
Ecotoxicogenomic Approaches for Understanding Molecular Mechanisms of Environmental Chemical Toxicity Using Aquatic Invertebrate, Daphnia Model Organism

Abstract
:1. Introduction


1.1. Practice of Ecotoxicogenomics in Genomic or Transcriptomic Responses

1.2. Practice of Ecotoxicogenomics in Proteomic Response
1.3. Practice of Ecotoxicogenomics in Metabolomic Response
1.4. Practice of Ecotoxicogenomics in Epigenetic Response
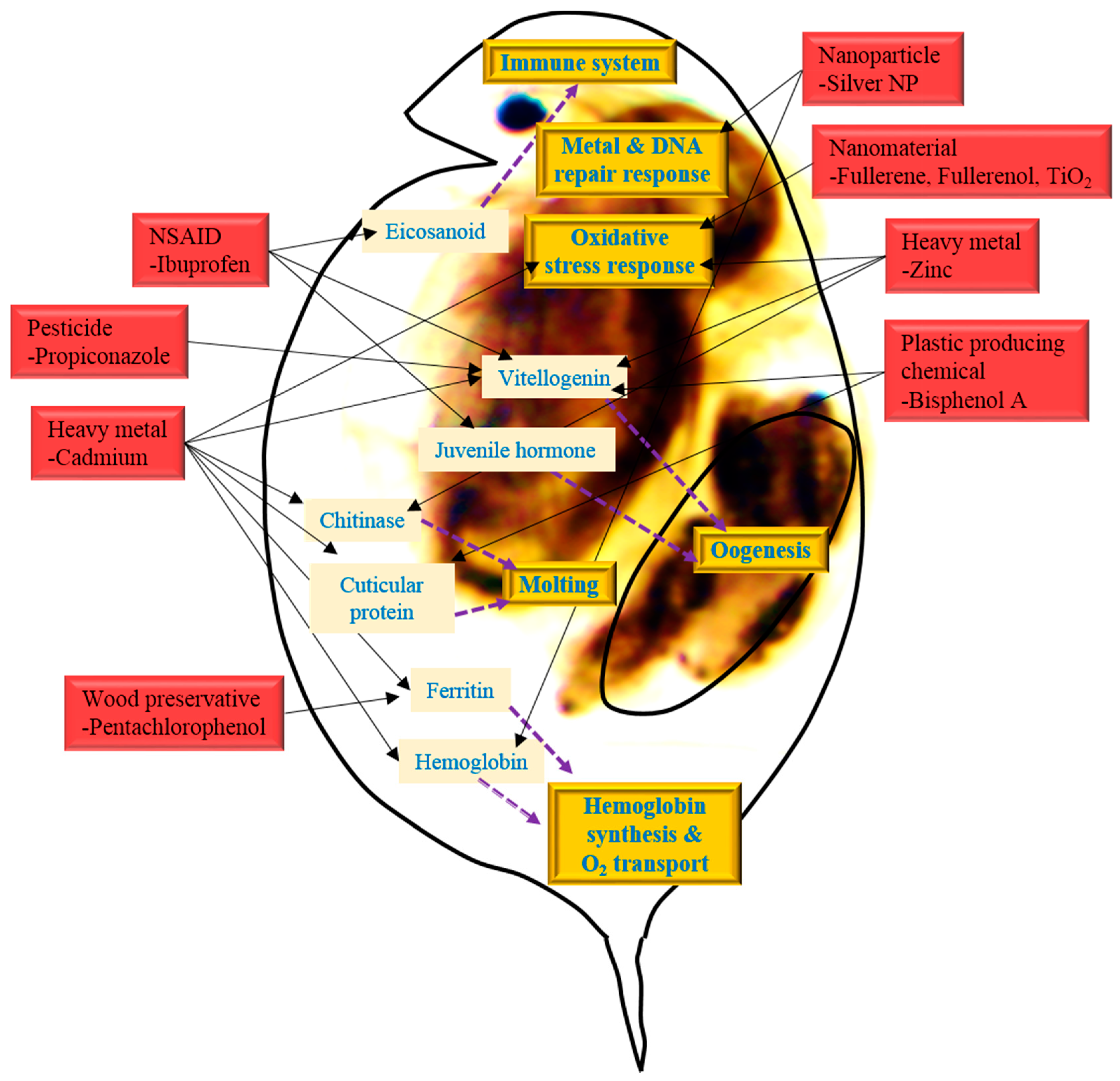
2. Suggestion of Promising Biomarkers of Environmental Toxicity or Exposure

3. Adverse Outcomes from Environmental Stressor Exposure

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chemicals | Species | Chemical Exposure | Effect | Reference |
|---|---|---|---|---|
| EDCs | Daphnia magna | Nonylphenol at 0.024 mg/L for 48 h | Abnormal proportion of juveniles | [119] |
| Daphnia magna | Juvenile hormone at 330 ng/L for 21 days | Production of male offspring | [121] | |
| Ceriodaphnia dubia | Styrene at 0.04–1.7 μg/L for 7 days | Mortality and reduced fertility | [120] | |
| Daphnia magna | Bisphenol A at 6.67 and 10 mg/L for 21 days | Reduced offspring production | [42] | |
| Heavy metals | Daphnia magna | Cadmium at 6, 20, and 37 μg/L for 24 h | Reduced survival and somatic growth | [39] |
| Daphnia magna | Cadmium chloride at 71 μg/L for 24 h | Increased mortality, reduced survival, depleted glutathione level, and induced oxidative stress | [75] | |
| Ceriodaphnia dubia and Daphnia carinata | Copper, lead, zinc (at 1.3, 1.1, 13.0 mg/L, respectively) for 48 h or 7 days | Increased mortality and reduced reproduction ability | [123] | |
| Organic compounds | Daphnia pulex | Organic selenium at 0.025 mg/L for 48 h | Death, immobility, and molting delay | [122] |
| Daphnia magna | Propiconazole (pesticide) at 1 μg/L for 4 and 8 days | Impaired adult growth, decreased offspring development, impaired oocyte maturation, and interrupted resulting reproduction | [41] | |
| Daphnia magna | Fenvalerate (insecticide) at 0.6 μg/L for 24 h | Increased mortality, reduced survival, increased arginine phosphate level, and disrupted amino sugar metabolism | [75] | |
| Daphnia magna | Uncoupler of oxidative phosphorylation 2,4-dinitrophenol at 1.5 mg/L for 24 h | Increased mortality, reduced survival, and increased arginine phosphate level | [75] | |
| Daphnia magna | Alkylpolyglucosides (GCP 650, GCP 600, GCP 215) at IC50 (29, 14, 111 mg/L, respectively) for 24 h | Increased immobility | [124] | |
| Drugs | Daphnia magna | Ibuprofen (non-steroidal anti-inflammatory drug or NSAID) at 20, 40, 80 mg/L for 8 days | Reduced fecundity and arrested early embryogenesis | [104] |
