Coexpression and Secretion of Endoglucanase and Phytase Genes in Lactobacillus reuteri

Abstract
:1. Introduction
2. Results
2.1. Construction of Endoglucanase–Phytase-Coexpressing Plasmids


2.2. Coexpression of Endoglucanase and Phytase Genes in L. reuteri XC1


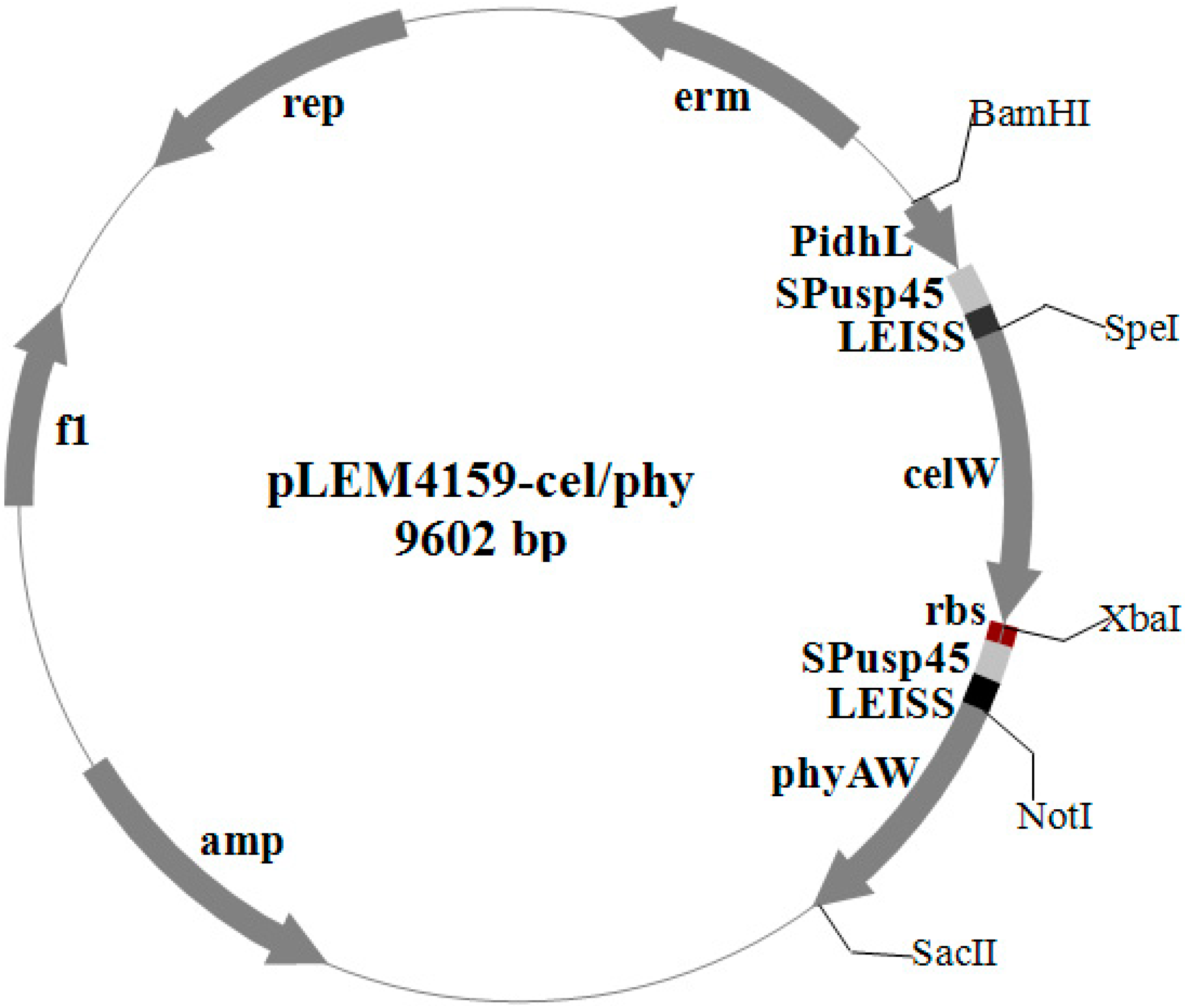{kind=link}
{kind=link}
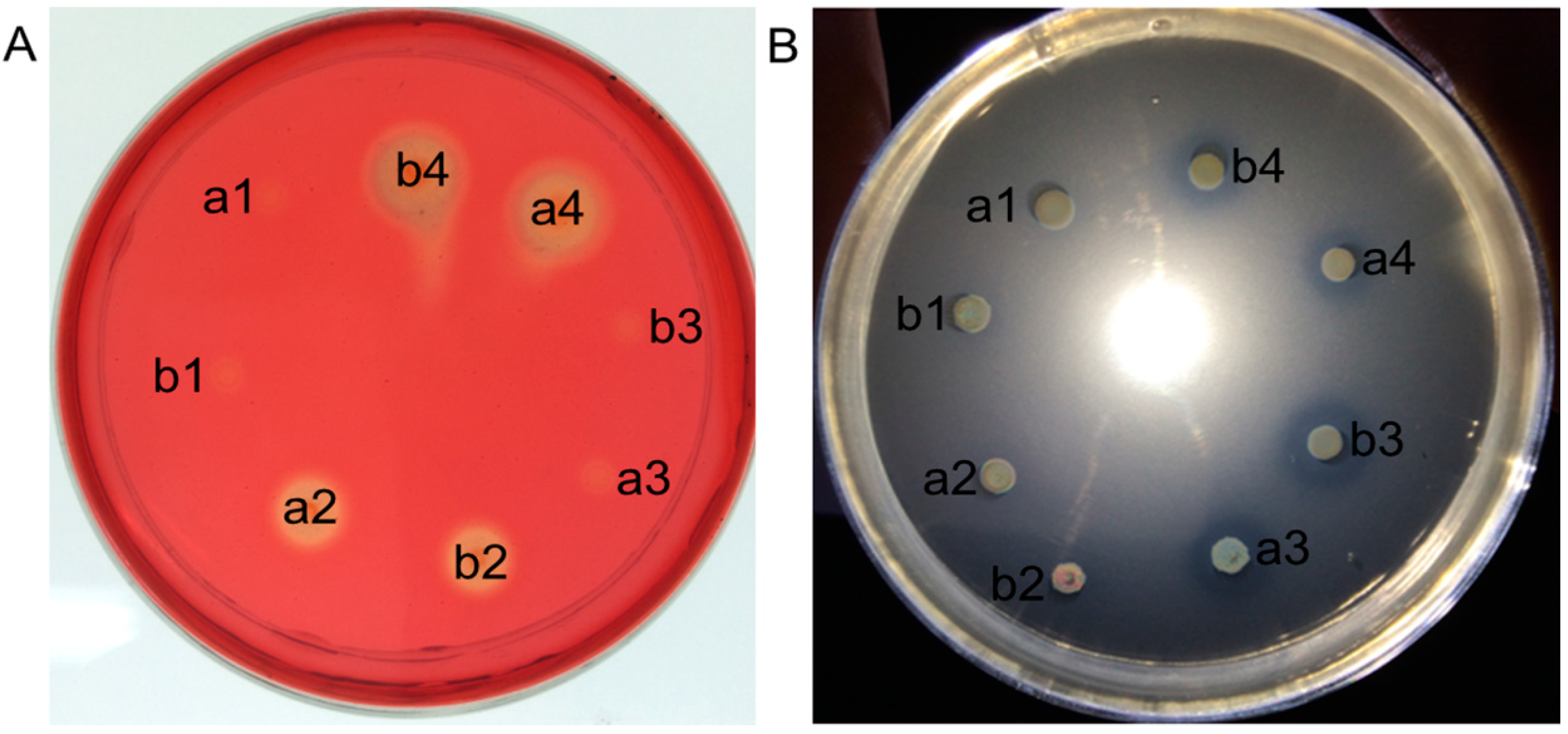{kind=link}
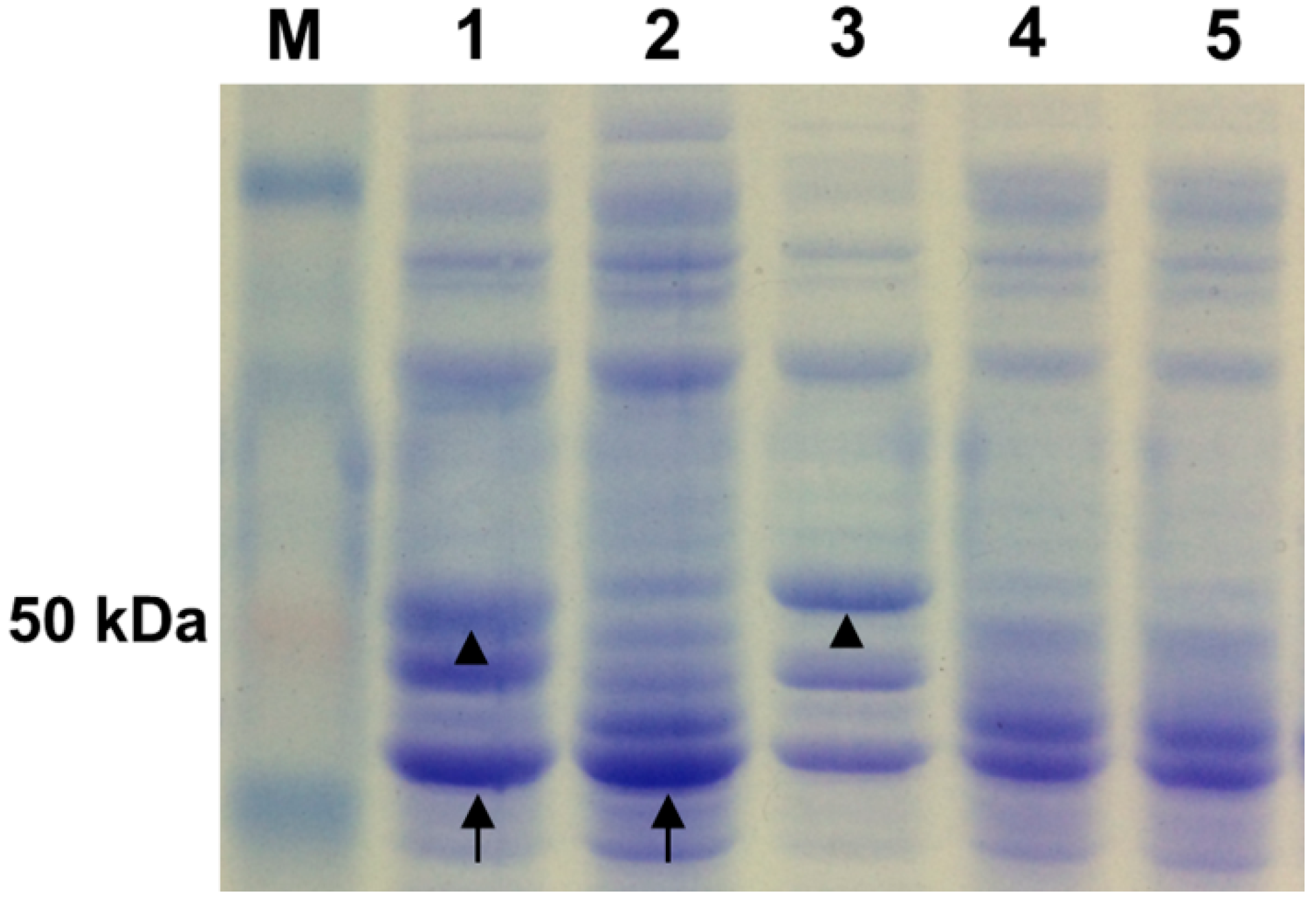{kind=link}
| Strain | Enzyme Activity (U/mL) 1 | |||
|---|---|---|---|---|
| Endoglucanase | Phytase | |||
| Intracellular | Extracellular | Intracellular | Extracellular | |
| L. reuteri pLEM4156 | Nd 2 | Nd | Nd | Nd |
| L. reuteri pLEM4157 (cel) | 0.37 ± 0.09 | 0.96 ± 0.08 | Nd | Nd |
| L. reuteri pLEM4158 (phy) | Nd | Nd | 0.26 ± 0.14 | 0.51 ± 0.13 |
| L. reuteri pLEM4159-cel/phy | 0.15 ± 0.06 | 0.68 ± 0. 17 | 0.14 ± 0.16 | 0.42 ± 0.05 |

2.3. Growth Performance of Broilers
| Performance | Control Group 1 | L. reuteri Group | SEM | p Value | ||||
|---|---|---|---|---|---|---|---|---|
| NC | PC | pLEM4156 | pLEM4157 (cel) | pLEM4158 (phy) | pLEM4159-cel/phy | |||
| days (1–21) | ||||||||
| BWG, g/bird/day | 27.85 | 28.14 | 27.83 | 27.57 | 28.23 | 27.32 | 0.173 | 0.697 |
| Feed intake, g/bird/day | 39.36 | 39.73 | 39.26 | 39.33 | 39.46 | 38.75 | 0.220 | 0.900 |
| FCR | 1.42 | 1.41 | 1.41 | 1.43 | 1.40 | 1.42 | 0.006 | 0.933 |
| days (22–42) | ||||||||
| BWG, g/bird/day | 57.47 | 62.21 | 61.46 | 59.22 | 61.13 | 60.66 | 0.854 | 0.664 |
| Feed intake, g/bird/day | 115.19 | 115.77 | 117.47 | 111.50 | 112.24 | 111.07 | 1.207 | 0.585 |
| FCR | 2.01 a | 1.86 b | 1.93 a,b | 1.89 a,b | 1.83 b | 1.84 b | 0.019 | 0.049 |
| days (1–42) | ||||||||
| BWG, g/bird/d | 42.76 | 45.31 | 44.74 | 43.48 | 44.77 | 44.06 | 0.439 | 0.598 |
| Feed intake, g/bird/d | 77.28 | 77.75 | 78.37 | 75.42 | 75.85 | 74.91 | 0.614 | 0.532 |
| FCR | 1.81 a | 1.72 b | 1.76 a,b | 1.74 b | 1.69 b | 1.70 b | 0.011 | 0.015 |
2.4. Tibial Parameters of Broiler Chickens
| Tibia | Control Group 1 | L. reuteri Group | SEM | p Value | ||||
|---|---|---|---|---|---|---|---|---|
| NC | PC | pLEM4156 | pLEM4157 (cel) | pLEM4158 (phy) | pLEM4159-cel/phy | |||
| day 21 | ||||||||
| Weight, g | 0.86 | 0.71 | 0.87 | 0.80 | 0.78 | 0.78 | 0.021 | 0.317 |
| Length, cm | 4.41 | 4.31 | 4.52 | 4.36 | 4.40 | 4.30 | 0.036 | 0.667 |
| Ash, % | 48.47 a | 52.31 c | 49.15 a | 49.46 a,b | 50.92 a,b,c | 51.71 b,c | 0.406 | 0.016 |
| P, % | 9.70 a | 10.58 b | 9.77 | 9.75 a | 9.91 a | 10.13 a,b | 0.090 | 0.030 |
