Subchronic Toxicity of Copper Oxide Nanoparticles and Its Attenuation with the Help of a Combination of Bioprotectors


Abstract
:1. Introduction


2. Results and Discussion
2.1. Copper Accumulation in Different Organs and Morphological Changes Caused by it

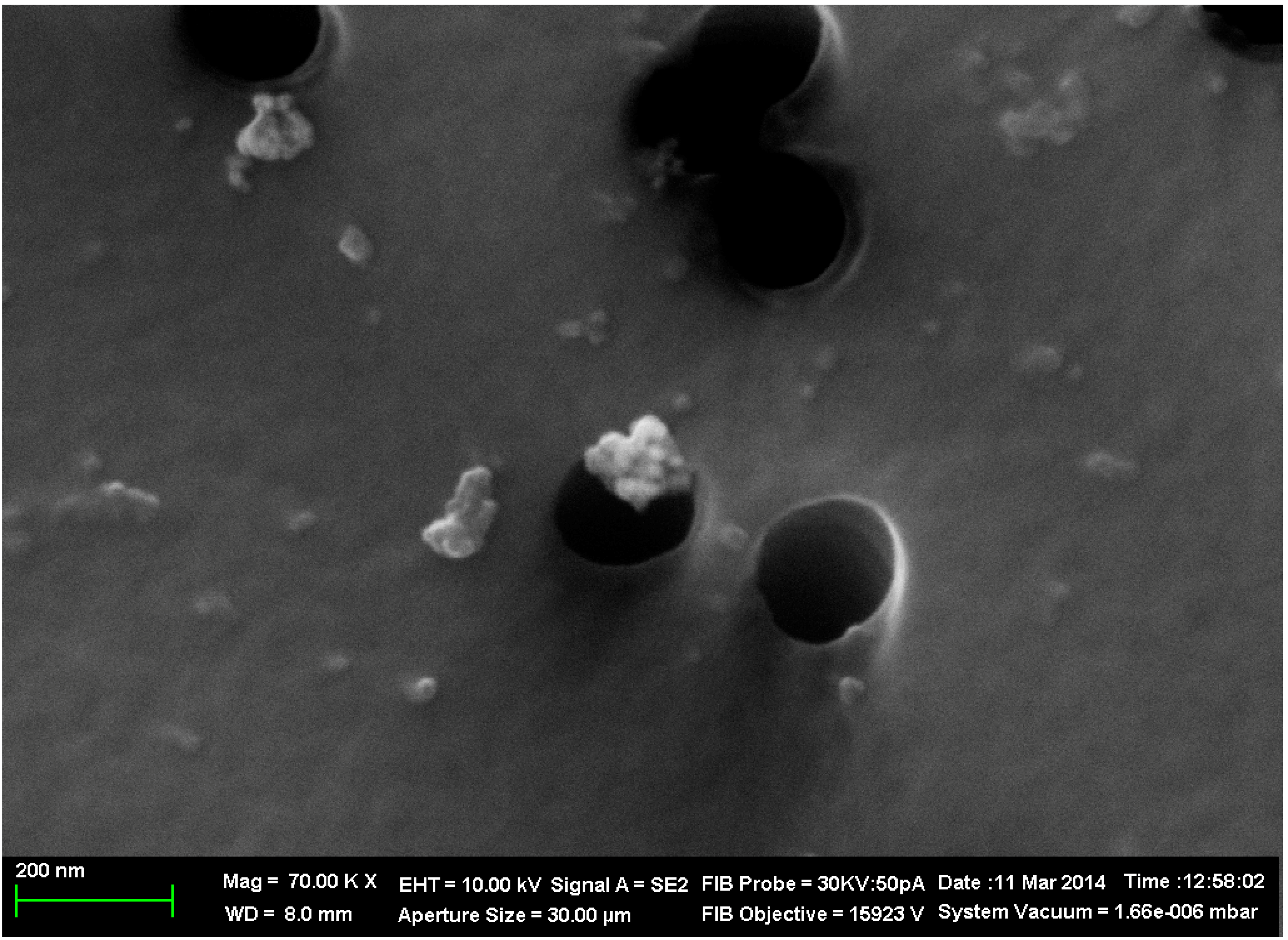{kind=link}
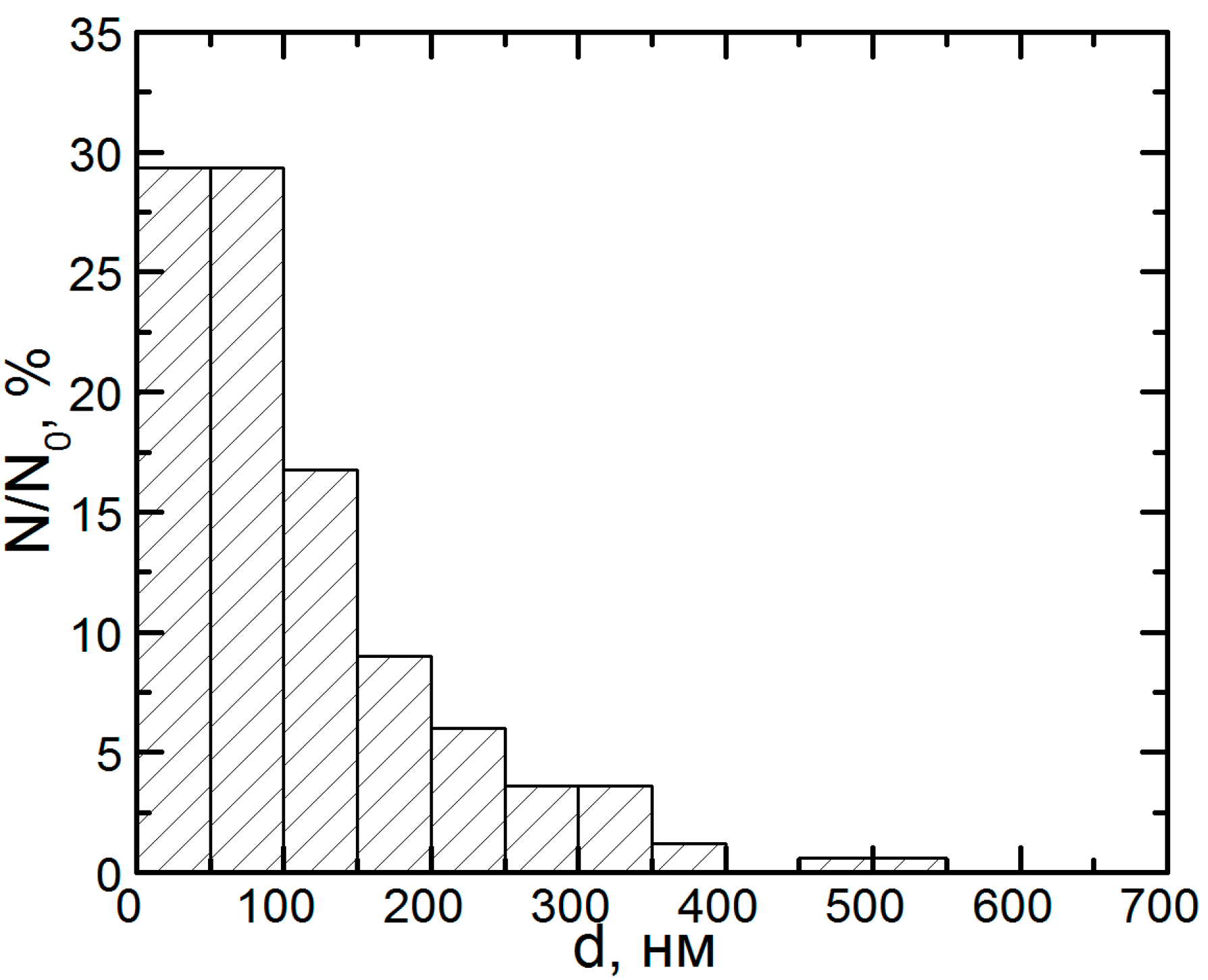{kind=link}
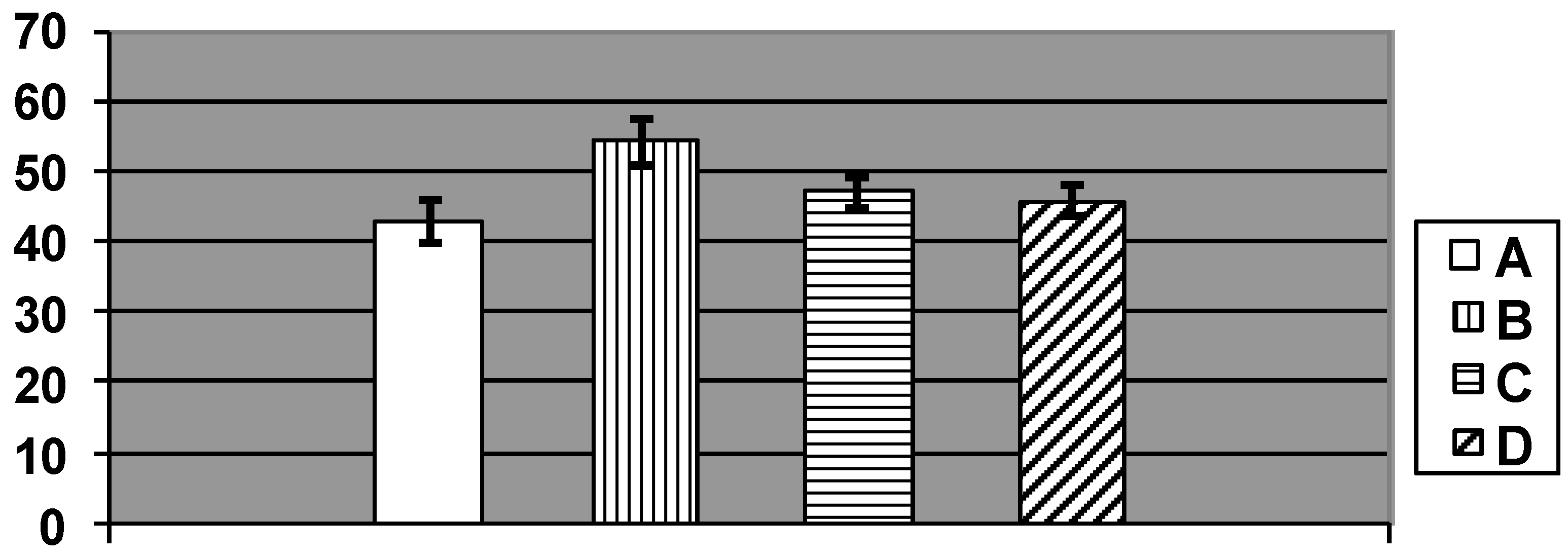{kind=link}
{kind=link}
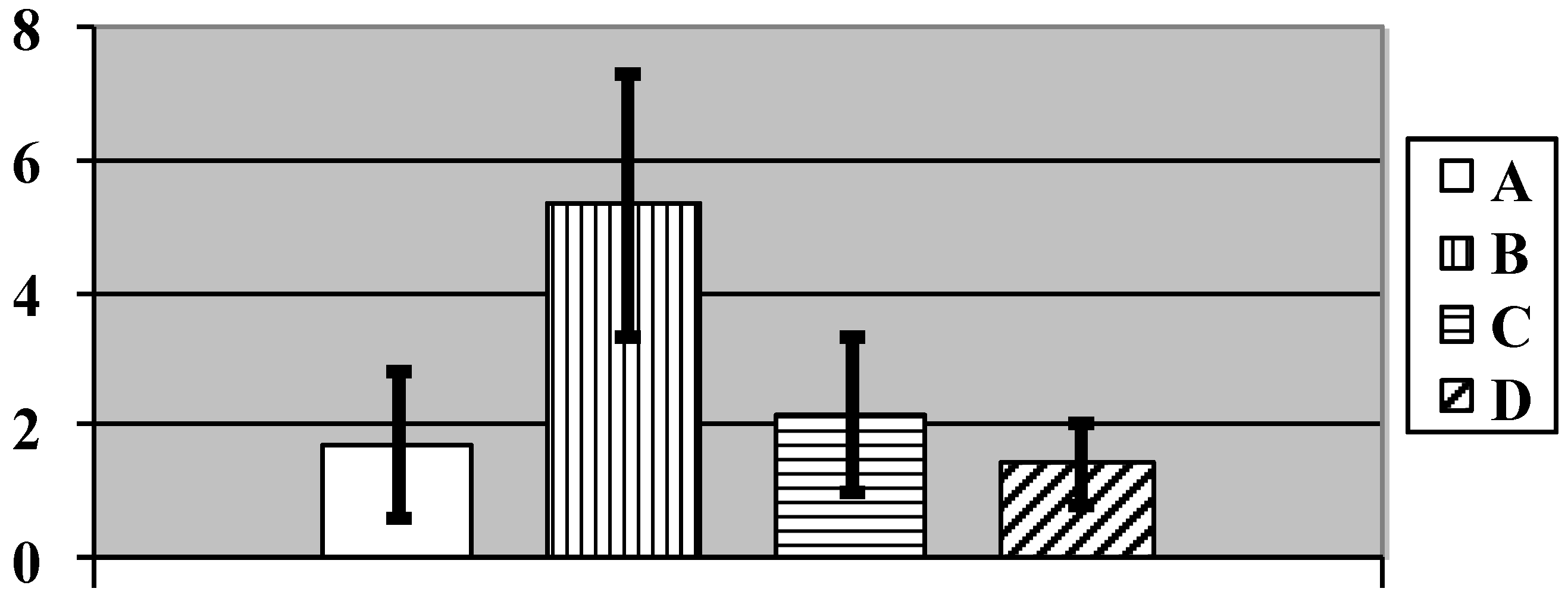{kind=link}
{kind=link}
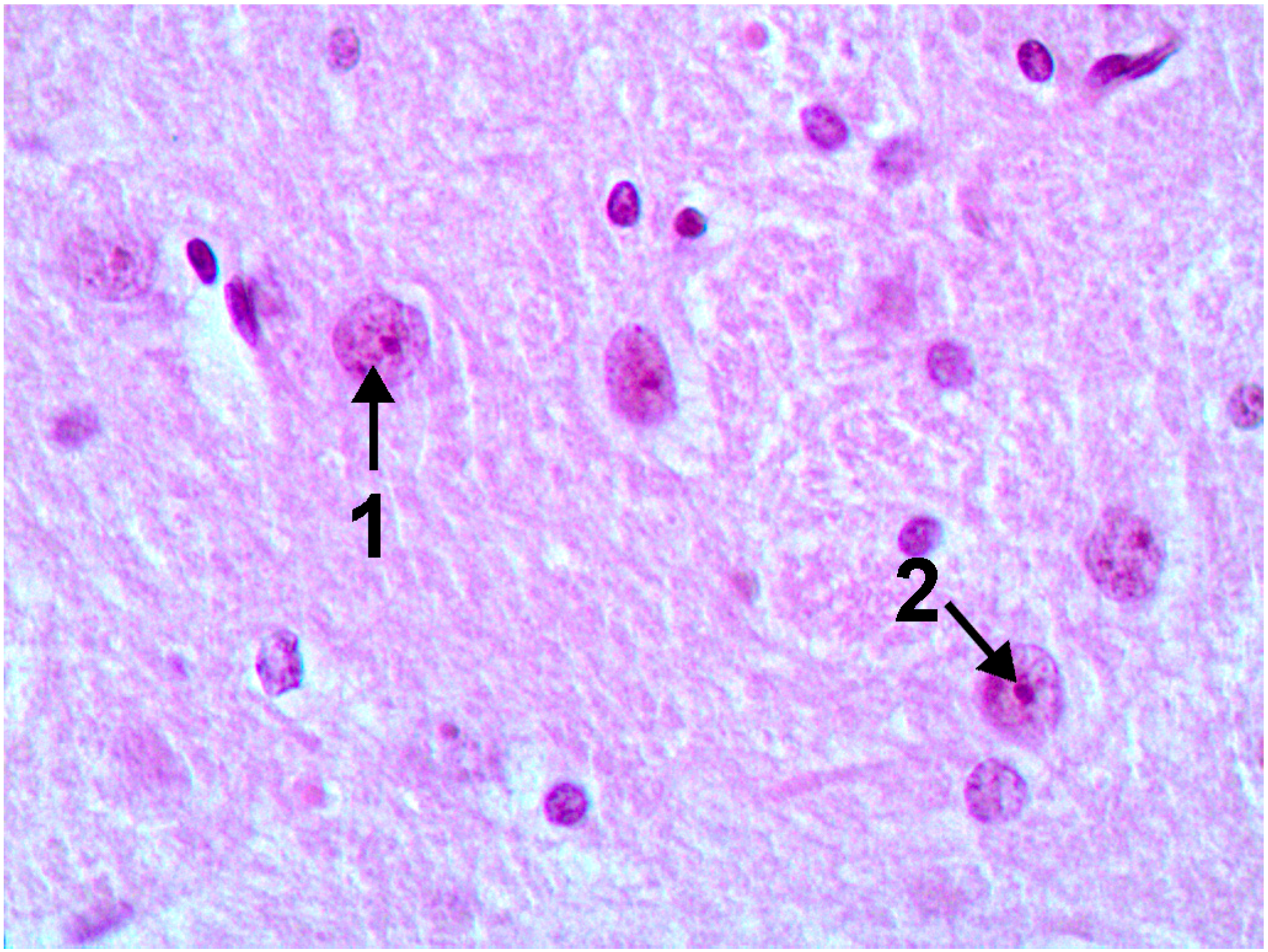{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
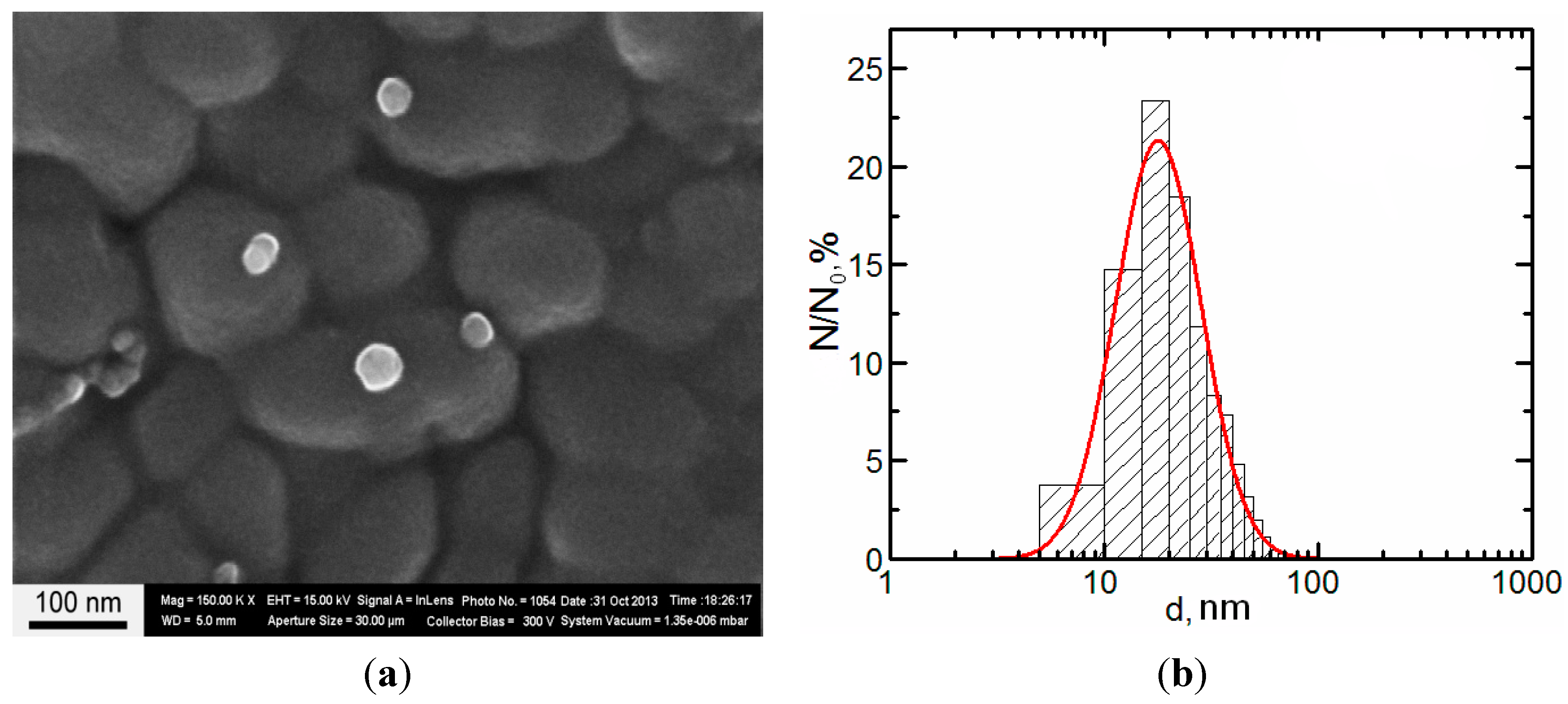{kind=link}
| Group of Rats Given | Kidneys | Liver | Spleen | Brain |
|---|---|---|---|---|
| Water (control) | 42.4 ± 2.9 | 12.2 ± 2.4 | 22.5 ± 2.1 | 18.9 ± 0.7 |