| Daphnia magna | Beta-blocker anti-hypertension drug propanolol at 1.4 mg/L for 24 h | Increased mortality, reduced survival, and disrupted fatty acid metabolism and eicosanoid biosynthesis | [75] | |
| Daphnia magna | Mefenamicacid at EC50 (17.16 mg/L) for 48 h and 1 mg/L for 21 days | Increased immobility and reduced offspring production | [125] | |
| Nanoparticles | Daphnia magna | Silver nanoparticles with surface coating at LC50 (0.88 μg/L) for 48 h | Increased mortality and reduced survival | [126] |
| Daphnia magna | Polyvinylpyrrolidone-coated silver nanoparticles at LC50 (0.18 mg/L) for 24 h | Increased mortality, reduced survival, disrupted proteolysis and cell cycle | [44] | |
| Daphnia magna | Coated silver nanoparticles (Ag-GAs, Ag-PEGs, and Ag-PVPs) at LC50 (3.41, 3.16, 14.81 μg/L, respectively) for 48 h | Increased mortality | [127] | |
| Daphnia magna | Collargol (protein-coated nano Ag) and AgNO3 nanoparticles at EC50 (20–27 ppb) for 48 h | Increased immobility | [128] | |
| Daphnia magna | Ag and CuO nanoparticles at EC50 (3.8 and 2.6 mg/L, respectively) for 24 h | Increased immobility | [129] | |
| Daphnia magna | Titanium dioxide nanoparticles at 0.01–10 mg/L for 48 h | Reduced survival offspring production, and digestion ability | [130] | |
| Daphnia magna | Nano and bulk titanium dioxide at 20 g/L for 48 h | Undetectable toxicity | [131] | |
| Daphnia magna | Bulk CuO, nano CuO and CuSO4 at L(E)C50 (165, 3.2, 0.17 mg/L, respectively) for 48 h | Increased immobility | [131] | |
| Daphnia magna | Bulk ZnO, nano ZnO and ZnSO4·7H2O at L(E)C50 (1.8, 1.9, 1.1 mg/L, respectively) for 48 h | Increased immobility | [131] |
4. Perspectives
Acknowledgments
Conflicts of Interest
References
- Bartosiewicz, M.; Penn, S.; Buckpitt, A. Applications of gene arrays in environmental toxicology: Fingerprints of gene regulation associated with cadmium chloride, benzo(a)pyrene, and trichloroethylene. Environ. Health Perspect. 2001, 109, 71–74. [Google Scholar] [CrossRef] [PubMed]
- Snape, J.R.; Maund, S.J.; Pickford, D.B.; Hutchinson, T.H. Ecotoxicogenomics: The challenge of integrating genomics into aquatic and terrestrial ecotoxicology. Aquat. Toxicol. 2004, 67, 143–154. [Google Scholar] [CrossRef] [PubMed]
- Miracle, A.L.; Ankley, G.T. Ecotoxicogenomics: Linkages between exposure and effects in assessing risks of aquatic contaminants to fish. Reprod. Toxicol. 2005, 19, 321–326. [Google Scholar] [CrossRef] [PubMed]
- Iguchi, T.; Watanabe, H.; Katsu, Y. Application of ecotoxicogenomics for studying endocrine disruption in vertebrates and invertebrates. Environ. Health. Perspect. 2006, 114, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, H.; Iguchi, T. Using ecotoxicogenomics to evaluate the impact of chemicals on aquatic organisms. Mar. Biol. 2006, 149, 107–115. [Google Scholar] [CrossRef]
