammatory effects of baicalin and baicalein in human leukocytes. Eur. J. Pharmacol 2003, 465, 171–181. [Google Scholar]
- Dong, L.H.; Wen, J.K.; Miao, S.B.; Jia, Z.; Hu, H.J.; Sun, R.H.; Wu, Y.; Han, M. Baicalin inhibits PDGF-BB-stimulated vascular smooth muscle cell proliferation through suppressing PDGFRβ-ERK signaling and increase in p27 accumulation and prevents injury-induced neointimal hyperplasia. Cell Res 2010, 20, 1252–1262. [Google Scholar]
- Lin, Y.L.; Dai, Z.K.; Lin, R.J.; Chu, K.S.; Chen, I.J.; Wu, J.R.; Wu, B.N. Baicalin, a flavonoid from Scutellaria baicalensis Georgi, activates large-conductance Ca2+-activated K+ channels via cyclic nucleotide-dependent protein kinases in mesenteric artery. Phytomed. Int. J. Phytother. Phytopharmacol 2010, 17, 760–770. [Google Scholar]
- Lee, H.Z.; Leung, H.W.; Lai, M.Y.; Wu, C.H. Baicalein induced cell cycle arrest and apoptosis in human lung squamous carcinoma CH27 cells. Anticancer Res 2005, 25, 959–964. [Google Scholar]
- Ma, Z.; Otsuyama, K.; Liu, S.; Abroun, S.; Ishikawa, H.; Tsuyama, N.; Obata, M.; Li, F.J.; Zheng, X.; Maki, Y.; Miyamoto, K.; Kawano, M.M. Baicalein, a component of Scutellaria radix from Huang-Lian-Jie-Du-Tang (HLJDT), leads to suppression of proliferation and induction of apoptosis in human myeloma cells. Blood 2005, 105, 3312–3318. [Google Scholar]
- Woo, A.Y.; Cheng, C.H.; Waye, M.M. Baicalein protects rat cardiomyocytes from hypoxia/reoxygenation damage via a prooxidant mechanism. Cardiovasc. Res 2005, 65, 244–253. [Google Scholar]
- Chao, J.I.; Su, W.C.; Liu, H.F. Baicalein induces cancer cell death and proliferation retardation by the inhibition of CDC2 kinase and survivin associated with opposite role of p38 mitogen-activated protein kinase and AKT. Mol. Cancer Ther 2007, 6, 3039–3048. [Google Scholar]
- Cogolludo, A.; Moreno, L.; Villamor, E. Mechanisms controlling vascular tone in pulmonary arterial hypertension: implications for vasodilator therapy. Pharmacology 2007, 79, 65–75. [Google Scholar]
- Chen, Z.Y.; Su, Y.L.; Lau, C.W.; Law, W.I.; Huang, Y. Endothelium-dependent contraction and direct relaxation induced by baicalein in rat mesenteric artery. Eur. J. Pharmacol 1999, 374, 41–47. [Google Scholar]
- Akyurek, L.M.; Boehm, M.; Olive, M.; Zhou, A.X.; San, H.; Nabel, E.G. Deficiency of cyclin-dependent kinase inhibitors p21 Cip1 and p27 Kip1 accelerates atherogenesis in apolipoprotein E-deficient mice. Biochem. Biophys. Res. Commun 2010, 396, 359–363. [Google Scholar]
- Tanner, F.C.; Boehm, M.; Akyurek, L.M.; San, H.; Yang, Z.Y.; Tashiro, J.; Nabel, G.J.; Nabel, E.G. Differential effects of the cyclin-dependent kinase inhibitors p27 (Kip1), p21 (Cip1), and p16 (Ink4) on vascular smooth muscle cell proliferation. Circulation 2000, 101, 2022–2025. [Google Scholar]
- Fouty, B.W.; Grimison, B.; Fagan, K.A.; le Cras, T.D.; Harral, J.W.; Hoedt-Miller, M.; Sclafani, R.A.; Rodman, D.M. p27 (Kip1) is important in modulating pulmonary artery smooth muscle cell proliferation. Am. J. Respir. Cell. Mol. Biol 2001, 25, 652–658. [Google Scholar]
- Mizuno, S.; Kadowaki, M.; Demura, Y.; Ameshima, S.; Miyamori, I.; Ishizaki, T. p42/44 mitogen-activated protein kinase regulated by p53 and nitric oxide in human pulmonary arterial smooth muscle cells. Am. J. Respir. Cell. Mol. Biol 2004, 31, 184–192. [Google Scholar]
- Mizuno, S.; Bogaard, H.J.; Kraskauskas, D.; Alhussaini, A.; Gomez-Arroyo, J.; Voelkel, N.F.; Ishizaki, T. p53 Gene deficiency promotes hypoxia-induced pulmonary hypertension and vascular remodeling in mice. Am. J. Physiol. Lung. Cell. Mol. Physiol 2011, 300, L753–L761. [Google Scholar]