| Nano copper oxide | 62.5 ± 7.1 * | 28.8 ± 6.3 * | 24.2 ± 1.5 | 21.5 ± 1.7 |
| Nano copper oxide + BPC | 59.4 ± 10.0 | 22.1 ± 3.5 * | 18.0 ± 2.5 ° | 18.8 ± 1.4 |
| BPC | 50.4 ± 5.6 | 10.6 ± 0.3 | 25.3 ± 2.2 | 20.8 ± 1.5 |


| Groups of Rats Given | Glomerular Surface Area (mm2) | Urinary Space (mm2) | Brush Border Loss (% Lengthwise) | Epithelial Desquamation (% Lengthwise) |
|---|---|---|---|---|
| Water (control) | 2.47 ± 0.06 | 0.71 ± 0.04 | 5.39 ± 0.42 | 0.33 ± 0.13 |
| Nano copper oxide | 2.43 ± 0.04 | 0.89 ± 0.03 * | 8.36 ± 0.76 * | 1.16 ± 0.38 * |
| Nano copper oxide + BPC | 2.82 ± 0.075 * | 0.97 ± 0.05 *,▪ | 5.98 ± 0.46 ▪ | 0.98 ± 0.35 |
| BPC | 2.41 ± 0.05 | 0.74 ± 0.05 | 6.03 ± 0.57 | 0.73 ± 0.21 |








| Organs | Groups of Rats Given | |||
|---|---|---|---|---|