- Altshuler, I.; Demiri, B.; Xu, S.; Constantin, A.; Yan, N.D.; Cristescu, M.E. An Integrated multi-disciplinary approach for studying multiple stressors in freshwater ecosystems: Daphnia as a model organism. Integr. Comp. Biol. 2011, 51, 623–633. [Google Scholar] [CrossRef] [PubMed]
- Nuwaysir, E.F.; Bittner, M.; Trent, J.; Barrett, J.C.; Afshari, C.A. Microarrays and toxicology: The advent of toxicogenomics. Mol. Carcinog. 1999, 24, 153–159. [Google Scholar] [CrossRef]
- Kurelec, B. The genotoxic disease syndrome. Mar. Environ. Res. 1993, 35, 341–348. [Google Scholar] [CrossRef]
- Oakshott, J.G. Selection at the Adh locus in Drosophila melanogaster imposed by environmental ethanol. Genet. Res. 1976, 26, 265–274. [Google Scholar] [CrossRef]
- Hawkins, A.J.S.; Rusia, J.; Bayne, B.L.; Day, A.J. The metabolic/physiological basis of genotype-dependent mortality during copper exposure in Mytilus edulis. Mar. Environ. Res. 1989, 28, 139–144. [Google Scholar] [CrossRef]
- Schat, H.; Ten Bookum, W.M. Genetic control of copper tolerance in Silene vulgaris. Heredity 1992, 68, 219–229. [Google Scholar] [CrossRef]
- Hebert, P.D.N. The population biology of Daphnia (Crustacea, Daphnidae). Biol. Rev. 1978, 53, 387–426. [Google Scholar] [CrossRef]
- Lampert, W. Daphnia: Model herbivore, predator and prey. Pol. J. Ecol. 2006, 54, 607–620. [Google Scholar]
- Cristescu, M.E.; Colbourne, J.K.; Radivojac, J.; Lynch, M. A microsatellite-based linkage map of the water flea Daphnia pulex: On the prospect of crustacean genomics. Genomics 2006, 88, 415–430. [Google Scholar] [CrossRef] [PubMed]
- Eads, B.D.; Andrews, J.; Colbourne, J.K. Ecological genomics in Daphnia: Stress responses and environmental sex determination. Heredity 2008, 100, 184–190. [Google Scholar] [CrossRef] [PubMed]
- Colbourne, J.K.; Hebert, P.D.N.; Taylor, D.J. Evolutionary origins of phenotypic diversity in daphnia. In Molecular Evolution and Adaptive Radiation; Givnish, T.J., Sytsma, K.J., Eds.; Cambridge University Press: Cambridge, UK, 1997; pp. 163–188. [Google Scholar]
- Schindler, D.W. Detecting ecosystem responses to anthropogenic stress. Can. J. Fish. Aquat. Sci. 1987, 44, 6–25. [Google Scholar] [CrossRef]
- Hebert, P.D.N.; Crease, T. Clonal diversity in populations of Daphnia pulex reproducing by obligate parthenogenesis. Heredity 1983, 51, 353–369. [Google Scholar] [CrossRef]
- Stich, H.B.; Lampert, W. Growth and reproduction of migrating and non-migrating Daphnia species under simulated food and temperature conditions of diurnal vertical migration. Oecologia 1984, 61, 192–196. [Google Scholar] [CrossRef]
- Dawidowicz, P.; Loose, C.J. Metabolic costs during predator-induced diel vertical migration of Daphnia. Limnol. Oceanogr. 1992, 37, 1589–1595. [Google Scholar] [CrossRef]