| Ca, % | 15.15 a | 17.74 b | 16.84 a,b | 17.82 b | 17.14 a,b | 18.99 b | 0.373 | 0.044 |
| day 42 | ||||||||
| Weight, g | 1.99 | 2.31 | 2.45 | 1.98 | 2.11 | 2.13 | 0.060 | 0.128 |
| Length, cm | 6.40 | 6.74 | 6.71 | 6.31 | 6.36 | 6.33 | 0.068 | 0.128 |
| Ash, % | 52.18 | 50.54 | 52.36 | 50.89 | 52.94 | 50.84 | 0.269 | 0.376 |
| P, % | 9.91 a | 10.20 a,b | 10.10 a,b | 10.29 a,b | 10.66 b | 10.20 a,b | 0.081 | 0.026 |
| Ca, % | 17.27 | 18.09 | 16.28 | 20.00 | 17.68 | 17.58 | 0.586 | 0.473 |
2.5. Microflora Population in the Cecum
| Groups | Control Group 1 | L. reuteri Group | SEM | p Value | ||||
|---|---|---|---|---|---|---|---|---|
| NC | PC | pLEM4156 | pLEM4157 (cel) | pLEM4158 (phy) | pLEM4159-cel/phy | |||
| day 21 | ||||||||
| Escherichia coli | 5.44 a,b | 6.12 b | 4.84 a,b | 5.26 a,b | 4.70 a,b | 2.99 a | 0.347 | 0.05 |
| Bifidobacterium spp. | 1.50 | 1.45 | 1.99 | 2.19 | 1.79 | 2.24 | 0.116 | 0.231 |
| Bacteroides vulgatus | 2.50 | 3.19 | 2.31 | 2.25 | 2.26 | 2.07 | 0.159 | 0.493 |
| Veillonella spp. | 1.52 a,b | 1.82 b | 0.96 a,b | 0.45 a | 0.65 a,b | 0.89 a,b | 0.170 | 0.048 |
| Clostridium IV | 6.34 | 6.29 | 5.69 | 4.94 | 5.66 | 6.20 | 0.237 | 0.481 |
| Lactobacillus spp. | 3.74 | 4.44 | 5.30 | 4.66 | 5.18 | 5.42 | 0.251 | 0.322 |
| Enterococcus faecalis | 1.42 | 2.20 | 1.26 | 2.47 | 1.23 | 1.91 | 0.241 | 0.594 |
| day 42 | ||||||||
| Escherichia coli | 3.68 | 3.23 | 4.51 | 3.02 | 3.18 | 4.04 | 0.224 | 0.422 |
| Bifidobacterium spp. | 0.68 | 0.72 | 1.62 | 1.11 | 0.94 | 1.45 | 0.135 | 0.295 |
| Bacteroides vulgatus | 3.39 | 3.97 | 2.81 | 2.51 | 3.19 | 4.04 | 0.331 | 0.761 |
| Veillonella spp. | 0.50 | 0.66 | 0.68 | 1.57 | 1.56 | 1.10 | 0.174 | 0.291 |
| Clostridium IV | 4.47 | 4.39 | 5.91 | 3.91 | 4.54 | 5.57 | 0.300 | 0.432 |
| Lactobacillus spp. | 3.59 | 3.04 | 4.16 | 4.31 | 4.31 | 4.97 | 0.295 | 0.531 |
| Enterococcus faecalis | 0.60 | 0.39 | 0.84 | 1.15 | 1.25 | 1.05 | 0.193 | 0.802 |
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains, Plasmids, and Growth Conditions
4.2. DNA Isolation and Manipulation
4.3. Construction of Coexpression Vector
4.4. Construction of Endoglucanase and Phytase Coexpression Plasmid
4.5. Transformation of Plasmid DNA
4.6. Determination of Endoglucanase and Phytase Activity
4.7. Sodium Dodecyl Sulfate Polyacrylamide Gel Electrophoresis (SDS-PAGE) Analysis
4.8. Broiler Chickens, Diet and Growth Trail
4.9. Determination of Tibia Parameters