| Nano Copper Oxide | Nano Copper Oxide + BPC | BPC | Water (Control) | |
| Liver | 0.426 ± 0.0020 * | 0.404 ± 0.002 *,▫,▪ | 0.394 ± 0.0040 | 0.396 ± 0.0020 |
| Kidneys | 0.382 ± 0.0015 | 0.406 ± 0.0027 *,▫,▪ | 0.393 ± 0.0025 * | 0.383 ± 0.0025 |
| Spleen | 0.460 ± 0.0020 * | 0.418 ± 0.0015 *,▫,▪ | 0.377 ± 0.0028 * | 0.369 ± 0.0016 |
| Brain | 0.355 ± 0.0020 | 0.335 ± 0.0021 *,▫ | 0.356 ± 0.0025 | 0.354 ± 0.0028 |
2.2. Functional and Biochemical Signs of Intoxication
- A statistically significant increase in mass coefficients of the liver, kidneys and spleen;
- Statistically significant adverse changes in the red blood cells (a decrease in the haemoglobin content and number of erythrocytes with an increased percentage of reticulocytes—this result is most likely due to the haemolytic effect of copper) and in the porphyrin metabolism (an insignificant increase in renal excretion of coproporphyrin with a statistically significantly reduced excretion of delta-aminolevulinic acid);
- General inhibition of oxidative metabolism (judging by the statistically significant decrease in the SDH activity of blood lymphocytes), and of lipid peroxidation (decreased renal excretion of malonyldialdehyde (MDA)).
- A reduced hemoglobin content against an increased percentage of reticulocytes;
- A lowered protein-producing function of the liver (judging by total protein and albumin contents of the blood serum);
- A lowered level of oxidative metabolism (judging by the succinate dehydrogenase activity of blood lymphocytes) and of lipid peroxidation (judging by the MDA content of the blood serum).