- Gerhardt, A.; de Bisthoven, B.L.; Soares, A.M.V. Evidence for the stepwise stress model: Gambusia holbrooki and Daphnia magna under acid mine drainage and acidified reference water stress. Environ. Sci. Technol. 2005, 39, 4150–4158. [Google Scholar] [CrossRef] [PubMed]
- Tollrian, R. Neckteeth formation in Daphnia pulex as an example of continuous phenotypic plasticity: Morphological effects of Chaoborus kairomone concentration and their quantification. J. Plankton Res. 1993, 15, 1309–1318. [Google Scholar] [CrossRef]
- Tautz, D. Not just another genome. BMC. Biol. 2011, 9, 8. [Google Scholar] [CrossRef] [PubMed]
- Fryer, G. Functional morphology and the adaptive radiation of the Daphnidae (Branchiopoda: Anomopoda). Philos. Trans. R. Soc. B. 1991, 331, 1–99. [Google Scholar] [CrossRef]
- Hebert, P.D.N. The Daphnia of North America: An Illustrated Fauna; University of Guelph: Guelph, ON, Canada, 1995. [Google Scholar]
- Wellborn, G.A.; Skelly, D.K.; Werner, E.E. Mechanisms creating community structure across a freshwater habitat gradient. Annu. Rev. Ecol. Syst. 1996, 27, 337–363. [Google Scholar] [CrossRef]
- Crease, T.J.; Lee, S.K.; Yu, S.L.; Spitze, K.; Lehman, N.; Lynch, M. Allozyme and mtDNA variation in populations of the Daphnia pulex complex from both sides of the Rocky Mountains. Heredity 1997, 79, 242–251. [Google Scholar] [CrossRef]
- Pfrender, M.E.; Spitze, K.; Lehman, N. Multi-locus genetic evidence for rapid ecologically based speciation in Daphnia. Mol. Ecol. 2000, 9, 1717–1735. [Google Scholar] [CrossRef] [PubMed]
- Boverhof, D.R.; Gollapudi, B.B. Applications of Toxicogenomics. In Safety Evaluation and Risk Assessment, 1st ed.; Wiley: Singapore, 2011. [Google Scholar]
- Larkin, P.; Villeneuve, D.L.; Knoebl, I.; Miracle, A.L.; Carter, B.J.; Liu, L.; Denslow, N.D.; Ankley, G.T. Development and validation of a 2000-gene microarray for the fathead minnow (Pimephales promelas). Environ. Toxicol. Chem. 2007, 26, 1497–1506. [Google Scholar] [CrossRef] [PubMed]
- Morgan, M.B.; Edge, S.E.; Snell, T.W. Profiling differential gene expression of corals along a transect of waters adjacent to the Bermuda municipal dump. Mar. Pollut. Bull. 2005, 51, 524–533. [Google Scholar] [CrossRef] [PubMed]
- Park, H.R.; Yang, H.; Kim, G.D.; Son, G.W.; Park, Y.S. Microarray analysis of gene expression in 3-methylcholanthrene-treated human endothelial cells. Mol. Cell. Toxicol. 2014, 10, 19–27. [Google Scholar] [CrossRef]
- Carney, S.A.; Chen, J.; Burns, C.G.; Xiong, K.M.; Peterson, R.E.; Heideman, W. Aryl hydrocarbon receptor activation produces heart-specific transcriptional and toxic responses in developing zebrafish. Mol. Pharmacol. 2006, 70, 549–561. [Google Scholar] [CrossRef] [PubMed]