4.10. Analysis of Microbial Distribution in the Cecum by Real-Time Quantitative PCR
4.11. Statistical Analysis
5. Conclusions
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Yin, Q.Q.; Zheng, Q.H.; Kang, X.T. Biochemical characteristics of phytases from fungi and the transformed microorganism. Anim. Feed Sci. Technol. 2007, 132, 341–350. [Google Scholar] [CrossRef]
- Liu, J.R.; Yu, B.; Liu, F.H.; Cheng, K.J.; Zhao, X. Expression of rumen microbial fibrolytic enzyme genes in probiotic Lactobacillus reuteri. Appl. Environ. Microbiol. 2005, 71, 6769–6775. [Google Scholar] [CrossRef]
- Selle, P.H.; Ravindran, V.; Caldwell, R.A.; Bryden, W.L. Phytate and phytase: Consequences for protein utilisation. Nutr. Res. Rev. 2000, 13, 255–278. [Google Scholar] [CrossRef]
- Choct, M.; Annison, G. Anti-nutritive effect of wheat pentosans in broiler chickens: Roles of viscosity and gut microflora. Br. Poult. Sci. 1992, 33, 821–834. [Google Scholar] [CrossRef]
- Bedford, M.R.; Schulze, H. Exogenous enzymes for pigs and poultry. Nutr. Res. Rev. 1998, 11, 91–114. [Google Scholar] [CrossRef]
- Correll, D.L. Phosphorus: A rate limiting nutrient in surface waters. Poult. Sci. 1999, 78, 674–682. [Google Scholar] [CrossRef]
- Choi, Y.M.; Suh, H.J.; Kim, J.M. Purification and properties of extracellular phytase from Bacillus sp. KHU-10. J. Protein Chem. 2001, 20, 287–292. [Google Scholar] [CrossRef]
- Hall, J.; Ali, S.; Surani, M.A.; Hazlewood, G.P.; Clark, A.J.; Simons, J.P.; Hirst, B.H.; Gilbert, H.J. Manipulation of the repertoire of digestive enzymes secreted into the gastrointestinal tract of transgenic mice. Nat. Biotechnol. 1993, 11, 376–379. [Google Scholar] [CrossRef]
- Knowlton, K.F.; Parsons, C.M.; Cobb, C.W.; Wilson, K.F. Exogenous phytase plus cellulase and phosphorus excretion in lactating dairy cows. Prof. Anim. Sci. 2005, 21, 212–216. [Google Scholar]
- Waldroup, P.W.; Kersey, J.H.; Saleh, E.A.; Fritts, C.A.; Yan, F.; Stilborn, H.L.; Crum, R.C., Jr.; Raboy, V. Nonphytate phosphorus requirement and phosphorus excretion of broiler chicks fed diets composed of normal or high available phosphate corn with and without microbial phytase. Poult. Sci. 2000, 79, 1451–1459. [Google Scholar] [CrossRef]
- Cromwell, G.L.; Stahly, T.S.; Coffey, R.D.; Monegue, H.J.; Randolph, J.H. Efficacy of phytase in improving the bioavailability of phosphorus in soybean meal and corn-soybean meal diets for pigs. J. Anim. Sci. 1993, 71, 1831–1840. [Google Scholar]