| Indices | Groups of Rat Receiving | ||||
|---|---|---|---|---|---|
| Nano Copper Oxide | Nano Copper Oxide + BPC | BPC | Water (Controls) | ||
| Initial body mass, g | 178.8 ± 2.5 | 175.8 ± 1.7 | 178.8 ± 4.9 | 174.6 ± 2.7 | |
| Body mass after period of injections, g | 225.8 ± 8.0 | 232.5 ± 6.2 | 227.5 ± 6.8 | 217.0 ± 6.4 | |
| Temporal summation of sub-threshold impulses, s | 12.2±1.1 | 12.35 ± 1.6 | 13.4 ± 0.9 | 13.4 ± 0.8 | |
| Number of head-dips into holes during 3 min | 6.1± 1.1 | 5.1 ± 1.0 * | 8.1 ± 1.0 | 9.1 ± 1.70 | |
| Haemoglobin, g/L | 13.4 ± 0.4 * | 14.3 ± 0.3 *,▪,▫ | 16.7 ± 0.5 | 16.6 ± 0.4 | |
| Erythrocytes, 1012 g/L | 1.6 ± 0.1 * | 1.63 ± 0.0 * | 1.69 ± 0.1 | 1.84 ± 0.0 | |
| Reticulocytes, ‰ | 28.9 ± 2.1 * | 16.0 ± 1.6 *,▪,▫ | 11.0 ± 1.9 | 10.45 ± 0.8 | |
| Lymphocytes, % | 56.9 ± 2.9 | 52.3 ± 3.4 | 55.2 ± 3.1 | 56.3 ± 2.2 | |
| Segmented neutrophils, % | 57.7 ± 4.2 | 62.9 ± 5.3 | 60.7 ± 4.3 | 54.7 ± 2.1 | |
| Band neutrophils,% | 3.2 ± 0.7 | 3.9 ± 0.7 | 3.6 ± 0.5 | 3.1 ± 0.8 | |
| Monocytes, % | 19.6 ± 2.2 | 19.6 ± 2.8 | 16.6 ± 2.3 | 18.4 ± 1.4 | |
| Eosinophils, % | 7.0 ± 1.4 | 8.0 ± 1.5 | 7.3 ± 2.2 | 10.3 ± 1.4 | |
| Basophils, % | 0.6 ± 0.2 | 1.1 ± 0.3 | 0.73 ± 0.2 | 0.9 ± 0.3 | |
| Total protein in blood serum, g/L | 70.3 ± 1.2 | 74.8 ± 0.7 ▪ | 73.2 ± 1.4 | 72.4 ± 1.3 | |
| Albumins in blood serum, g/L | 36.9 ± 0.7 | 40.6 ± 0.6 ▪ | 39.1 ± 0.5 | 38.3 ± 1.0 | |
| Globulins in blood serum, g/L | 33.3 ± 1.0 | 34.2 ± 0.7 | 34.1 ±1.2 | 34.0 ± 1.1 | |
| A/G index | 1.1 ± 0.0 | 1.2 ± 0.0 | 1.16 ± 0.0 | 1.14 ± 0.1 | |
| SDH activity, number of formasan granules in 50 lymphocytes | 656.8 ± 12.0 * | 807.5 ± 20.9 *,▪ | 766.1 ± 19.5 | 737.1 ± 10.7 | |
| SH-gtoups, mmol/L | 1.1 ± 0.0 | 1.0 ± 0.1 | 1.1 ± 0.0 | 1.1 ± 0.0 | |
| GSH-gtoups, mmol/L | 0.6 ± 0.1 | 0.5 ± 0.1 | 0.4 ± 0.1 | 0.5 ± 0.0 | |
| ALT activity in blood serum, mmol/h L | 49.3 ± 2.7 | 54.6 ± 3.3 | 62.5 ± 3.9 | 52.8 ± 3.6 | |
| AST activity in blood serum, mmol/h L | 234.8 ± 13.3 | 218.8 ± 14.5 | 235.9 ± 16.5 | 254.7 ± 16.8 | |
| Catalase in blood serum, μmol/L | 1.4 ± 1.1 | 1.2 ± 0.2 | 1.5 ± 0.1 | 1.2 ± 0.2 | |
| MDA in blood serum, nmol/L | 3.7 ± 0.2 * | 4.3 ± 0.2 ▪ | 4.1 ± 0.2 | 4.0 ± 0.1 | |
| Ceruloplasmin in blood serum, mg % | 80.0 ± 5.5 | 100.5 ± 3.1 ▪ | 109.9 ± 8.0 | 90.9 ± 45.0 | |
| Bilirubin in blood serum, μmol/L | 2.4 ± 0.3 | 2.8 ± 0.4 | 3.2 ± 0.6 | 2.8 ± 0.6 | |
| Alkaline phosphatase in blood serum, nmol/s L | 117.5 ± 11.1 | 185.7 ± 18.5 *,▪ | 142.8 ± 11.4 * | 97.8 ± 11.7 | |
| γ-Glutamintransferase, units/L | 3.5± 0.8 | 4.1 ± 1.1 | 5.6± 1.4 | 3.7 ± 1.0 | |
| Creatinine in blood serum, μmol/L | 34.6 ± 1.34 | 35.2 ± 1.43 | 35.8 ± 1.3 | 36.8 ± 1.0 | |
| Daily volume of urine, mL | 39.9 ± 6.2 | 42.6 ± 4.8 | 43.9 ± 6.3 | 38.3 ± 7.0 | |
| Creatinine in urine, mol/L | 1.1 ± 0.3 | 0.8 ± 0.1 | 0.8 ± 0.1 | 0.8 ± 0.1 | |
| Coproporphyrin in urine, nM/L | 89.2 ± 9.5 | 76.4 ± 13.4 | 85.2 ± 17.2 | 69.0 ± 12.1 | |
| δ—ALK in urine, μmol/L | 5.9 ± 1.1 * | 6.3 ± 1.3 | 8.3 ± 1.3 | 10.4 ± 1.8 | |
| Liver mass, g per 100 g body mass | 4.0 ± 0.1 * | 4.0 ± 0.2 | 3.5 ± 0.1 | 3.6 ± 0.1 | |
| Kidney mass, g per 100 g body mass | 0.6 ± 0.0 * | 0.7 ± 0.0 * | 0.7± 0.0 * | 0.4 ± 0.0 | |
| Spleen mass, g per 100 g body mass | 0.5 ± 0.0 * | 0.5 ± 0.1 | 0.4 ± 0.0 | 0.4 ± 0.0 | |
| Brain mass, g per 100 g body mass | 0.8 ± 0.0 | 0.7± 0.0 | 0.8 ± 0.0 | 0.8 ± 0.0 | |
3. Experimental Section

- Weighing;
- Estimation of the CNS ability to the temporal summation of sub-threshold impulses—A variant of withdrawal reflex and its facilitation by repeated electrical stimulations in intact, conscious rat [44];
- Collection of daily urine for analysis of its density, urine output, coproporhyrin, δ-aminolevulinic acid (δ-ALA), and creatinine contents;
3.1. Testing of Genotoxicity (the Random Amplification of Polymorphic DNA (RAPD) Test, Random Amplification of Polymorphic DNA)
3.2. Choice of Bioprotectors
- Glutamate as an effective cell membrane stabilizer through the intensification of ATP synthesis under exposure to the damaging action of various cytotoxic particles (e.g., [7,48,49,50,51] and, at the same time, as one of the three precursors of glutathione, a powerful cell protector against free radicals.
- The other two precursors of glutathione: Glycine and cysteine (the latter in a highly active and metabolically well available form of N-acetylcysteine).
- Other components of the organism’s anti-oxidant system (vitamins А, Е, and С, and selenium).
- Trace elements, which are physiological antagonists of copper (zink, iron, molybdenum, manganese).