- Santos, E.M.; Paull, G.C.; van Look, K.J.; Workman, V.L.; Holt, W.V.; van Aerle, R.; Kille, P.; Tyler, C.R. Gonadal transcriptome responses and physiological consequences of exposure to estrogen in breeding zebrafish (Danio rerio). Aquat. Toxicol. 2007, 83, 134–142. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.L.; Bencic, D.; Biales, A.; Lattier, D.; Kostich, M.; Villeneuve, D.; Ankley, G.T.; Lazorchak, J.; Toth, G. DNA microarray-based ecotoxicological biomarker discovery in a small fish model species. Environ. Toxicol. Chem. 2008, 27, 664–675. [Google Scholar] [CrossRef] [PubMed]
- Heckmann, L.H.; Connon, R.; Hutchinson, T.H.; Maund, S.J.; Sibly, R.M.; Callaghan, A. Expression of target and reference genes in Daphnia magna exposed to ibuprofen. BMC Genomics 2006, 7, 175. [Google Scholar] [CrossRef] [PubMed]
- Poynton, H.C.; Varshavsky, J.R.; Chang, B.; Cavigiolio, G.; Chan, S.; Holman, P.S.; Loguinov, A.V.; Bauer, D.J.; Komachi, K.; Theil, E.C.; et al. Daphnia magna ecotoxicogenomics provides mechanistic insights into metal toxicity. Environ. Sci. Technol. 2007, 41, 1044–1050. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, H.; Takahashi, E.; Nakamura, Y.; Oda, S.; Tatarazako, N.; Iguchi, T. Development of a Daphnia magna DNA microarray for evaluating the toxicity of environmental chemicals. Environ. Toxicol. Chem. 2007, 26, 669–676. [Google Scholar] [CrossRef] [PubMed]
- Connon, R.; Hooper, H.L.; Sibly, R.M.; Lim, F.L.; Heckmann, L.H.; Moore, D.J.; Watanabe, H.; Soetaert, A.; Cook, K.; Maund, S.J.; et al. Linking molecular and population stress responses in Daphnia magna exposed to cadmium. Environ. Sci. Technol. 2008, 42, 2181–2188. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Reyero, N.; Griffitt, R.J.; Liu, L. Construction of a robust microarray from a non-model species (largemouth bass) using pyrosequencing technology. J. Fish. Biol. 2008, 72, 2354–2376. [Google Scholar] [CrossRef] [PubMed]
- Soetaert, A.; Moens, L.N.; van der Ven, K.; van Leemput, K.; Naudts, B.; Blust, R.; de Coen, W.M. Molecular impact of propiconazole on Daphnia magna using a reproduction-related cDNA array. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2006, 142, 66–76. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.W.; Lee, S.M.; Yum, S.S.; Iguchi, T.; Seo, Y.R. Genomic expression responses toward bisphenol-A toxicity in Daphnia magna in terms of reproductive activity. Mol. Cell. Toxicol. 2013, 9, 149–158. [Google Scholar] [CrossRef]
- Dong, Y.; Zhai, L.; Zhang, L.; Jia, L.; Wang, X. Bisphenol A impairs mitochondrial function in spleens of mice via oxidative stress. Mol. Cell. Toxicol. 2013, 9, 401–406. [Google Scholar] [CrossRef]