- Simons, P.C.M.; Versteegh, H.A.J.; Jongbloed, A.W.; Kemme, P.A.; Slump, P.; Bos, K.D.; Wolters, M.G.E.; Beudeker, R.F.; Verschoor, G.J. Improvement of phosphorus availability by microbial phytase in broilers and pigs. Br. J. Nutr. 1990, 64, 525–540. [Google Scholar] [CrossRef]
- Mathlouthi, N.; Juin, H.; Larbier, M. Effect of xylanase and β-glucanase supplementation of wheat-or wheat-and barley-based diets on the performance of male turkeys. Br. Poult. Sci. 2003, 44, 291–298. [Google Scholar] [CrossRef]
- Mathlouthi, N.; Mohamed, M.A.; Larbier, M. Effect of enzyme preparation containing xylanase and β-glucanase on performance of laying hens fed wheat/barley-or maize/soybean meal-based diets. Br. Poult. Sci. 2003, 44, 60–66. [Google Scholar] [CrossRef]
- Bates, E.E.; Gilbert, H.J.; Hazlewood, G.P.; Huckle, J.; Laurie, J.I.; Mann, S.P. Expression of a Clostridium thermocellum endoglucanase gene in Lactobacillus plantarum. Appl. Environ. Microbiol. 1989, 55, 2095–2097. [Google Scholar]
- Liu, J.R.; Yu, B.; Zhao, X.; Cheng, K.J. Coexpression of rumen microbial β-glucanase and xylanase genes in Lactobacillus reuteri. Appl. Microbiol. Biotechnol. 2007, 77, 117–124. [Google Scholar] [CrossRef]
- Cho, J.S.; Choi, Y.J.; Chung, D.K. Expression of Clostridium thermocellum endoglucanase gene in Lactobacillus gasseri and Lactobacillus johnsonii and characterization of the genetically modified probiotic lactobacilli. Curr. Microbiol. 2000, 40, 257–263. [Google Scholar] [CrossRef]
- Zegers, N.D.; Kluter, E.; van der Stap, H.; van Dura, E.; van Dalen, P.; Shaw, M.; Baillie, L. Expression of the protective antigen of Bacillus anthracis by Lactobacillus casei: Towards the development of an oral vaccine against anthrax. J. Appl. Microbiol. 2001, 87, 309–314. [Google Scholar]
- Bateup, J.M.; McConnell, M.A.; Jenkinson, H.F.; Tannock, G.W. Comparison of Lactobacillus strains with respect to bile salt hydrolase activity, colonization of the gastrointestinal tract, and growth rate of the murine host. Appl. Environ. Microbiol. 1995, 61, 1147–1149. [Google Scholar]
- Zuo, R.; Chang, J.; Yin, Q.; Chen, L.; Chen, Q.; Yang, X.; Zheng, Q.; Ren, G.; Feng, H. Phytase gene expression in Lactobacillus and analysis of its biochemical characteristics. Microbiol. Res. 2010, 165, 329–335. [Google Scholar] [CrossRef]
- Kerovuo, J.; Tynkkynen, S. Expression of Bacillus subtilis phytase in Lactobacillus plantarum 755. Lett. Appl. Microbiol. 2001, 30, 325–329. [Google Scholar] [CrossRef]