- Pectin enterosorbent as an agent that prevents the re-absorption of the metal excreted into the intestines with bile.
| Bioprotectors (See the Text for Explanation) | Estimated Dosage and the Mode of Administration |
|---|---|
| Sodium glutamate | 800–900 mg/kg (as a 1.5% drink instead of water) |
| Apple pectin | 1 g/kg (added to the fodder) |
| Glycine | 12 mg per rat (added to the fodder) |
| Acetyl-cysteine | 30 mg per rat (added to the fodder) |
| A commercial fish oil rich in vitamin A and omega 3 rich PUFA | 1 mL per rat (by gavage) |
| Multivitamin-Multimineral Preparations (Added to the Fodder) Supplying | |
| Vitamin B12 | 0.15 mcg per rat |
| Vitamin C | 4.4 mg per rat |
| Vitamin E | 0.84 vg per rat |
| Iron | 0.6 mg per rat |
| Selenium | 5.8 mcg per rat |
| Zink | 1.25 mg per rat |
| Molibdenum | 3.75 mcg per rat |
| Manganese | 16.7 mcg per rat |
4. Conclusions
Author Contributions
Conflicts of Interest
References
- Katsnelson, B.A.; Privalova, L.I.; Kuzmin, S.V.; Degtyareva, T.D.; Sutunkova, M.P.; Yeremenko, O.S.; Minigalieva, I.A.; Kireyeva, E.P.; Khodos, M.Y.; Kozitsina, A.N.; et al. Some peculiarities of pulmonary clearance mechanisms in rats after intratracheal instillation of magnetite (Fe3O4) suspensions with different particle sizes in the nanometer and micrometer ranges: Are we defenseless against nanoparticles? Int. J. Occup. Environ. Health 2010, 16, 508–524. [Google Scholar] [CrossRef]
- Katsnelson, B.A.; Privalova, L.I.; Degtyareva, T.D.; Sutunkova, M.P.; Yeremenko, O.S.; Minigalieva, I.A.; Kireyeva, E.P.; Kozitsina, A.N.; Malakhova, N.A.; Glazyrina, J.A.; et al. Experimental estimates of the toxicity of iron oxide Fe3O4 (magnetite) nanoparticles. Cent. Eur. J. Occup. Environ. Med. 2010, 16, 47–63. [Google Scholar]
- Katsnelson, B.A.; Degtyareva, T.D.; Minigalieva, I.A.; Privalova, L.I.; Kuzmin, S.V.; Yeremenko, O.S.; Kireyeva, E.P.; Sutunkova, M.P.; Valamina, I.E.; Khodos, M.Y.; et al. Sub-chronic systemic toxicity and bio-accumulation of Fe3O4 nano- and microparticles following repeated intraperitoneal administration to rats. Int. J. Toxicol. 2010, 30, 60–67. [Google Scholar]
- Katsnelson, B.A.; Privalova, L.I.; Sutunkova, M.P.; Tulakina, L.G.; Pichugova, S.V.; Beikin, J.B.; Khodos, M.Y. The “in vivo” interaction between iron oxide Fe3O4 nanoparticles and alveolar macrophages. Bull. Exp. Biol. Med. 2012, 152, 627–631. [Google Scholar] [CrossRef]
- Katsnelson, B.A.; Privalova, L.I.; Sutunkova, M.P.; Khodos, M.Y.; Shur, V.Y.; Shishkin, E.I.; Tulakina, L.G.; Pichugova, S.V.; Beikin, J.B. Uptake of some metallic nanoparticles by, and their impact on pulmonary macrophages in vivo as viewed by optical, atomic force, and transmission electron microscopy. J. Nanomed. Nanotechnol. 2012, 3, 1–8. [Google Scholar]
- Katsnelson, B.A.; Privalova, L.I.; Kuzmin, S.V.; Gurvich, V.B.; Sutunkova, M.P.; Kireyeva, E.P.; Minigalieva, I.A. An approach to tentative reference levels setting for nanoparticles in the workroom air based on comparing their toxicity with that of their micrometric counterparts: A case study of iron oxide Fe3O4. ISRN Nanotechnol. 2012, 2012, 12. [Google Scholar]
- Katsnelson, B.A.; Privalova, L.I.; Gurvich, V.B.; Makeyev, O.H.; Shur, V.Y.; Beikin, J.B.; Sutunkova, M.P.; Kireyeva, E.P.; Minigalieva, I.A.; Loginova, N.V; et al. Comparative in vivo assessment of some adverse bio-effects of equidimensional gold and silver nanoparticles and the attenuation of nanosilver’s effects with a complex of innocuous bioprotectors. Int. J. Mol. Sci. 2013, 14, 2449–2483. [Google Scholar] [CrossRef]
- Katsnelson, B.A.; Privalova, L.I.; Gurvich, V.B.; Makeyev, O.H.; Shur, V.Y.; Beikin, J.B.; Sutunkova, M.P.; Kireyeva, E.P.; Minigalieva, I.A.; Loginova, N.V.; et al. Some inferences from toxicological experiments with metal nanoparticles: The pulmonary phagocytosis response to their deposition. subchronic systemic toxicity and genotoxicity; the attenuation of the latter with bioprotectors. In Second Int. Sch.-Conf. Appl. Nanotechno. Nanotoxicol; The Siberian Branch of the RAS Boreskov Institute of Caralysis: Novosibirsk, Russia, 2013; pp. 48–49. [Google Scholar]
- Loginova, N.V. Experimental assessment of efficacy of a bio-prophylactic complex aimed at attenuating toxic effects of copper oxide nanoparticles. Med. Promyshlenaya Ekol. 2014. in press (In Russian) [Google Scholar]
- Karlsson, H.L.; Cronholm, P.; Gustafsson, J.; Möller, L. Copper oxide nanoparticles are highly toxic: A comparison between metal oxide nanoparticles and carbon nanotubes. Chem. Res. Toxicol. 2008, 21, 1726–1732. [Google Scholar]
- Bondarenko, O.; Ivask, A.; Käkinen, A.; Kahru, A. Sub-toxic effects of CuO nanoparticles on bacteria: Kinetics, role of Cu ions and possible mechanisms of action. Environ. Pollut. 2012, 169, 81–89. [Google Scholar] [CrossRef]
- Pang, C.; Selck, H.; Misra, S.K.; Berhanu, D.; Dybowska, A.; Valsami-Jones, E.; Forbes, V.E. Effects of sediment-associated copper to the deposit-feeding snail, Potamopyrgus antipodarum: A comparison of Cu added in aqueous form or as nano- and micro-CuO particles. Aquat. Toxicol. 2012, 15, 114–122. [Google Scholar]