- Poynton, H.C.; Lazorchak, J.M.; Impellitteri, C.A.; Blalock, B.J.; Rogers, K.; Allen, H.J.; Loguinov, A.; Heckman, J.L.; Govindasmawy, S. Toxicogenomic responses of nanotoxicity in Daphnia magna exposed to silver nitrate and coated silver nanoparticles. Environ. Sci. Technol. 2012, 46, 6288–6296. [Google Scholar] [CrossRef] [PubMed]
- Villeneuve, D.L.; Larkin, V.P.; Knoebl, I.; Miracle, A.L.; Kahl, M.D.; Jensen, K.M.; Makynen, E.A.; Durhan, E.J.; Carter, B.J.; Denslow, N.D.; et al. A graphical systems model to facilitate hypothesis-driven ecotoxicogenomics research on the teleost brain-pituitary-gonadal axis. Environ. Sci. Technol. 2007, 41, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Villeneuve, D.L.; Knoebl, I.; Larkin, P.; Miracle, A.L.; Carter, B.J.; Denslow, N.D.; Ankley, G.T. Altered gene expression in the brain and liver of female fathead minnows Pimephales promelas Rafinesque exposed to fadrozole. J. Fish. Biol. 2008, 72, 2281–2340. [Google Scholar] [CrossRef]
- Denslow, N.D.; Kocerha, J.; Sepúlveda, M.S.; Gross, T.; Holm, S.E. Gene expression fingerprints of largemouth bass (Micropterus salmoides) exposed to pulp and paper mill effluents. Mutat. Res. 2004, 552, 19–34. [Google Scholar] [CrossRef] [PubMed]
- Williams, T.D.; Diab, A.M.; George, S.G.; Godfrey, R.E.; Sabine, V.; Conesa, A.; Minchin, S.D.; Watts, P.C.; Chipman, J.K. Development of the GENIPOL European flounder (Platichthys flesus) microarray and determination of temporal transcriptional responses to cadmium at low dose. Environ. Sci. Technol. 2006, 40, 6479–6488. [Google Scholar] [CrossRef] [PubMed]
- Yeung, L.W.Y.; Guruge, K.S.; Yamanaka, N.; Miyazaki, S.; Lam, P.K.S. Differential expression of chicken hepatic genes responsive to PFOA and PFOS. Toxicology 2007, 237, 111–125. [Google Scholar] [CrossRef] [PubMed]
- Krasnov, A.; Koskinen, H.; Pehkonen, P.; Rexroad, C.E.; Afanasyev, S.; Mölsä, H. Gene expression in the brain and kidney of rainbow trout in response to handling stress. BMC Genomics 2005, 6, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krasnov, A.; Koskinen, H.; Pehkonen, P.; Rexroad, C.; Afanasyev, S.; Mölsä, H.; Oikari, A. Transcriptome responses to carbon tetrachloride and pyrene in the kidney and liver of juvenile rainbow trout (Oncorhynchus mykiss). Aquat. Toxicol. 2005, 74, 70–81. [Google Scholar] [CrossRef] [PubMed]
- Kassahn, K.S.; Crozier, R.H.; Ward, A.C.; Stone, G.; Caley, M.J. From transcriptome to biological function: Environmental stress in an ectothermic vertebrate, the coral reef fish Pomacentrus moluccensis. BMC Genomics 2007, 8, 358. [Google Scholar] [CrossRef] [PubMed]