- Pérez-Arellano, I.; Pérez-Martı́nez, G. Optimization of the green fluorescent protein (GFP) expression from a lactose-inducible promoter in Lactobacillus casei. FEMS Microbiol. Lett. 2006, 222, 123–127. [Google Scholar] [CrossRef]
- Yu, Q.H.; Dong, S.M.; Zhu, W.Y.; Yang, Q. Use of green fluorescent protein to monitor Lactobacillus in the gastro-intestinal tract of chicken. FEMS Microbiol. Lett. 2007, 275, 207–213. [Google Scholar] [CrossRef]
- Wells, J.M.; Mercenier, A. Mucosal delivery of therapeutic and prophylactic molecules using lactic acid bacteria. Nat. Rev. Microbiol. 2008, 6, 349–362. [Google Scholar] [CrossRef]
- Rodrıguez, J.M.; Martınez, M.I.; Horn, N.; Dodd, H.M. Heterologous production of bacteriocins by lactic acid bacteria. Int. J. Food Microbiol. 2003, 80, 101–116. [Google Scholar] [CrossRef]
- Liu, J.R.; Lai, S.F.; Yu, B. Evaluation of an intestinal Lactobacillus reuteri strain expressing rumen fungal xylanase as a probiotic for broiler chickens fed on a wheat-based diet. Br. Poult. Sci. 2007, 48, 507–514. [Google Scholar] [CrossRef]
- Sieo, C.C.; Abdullah, N.; Tan, W.S.; Ho, Y.W. In vivo study on the persistence of transformed β-glucanase-producing Lactobacillus strains in the gastrointestinal tract of chickens. J. Anim. Feed Sci. 2006, 15, 261. [Google Scholar]
- Stephenson, D.P.; Moore, R.J.; Allison, G.E. Transformation of, and heterologous protein expression in, Lactobacillus agilis and Lactobacillus vaginalis isolates from the chicken gastrointestinal tract. Appl. Environ. Microbiol. 2011, 77, 220–228. [Google Scholar] [CrossRef]
- Gory, L.; Montel, M.C.; Zagorec, M. Use of green fluorescent protein to monitor Lactobacillus sakei in fermented meat products. FEMS Microbiol. Lett. 2006, 194, 127–133. [Google Scholar] [CrossRef]
- Boot, H.J.; Kolen, C.P.; Andreadaki, F.J.; Leer, R.J.; Pouwels, P.H. The Lactobacillus acidophilus S-layer protein gene expression site comprises two consensus promoter sequences, one of which directs transcription of stable mRNA. J. Bacteriol. 1996, 178, 5388–5394. [Google Scholar]
- Djordjevic, G.; Bojovic, B.; Banina, A.; Topisirovic, L. Cloning of promoter-like sequences from Lactobacillus paracasei subsp. paracasei CG11 and their expression in Escherichia coli, Lactococcus lactis, and Lactobacillus reuteri. Can. J. Microbiol. 1994, 40, 1043–1050. [Google Scholar] [CrossRef]