- Studer, A.M.; Limbach, L.K.; van Duc, L.; Krumeich, F.; Athanassiou, E.K.; Gerber, L.C.; Moch, H.; Stark, W.J. Nanoparticle cytotoxicity depends on intracellular solubility: Comparison of stabilized copper metal and degradable copper oxide nanoparticles. Toxicol. Lett. 2010, 1, 169–174. [Google Scholar]
- Cronholm, P.; Karlsson, H.L.; Hedberg, J.; Lowe, T.A.; Winnberg, L.; Elihn, K.; Wallinder, I.O.; Möller, L. Intracellular uptake and toxicity of Ag and CuO nanoparticles: A comparison between nanoparticles and their corresponding metal ions. Small 2013, 8, 970–982. [Google Scholar]
- Cuillel, M.; Chevallet, M.; Charbonnier, P.; Fauquant, C.; Pignot-Paintrand, I.; Arnaud, J.; Cassio, D.; Michaud-Soret, I.; Mintz, E. Interference of CuO nanoparticles with metal homeostasis in hepatocytes under sub-toxic conditions. Nanoscale 2014, 16, 1707–1715. [Google Scholar]
- Fröhlich, E. Cellular targets and mechanisms in the cytotoxic action of non-biodegradable engineered nanoparticles. J. Curr. Drug Metab. 2013, 14, 976–988. [Google Scholar] [CrossRef]
- Chen, Z.; Meng, H.; Xing, G.; Chen, C.; Zhao, Y.; Jia, G.; Wang, T.; Yuan, H.; Ye, C.; Zhao, F.; et al. Acute toxicological effects of copper nanoparticles in vivo. Toxicol. Lett. 2006, 25, 109–120. [Google Scholar]
- Liao, M.; Liu, H. Gene expression profiling of nephrotoxicity from copper nanoparticles in rats after repeated oral administration. Environ. Toxicol. Pharmacol. 2012, 34, 67–80. [Google Scholar] [CrossRef]
- Pan, X.; Redding, J.E.; Wiley, P.A.; Wen, L.; McConnell, J.S.; Zhang, B. Mutagenicity evaluation of metal oxide nanoparticles by the bacterial reverse mutation assay. Chemosphere 2010, 79, 113–116. [Google Scholar] [CrossRef]
- Song, M.F.; Li, Y.S.; Kasai, H.; Kawai, K. Metal nanoparticle-induced micronuclei and oxidative DNA damage in mice. J. Clin. Biochem. Nutr. 2012, 50, 211–216. [Google Scholar] [CrossRef]
- Gomes, T.; Araújo, O.; Pereira, R.; Almeida, A.C.; Cravo, A.; Bebianno, M.J. Genotoxicity of copper oxide and silver nanoparticles in the mussel Mytilus galloprovincialis. Mar. Environ. Res. 2013, 84, 51–59. [Google Scholar] [CrossRef]
- Alarifi, S.; Ali, D.; Verma, A.; Alakhtani, S.; Ali, B.A. Cytotoxicity and genotoxicity of copper oxide nanoparticles in human skin keratinocytes cells. Int. J. Toxicol. 2013, 32, 296–307. [Google Scholar] [CrossRef]
- Akhtar, M.J.; Kumar, S.; Alhadlaq, H.A.; Alrokayan, S.A.; Abu-Salah, K.M.; Ahamed, M. Dose-dependent genotoxicity of copper oxide nanoparticles stimulated by reactive oxygen species in human lung epithelial cells. Toxicol. Ind. Health. Available online: http://www.ncbi.nlm.nih.gov/pubmed/24311626 (accessed on 5 December 2005). [CrossRef]
- Magaye, R.; Zhao, J.; Bowman, L.; Ding, M. Genotoxicity and carcinogenicity of cobalt-, nickel- and copper-based nanoparticles. Exp. Ther. Med. 2012, 4, 551–561. [Google Scholar]
- Xu, J.; Li, Z.; Xu, P.; Xiao, L.; Yang, Z. Nanosized copper oxide induces apoptosis through oxidative stress in podocytes. Arch. Toxicol. 2013, 87, 1067–1073. [Google Scholar] [CrossRef]
- Sokol, R.J.; Devereaux, M.W.; Traber, M.G.; Shikes, R.H. Copper toxicity and lipid peroxidation in isolated rat hepatocytes: Effect of vitamin E. Pediatr. Res. 1989, 25, 55–62. [Google Scholar] [CrossRef]
- Katsnelson, B.A.; Kuzmin, S.V.; Degtyareva, T.D.; Privalova, L.I.; Soloboyeva, J.I. “Biological prophylaxis”—One of the ways to proceed from the analytical environmental epidemiology to the population health protection. Cent. Eur. J. Occup. Environ. Med. 2008, 14, 41–42. [Google Scholar]
- Suttle, N.F. Control of hepatic copper retention in Texel ram lambs by dietary supplementation with copper antagonists followed by a copper depletion regimen. Anim. Food Sci. Technol. 2012, 11, 194–200. [Google Scholar] [CrossRef]
- Faddah, L.M.; Abdel Baky, N.A.; Al-Rasheed, N.M.; Fatani, A.U.; Atteya, M. Role of quercetin and arginine in ameliorating nano zinc oxide-induced nephrotoxicity in ra. BMC Complement. Altern. Med. 2012, 12, 60. [Google Scholar] [CrossRef]
- IPCS (International Program on Chemical Safety). Environmental Health Criteria 200. – Copper; World Health Organization: Geneva, Switzerland, 1998; p. 360. [Google Scholar]
- Gaforio, J.J.; Serrano, M.J.; Algarra, I.; Ortega, E.; Alvarez de Cienfuegos, G. Phagocytosis of apoptotic cells assessed by flow cytometry using 7-aminoactinomycin D. Cytometry 2002, 49, 8–11. [Google Scholar]
- Canbay, A.; Feldstein, A.E.; Higuchi, H.; Werneburg, N.; Grambihler, A.; Bronk, S.F.; Gores, G.J. Kupffer cell engulfment of apoptotic bodies stimulates death ligand and cytokine expression. Hepatology 2003, 38, 1188–1198. [Google Scholar]
- Sizova, E.; Miroshnikov, S.; Polyakova, V.; Gluschenko, N.; Skalny, A. Copper nanoparticles as modulators of apoptosis and structural changes in tissues. J. Biomater. Nanobiotechnol. 2012, 3, 97–104. [Google Scholar] [CrossRef]
- Cox, D.W. Disorders of copper transport. Br. Med. Bull. 1999, 55, 544–555. [Google Scholar] [CrossRef]
- Vanlandingham, J.W.; Zeidan, N.T.; Levenson, C.W. Copper regulation of neuronal apoptosis use of cDNA array to identify p53-responsive genes in human neurons. FASEB J. 2003, 17, 4–5. [Google Scholar] [CrossRef]