- Lam, S.H.; Winata, C.L.; Tong, Y.; Korzh, S.; Lim, W.S.; Jan Spitsbergen, V.K.; Mathavan, S.; Miller, L.D.; Liu, E.T. Transcriptome kinetics of arsenic-induced adaptive response in zebrafish liver. Physiol. Genomics 2006, 27, 351–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Volz, D.C.; Hinton, D.E.; Law, J.M.; Kullman, S.W. Dynamic gene expression changes precede dioxin-induced liver pathogenesis in medaka fish. Toxicol. Sci. 2006, 89, 524–534. [Google Scholar] [CrossRef] [PubMed]
- Soetaert, A.; van der Ven, K.; Moens, L.N.; Vandenbrouck, T.; van Remortel, P.; de Coen, W.M. Daphnia magna and ecotoxicogenomics: Gene expression profiles of the anti-ecdysteroidal fungicide fenarimol using energy-, molting- and life stage-related cDNA libraries. Chemosphere 2007, 67, 60–71. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, H.; Tatarazako, N.; Oda, S.; Nishide, H.; Uchiyama, I.; Morita, M.; Iguchi, T. Analysis of expressed sequence tags of the water flea Daphnia magna. Genome 2005, 48, 606–609. [Google Scholar] [CrossRef] [PubMed]
- Margulies, M.; Egholm, M.; Altman, W.E.; Attiya, S.; Bader, J.S.; Bemben, L.A.; Berka, J.; Braverman, M.S.; Chen, Y.J.; Chen, Z.T.; et al. Genome sequencing in microfabricated high-density picolitre reactors. Nature 2005, 437, 376–380. [Google Scholar] [CrossRef] [PubMed]
- Hardy, S.; Legagneux, V.; Audic, Y.; Paillard, L. Reverse genetics in eukaryotes. Biol. Cell 2010, 102, 561–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martyniuk, C.J.; Trudeau, V.L. Fish endocrinology meets functional genomics: What exactly is the message? Gen. Comp. Endocrinol. 2009, 164, 132–134. [Google Scholar] [CrossRef] [PubMed]
- Kato, Y.; Kobayashi, K.; Watanabe, H.; Iguchi, T. Environmental sex determination in the branchipod crustacean Daphnia magna: Deep conservation of a double-sex gene in the sex-determining pathway. PLoS Genet. 2011, 7, e1001345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wetmore, B.A.; Merrick, B.A. Toxicoproteomics: Proteomics applied to toxicology and pathology. Toxicol. Pathol. 2004, 32, 619–642. [Google Scholar] [CrossRef] [PubMed]
- Shepard, J.L.; Olsson, B.; Tedengren, M.; Bradley, B.P. Protein expression signatures identified in Mytilus edulis exposed to PCBs, copper and salinity stress. Mar. Environ. Res. 2000, 50, 337–340. [Google Scholar] [CrossRef]
- Rodríguez-Ortega, M.J.; Grøsvik, B.E.; Rodríguez-Ariza, A.; Goksøyr, A.; López-Barea, J. Changes in protein expression profiles in bivalve molluscs (Chamaelea gallina) exposed to four model environmental pollutants. Proteomics 2003, 3, 1535–1543. [Google Scholar] [CrossRef] [PubMed]
- Shrader, E.A.; Henry, T.R.; Greeley, M.S., Jr.; Bradley, B.P. Proteomics in zebrafish exposed to endocrine disrupting chemicals. Ecotoxicology 2003, 12, 485–458. [Google Scholar] [CrossRef] [PubMed]