- Chang, T.L.Y.; Chang, C.H.; Simpson, D.A.; Xu, Q.; Martin, P.K.; Lagenaur, L.A.; Schoolnik, G.K.; Ho, D.D.; Hillier, S.L.; Holodniy, M. Inhibition of HIV infectivity by a natural human isolate of Lactobacillus jensenii engineered to express functional two-domain CD4. Proc. Natl. Acad. Sci. USA 2003, 100, 11672–11677. [Google Scholar] [CrossRef]
- Le Loir, Y.; Nouaille, S.; Commissaire, J.; Bretigny, L.; Gruss, A.; Langella, P. Signal peptide and propeptide optimization for heterologous protein secretion in Lactococcus lactis. Appl. Environ. Microbiol. 2001, 67, 4119–4127. [Google Scholar]
- Asseldonk, M.; Vos, W.M.; Simons, G. Functional analysis of the Lactococcus lactis usp45 secretion signal in the secretion of a homologous proteinase and a heterologous α-amylase. Mol. Gen. Genet. 1993, 240, 428–434. [Google Scholar] [CrossRef]
- Steidler, L.; Hans, W.; Schotte, L.; Neirynck, S.; Obermeier, F.; Falk, W.; Fiers, W.; Remaut, E. Treatment of murine colitis by Lactococcus lactis secreting interleukin-10. Science 2000, 289, 1352–1355. [Google Scholar] [CrossRef]
- Mercenier, A.; Pouwels, P.H.; Chassy, B.M. Genetics and Biotechnology of Lactic Acid Bacteria; Springer Netherlands: Blackie Academic and Professional, Glasgow, UK, 1994; pp. 252–293. [Google Scholar]
- Hols, P.; Ferain, T.; Garmyn, D.; Bernard, N.; Delcour, J. Use of homologous expression-secretion signals and vector-free stable chromosomal integration in engineering of Lactobacillus plantarum for alpha-amylase and levanase expression. Appl. Environ. Microbiol. 1994, 60, 1401–1413. [Google Scholar]
- Savijoki, K.; Kahala, M.; Palva, A. High level heterologous protein production in Lactococcus and Lactobacillus using a new secretion system based on the Lactobacillus brevis S-layer signals. Gene 1997, 186, 255–262. [Google Scholar] [CrossRef]
- Li, W.; Yang, M.M.; Zhang, G.Q.; He, W.L.; Li, Y.X.; Chen, Y.L. Electrotransformation and expression of Cellulase genes in wild-type Lactobacillus reuteri. J. Mol. Microbiol. Biotechnol. 2012, 22, 228–234. [Google Scholar] [CrossRef]
- Heng, N.C.; Jenkinson, H.F.; Tannock, G.W. Cloning and expression of an endo-1, 3–1, 4-β-glucanase gene from Bacillus macerans in Lactobacillus reuteri. Appl. Environ. Microbiol. 1997, 63, 3336–3340. [Google Scholar]
- Selle, P.H.; Ravindran, V. Microbial phytase in poultry nutrition. Anim. Feed Sci. Technol. 2007, 135, 1–41. [Google Scholar] [CrossRef]
- Slominski, B.A. Recent advances in research on enzymes for poultry diets. Poult. Sci. 2011, 90, 2013–2023. [Google Scholar] [CrossRef]