- Pal, A.; Vasishta, R.; Prasad, R. Hepatic and hippocampus iron status is not altered in response to increased serum ceruloplasmin and serum “free” copper in Wistar rat model for non-Wilsonian brain copper toxicosis. Biol. Trace Elem. Res. 2013, 154, 403–411. [Google Scholar] [CrossRef]
- Lee, T.Y.; Liu, M.S.; Huang, L.J.; Lue, S.I.; Lin, L.C.; Kwan, A.L.; Yang, R.C. Bioenergetic failure correlates with autophagy and apoptosis in rat liver following silver nanoparticle intraperitoneally administration. Part. Fibre Toxicol. 2013, 10, 40. [Google Scholar] [CrossRef]
- Geiser, M.; Kreyling, W.G. Deposition and biokinetics of inhaled nanoparticles. Part. Fibre Toxicol. 2010, 7, 2. [Google Scholar] [CrossRef]
- Task Group. ICRP Publication 66: Human respiratory tract model for radiological protection. A report of a Task Group of the International Commission on Radiological Protection. Ann. ICRP 1994, 24, 1–482. [Google Scholar]
- Kreyling, W.G.; Geiser, M. Dosimetry of inhaled nanoparticles. In Nanoparticles in Medicine and Environment, Inhalation and Health Effects; Marijnissen, J.C., Gradon, L., Eds.; Springer: Berlin, Germany, 2009. [Google Scholar]
- Fröhlich, E.; Salar-Behzadi, S. Toxicological assessment of inhaled nanoparticles: Role of in vivo, ex vivo, in vitro, and in silico studies. Int. J. Mol. Sci. 2014, 15, 4795–4822. [Google Scholar] [CrossRef]
- Sadauskas, E.; Wallin, H.; Stolenberg, M.; Vogel, U.; Doering, P.; Larsen., A.; Danscher, G. Kupfer cells are central in the removal of nanoparticles from the organism. Part. Fibre Toxicol. 2007, 4, 10. [Google Scholar] [CrossRef]
- Lasagna-Reeves, C.; Gonzalez-Romero, D.; Barria, M.A.; Olmedo, I.; Clos, A.; Sadagopa Ramanujam, V.M.; Urayama, A.; Vergara, L.; Kogan, M.J.; Soto, C. Bioaccumulation and toxicity of gold nanoparticles after repeated administration in mice. Biochem. Biophys. Res. Commun. 2010, 393, 649–655. [Google Scholar] [CrossRef]
- Rylova, M.L. Methods of Investigating Long-Term Effects of Noxious Environmental Agents in Animal Experiments; Meditsina: Leningrad, Russia, 1964; p. 228. [Google Scholar]
- Abeyemi, O.O.; Yemitan, O.K.; Taiwo, A.E. Neurosedative and muscle-relaxant activities of ethyl acetate extract of Baphianitida nitida AFZEL. J. Ethnopharmacol. 2006, 106, 312–316. [Google Scholar] [CrossRef]
- Fernandez, S.P.; Wasowski, C.; Loscalzo, L.M.; Granger, R.E.; Johnston, G.A.; Paladini, A.C.; Marder, M. Central nervous system depressant action of flavonoid glycoside. Eur. J. Pharmacol. 2006, 539, 168–176. [Google Scholar] [CrossRef]
- Tietz, N.W. Clinical Guide to Laboratory Tests, 3rd ed.; W.B. Saunders Company: Philadelphia, PA, USA, 1995. [Google Scholar]
- Katsnelson, B.A.; Makeyev, O.H.; Kochneva, N.I.; Privalova, L.I.; Degtyareva, T.D.; Bukhantsev, V.A.; Minin, V.V.; Beresneva, O.Y.; Slyshkina, T.V.; Kostyukova, S.V. Testing a set of bioprotectors against the genotoxic effect of a combination of ecotoxicants. Cent. Eur. J. Occup. Environ. Med. 2007, 13, 251–264. [Google Scholar]
- Katsnelson, B.A.; Yeremenko, O.S.; Privalova, L.I.; Makeyev, O.H.; Degtyareva, T.D.; Beresneva, O.Y.; Valamina, I.E.; Minigalieva, I.A.; Sutunkova, M.P.; Kireyeva, E.P.; et al. Toxicity of monazite particulate and its attenuation with a complex of bio-protectors. Med. Lavoro 2009, 100, 455–470. [Google Scholar]
- Morosova, K.I.; Aronova, G.V.; Katsnelson, B.A.; Velichkovski, B.T.; Genkin, A.M.; Elnichnykh, L.N.; Privalova, L.I. On the defensive action of glutamate on the cytotoxicity and fibrogenicity of quartz dust. Br. J. Ind. Med. 1982, 39, 244–252. [Google Scholar]
- Privalova, L.I.; Katsnelson, B.A.; Sutunkova, M.P.; Valamina, I.E.; Beresneva, O.Y.; Degtyareva, T.D.; Yeremenko, O.S. Attenuation of some adverse health effects of chrysotile asbestos with a bioprotective complex in animal experiments. Cent. Eur. J. Occup. Environ. Med. 2007, 13, 265–276. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Privalova, L.I.; Katsnelson, B.A.; Loginova, N.V.; Gurvich, V.B.; Shur, V.Y.; Valamina, I.E.; Makeyev, O.H.; Sutunkova, M.P.; Minigalieva, I.A.; Kireyeva, E.P.; et al. Subchronic Toxicity of Copper Oxide Nanoparticles and Its Attenuation with the Help of a Combination of Bioprotectors. Int. J. Mol. Sci. 2014, 15, 12379-12406. https://doi.org/10.3390/ijms150712379
Privalova LI, Katsnelson BA, Loginova NV, Gurvich VB, Shur VY, Valamina IE, Makeyev OH, Sutunkova MP, Minigalieva IA, Kireyeva EP, et al. Subchronic Toxicity of Copper Oxide Nanoparticles and Its Attenuation with the Help of a Combination of Bioprotectors. International Journal of Molecular Sciences. 2014; 15(7):12379-12406. https://doi.org/10.3390/ijms150712379
Chicago/Turabian StylePrivalova, Larisa I., Boris A. Katsnelson, Nadezhda V. Loginova, Vladimir B. Gurvich, Vladimir Y. Shur, Irene E. Valamina, Oleg H. Makeyev, Marina P. Sutunkova, Ilzira A. Minigalieva, Ekaterina P. Kireyeva, and et al. 2014. "Subchronic Toxicity of Copper Oxide Nanoparticles and Its Attenuation with the Help of a Combination of Bioprotectors" International Journal of Molecular Sciences 15, no. 7: 12379-12406. https://doi.org/10.3390/ijms150712379