- Yu, L.; Quinn, D.A.; Garg, H.G.; Hales, C.A. Cyclin-dependent kinase inhibitor p27 Kip1, but not p21 WAF1/Cip1, is required for inhibition of hypoxia-induced pulmonary hypertension and remodeling by heparin in mice. Circ. Res 2005, 97, 937–945. [Google Scholar]
- Yu, A.Y.; Shimoda, L.A.; Iyer, N.V.; Huso, D.L.; Sun, X.; McWilliams, R.; Beaty, T.; Sham, J.S.; Wiener, C.M.; Sylvester, J.T.; Semenza, G.L. Impaired physiological responses to chronic hypoxia in mice partially deficient for hypoxia-inducible factor 1α. J. Clin. Invest 1999, 103, 691–696. [Google Scholar]
- Shimoda, L.A.; Manalo, D.J.; Sham, J.S.; Semenza, G.L.; Sylvester, J.T. Partial HIF-1α deficiency impairs pulmonary arterial myocyte electrophysiological responses to hypoxia. Am. J. Physiol. Lung. Cell. Mol. Physiol 2001, 281, L202–L208. [Google Scholar]
- Ke, Q.; Costa, M. Hypoxia-inducible factor-1 (HIF-1). Mol. Pharmacol 2006, 70, 1469–1480. [Google Scholar]
- Bilton, R.L.; Booker, G.W. The subtle side to hypoxia inducible factor (HIFα) regulation. Eur. J. Biochem 2003, 270, 791–798. [Google Scholar]
- Zhu, Y.; Zhao, T.; Itasaka, S.; Zeng, L.; Yeom, C.J.; Hirota, K.; Suzuki, K.; Morinibu, A.; Shinomiya, K.; Ou, G.; et al. Involvement of decreased hypoxia-inducible factor 1 activity and resultant G1-S cell cycle transition in radioresistance of perinecrotic tumor cells. Oncogene 2013, 32, 2058–2068. [Google Scholar]
- Garat, C.V.; Fankell, D.; Erickson, P.F.; Reusch, J.E.; Bauer, N.N.; McMurtry, I.F.; Klemm, D.J. Platelet-derived growth factor BB induces nuclear export and proteasomal degradation of CREB via phosphatidylinositol 3-kinase/AKT signaling in pulmonary artery smooth muscle cells. Mol. Cell. Biol 2006, 26, 4934–4948. [Google Scholar]
- Gao, D.; Inuzuka, H.; Tseng, A.; Wei, W. AKT finds its new path to regulate cell cycle through modulating Skp2 activity and its destruction by APC/Cdh1. Cell Div 2009, 4, 11. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequences(5′-to-3′) | PCR Product Size | Accession Number |
|---|---|---|---|
| p27 | Forward:5′-CGAACGGGCTCAAGAT-3′ Reverse:5′-CTGTGACGAGGCGATT-3′ | 198 bp | NM: U10440.1 |
| HIFα | Forward:5′-GAATTAAACCCAAAGAC-3′ Reverse:5′-CAAGAAAGCGACATAG-3′ | 143 bp | NM: 024359 |
| β-actin | Forward:5′-GTTGACATCCGTAAAGACC-3′ Reverse:5′-GGAGCCAGGGCAGTAA-3′ | 107 bp | EF: 156276.1 |
© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Zhang, L.; Pu, Z.; Wang, J.; Zhang, Z.; Hu, D.; Wang, J. Baicalin Inhibits Hypoxia-Induced Pulmonary Artery Smooth Muscle Cell Proliferation via the AKT/HIF-1α/p27-Associated Pathway. Int. J. Mol. Sci. 2014, 15, 8153-8168. https://doi.org/10.3390/ijms15058153
Zhang L, Pu Z, Wang J, Zhang Z, Hu D, Wang J. Baicalin Inhibits Hypoxia-Induced Pulmonary Artery Smooth Muscle Cell Proliferation via the AKT/HIF-1α/p27-Associated Pathway. International Journal of Molecular Sciences. 2014; 15(5):8153-8168. https://doi.org/10.3390/ijms15058153
Chicago/Turabian StyleZhang, Lin, Zhichen Pu, Junsong Wang, Zhifeng Zhang, Dongmei Hu, and Junjie Wang. 2014. "Baicalin Inhibits Hypoxia-Induced Pulmonary Artery Smooth Muscle Cell Proliferation via the AKT/HIF-1α/p27-Associated Pathway" International Journal of Molecular Sciences 15, no. 5: 8153-8168. https://doi.org/10.3390/ijms15058153
APA StyleZhang, L., Pu, Z., Wang, J., Zhang, Z., Hu, D., & Wang, J. (2014). Baicalin Inhibits Hypoxia-Induced Pulmonary Artery Smooth Muscle Cell Proliferation via the AKT/HIF-1α/p27-Associated Pathway. International Journal of Molecular Sciences, 15(5), 8153-8168. https://doi.org/10.3390/ijms15058153