- Yu, B.; Liu, J.R.; Hsiao, F.S.; Chiou, P.W.S. Evaluation of Lactobacillus reuteri Pg4 strain expressing heterologous β-glucanase as a probiotic in poultry diets based on barley. Anim. Feed Sci. Technol. 2008, 141, 82–91. [Google Scholar] [CrossRef]
- Jin, L.Z.; Ho, Y.W.; Abdullah, N.; Jalaludin, S. Growth performance, intestinal microbial populations, and serum cholesterol of broilers fed diets containing Lactobacillus cultures. Poult. Sci. 1998, 77, 1259–1265. [Google Scholar] [CrossRef]
- Askelson, T.E.; Campasino, A.; Lee, J.T.; Duong, T. Evaluation of phytate-degrading Lactobacillus culture administration to broiler chickens. Appl. Environ. Microbiol. 2014, 80, 943–950. [Google Scholar] [CrossRef]
- Watkins, B.A.; Miller, B.F. Competitive gut exclusion of avian pathogens by Lactobacillus acidophilus in gnotobiotic chicks. Poult. Sci. 1983, 62, 1772–1779. [Google Scholar] [CrossRef]
- Perney, K.M.; Cantor, A.H.; Straw, M.L.; Herkelman, K.L. The effect of dietary phytase on growth performance and phosphorus utilization of broiler chicks. Poult. Sci. 1993, 72, 2106–2114. [Google Scholar] [CrossRef]
- Leeson, S.; Namkung, H.; Cottrill, M.; Forsberg, C.W. Efficacy of new bacterial phytase in poultry diets. Can. J. Anim. Sci. 2000, 80, 527–528. [Google Scholar] [CrossRef]
- Ahmad, T.; Rasool, S.; Sarwar, M.; Haq, A.; Hasan, Z. Effect of microbial phytase produced from a fungus Aspergillus niger on bioavailability of phosphorus and calcium in broiler chickens. Anim. Feed Sci. Technol. 2000, 83, 103–114. [Google Scholar] [CrossRef]
- Sebastian, S.; Touchburn, S.P.; Chavez, E.R. Implications of phytic acid and supplemental microbial phytase in poultry nutrition: A review. Worlds Poult. Sci. J. 1998, 54, 27–47. [Google Scholar] [CrossRef]
- Gong, J.; Si, W.; Forster, R.J.; Huang, R.; Yu, H.; Yin, Y.; Yang, C.; Han, Y. 16S rRNA gene-based analysis of mucosa-associated bacterial community and phylogeny in the chicken gastrointestinal tracts: From crops to ceca. FEMS Microbiol. Ecol. 2007, 59, 147–157. [Google Scholar] [CrossRef]
- Bazzocchi, G.; Gionchetti, P.; Almerigi, P.F.; Amadini, C.; Campieri, M. Intestinal microflora and oral bacteriotherapy in irritable bowel syndrome. Dig. Liver Dis. 2002, 34, S48–S53. [Google Scholar] [CrossRef]
- Mathlouthi, N.; Mallet, S.; Saulnier, L.; Quemener, B.; Larbier, M. Effects of xylanase and β-glucanase addition on performance, nutrient digestibility, and physico-chemical conditions in the small intestine contents and caecal microflora of broiler chickens fed a wheat and barley-based diet. Anim. Res. 2002, 51, 395–406. [Google Scholar] [CrossRef]
- Djouzi, Z.; Andlueux, C. Compared effects of three oligosaccharides on metabolism of intestinal microflora in rats inoculated with a human faecal flora. Br. J. Nutr. 1997, 78, 313–324. [Google Scholar] [CrossRef]
- Sambrook, J.; Russell, D.W. Molecular Cloning: A Laboratory Manual, 3nd ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, New York, NY, USA, 2001. [Google Scholar]
- Serror, P.; Sasaki, T.; Ehrlich, S.D.; Maguin, E. Electrotransformation of Lactobacillus delbrueckii subsp. bulgaricus and L. delbrueckii subsp. lactis with various plasmids. Appl. Environ. Microbiol. 2002, 68, 46–52. [Google Scholar] [CrossRef]
- Walker, D.C.; Klaenhammer, T.R. Isolation of a novel IS3 group insertion element and construction of an integration vector for Lactobacillus spp. J. Bacteriol. 1994, 176, 5330–5340. [Google Scholar]
- Teather, R.M.; Wood, P.J. Use of Congo red-polysaccharide interactions in enumeration and characterization of cellulolytic bacteria from the bovine rumen. Appl. Environ. Microbiol. 1982, 43, 777–780. [Google Scholar]
- Piard, J.C.; Hautefort, I.; Fischetti, V.A.; Ehrlich, S.D.; Fons, M.; Gruss, A. Cell wall anchoring of the Streptococcus pyogenes M6 protein in various lactic acid bacteria. J. Bacteriol. 1997, 179, 3068–3072. [Google Scholar]
- Mappley, L.J.; Tchórzewska, M.A.; Nunez, A.; Woodward, M.J.; Bramley, P.M.; La Ragione, R.M. Oral treatment of chickens with Lactobacillus reuteri LM1 reduces Brachyspira pilosicoli-induced pathology. J. Med. Microbiol. 2013, 62, 287–296. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wang, L.; Yang, Y.; Cai, B.; Cao, P.; Yang, M.; Chen, Y. Coexpression and Secretion of Endoglucanase and Phytase Genes in Lactobacillus reuteri. Int. J. Mol. Sci. 2014, 15, 12842-12860. https://doi.org/10.3390/ijms150712842
Wang L, Yang Y, Cai B, Cao P, Yang M, Chen Y. Coexpression and Secretion of Endoglucanase and Phytase Genes in Lactobacillus reuteri. International Journal of Molecular Sciences. 2014; 15(7):12842-12860. https://doi.org/10.3390/ijms150712842
Chicago/Turabian StyleWang, Lei, Yuxin Yang, Bei Cai, Pinghua Cao, Mingming Yang, and Yulin Chen. 2014. "Coexpression and Secretion of Endoglucanase and Phytase Genes in Lactobacillus reuteri" International Journal of Molecular Sciences 15, no. 7: 12842-12860. https://doi.org/10.3390/ijms150712842