- Hogstrand, C.; Balesaria, S.; Glover, C.N. Application of genomics and proteomics for study of the integrated response to zinc exposure in a non-model fish species, the rainbow trout. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2002, 133, 523–535. [Google Scholar] [CrossRef]
- Bjørnstad, A.; Larsen, B.K.; Skadsheim, A.; Jones, M.B.; Andersen, O.K. The potential of ecotoxicoproteomics in environmental monitoring: Biomarker profiling in mussel plasma using Protein Chip array technology. J. Toxicol. Environ. Health A 2006, 69, 77–96. [Google Scholar] [CrossRef] [PubMed]
- Le, T.H.; Lim, E.S.; Hong, N.H.; Lee, S.K.; Shim, Y.S.; Hwang, J.R.; Kim, Y.H.; Min, J. Proteomic analysis in Daphnia magna exposed to As(III), As(V) and Cd heavy metals and their binary mixtures for screening potential biomarkers. Chemosphere 2013, 93, 2341–2348. [Google Scholar] [CrossRef] [PubMed]
- Rainville, L.C.; Carolan, D.; Varela, A.C.; Doyle, H.; Sheehan, D. Proteomic evaluation of citrate-coated silver nanoparticles toxicity in Daphnia magna. Analyst 2014, 7, 1678–1686. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.S.; Lee, M.Y.; David, A.E.; Park, Y.S. Nanoparticles for gene delivery: Therapeutic and toxic effects. Mol. Cell. Toxicol. 2014, 10, 1–8. [Google Scholar] [CrossRef]
- Ralston-Hooper, K.; Hopf, A.; Oh, C.; Zhang, X.; Adamec, J.; Sepúlveda, M.S. Development of GCxGC/TOF-MS metabolomics for use in ecotoxicological studies with invertebrates. Aquat. Toxicol. 2008, 2, 48–52. [Google Scholar] [CrossRef] [PubMed]
- Yoo, G.; Jeong, S.H.; Ryu, W.I.; Lee, H.; Kim, J.H.; Bae, H.C.; Son, S.W. Gene expression analysis reveals a functional role for the Ag-NPs-induced Egr-1 transcriptional factor in human keratinocytes. Mol. Cell. Toxicol. 2014, 10, 149–156. [Google Scholar] [CrossRef]
- Park, Y.H.; Bae, H.C.; Jang, Y.; Jeong, S.H.; Lee, H.N.; Ryu, W.I.; Yoo, M.G.; Kim, Y.R.; Kim, M.K.; Lee, J.K.; et al. Effect of the size and surface charge of silica nanoparticles on cutaneous toxicity. Mol. Cell. Toxicol. 2013, 9, 67–74. [Google Scholar] [CrossRef]
- Bundy, J.G.; Keun, H.C.; Sidhu, J.K.; Spurgeon, D.J.; Svendsen, C.; Kille, P.; Morgan, A.J. Metabolic profile biomarkers of metal contamination in a sentinel terrestrial species are applicable across multiple sites. Environ. Sci. Technol. 2007, 41, 4458–4464. [Google Scholar] [CrossRef] [PubMed]
- Taylor, N.S.; Weber, R.J.M.; Southam, A.D.; Payne, T.G.; Hrydziuszko, O.; Arvanitis, T.N.; Viant, M.R. A new approach to toxicity testing in Daphnia magna: Application of high throughput FT-ICR mass spectrometry metabolomics. Metabolomics 2009, 5, 44–58. [Google Scholar] [CrossRef]
- Taylor, N.S.; Weber, R.J.; White, T.A.; Viant, M.R. Discriminating between different acute chemical toxicities via changes in the daphnid metabolome. Toxicol. Sci. 2010, 118, 307–317. [Google Scholar] [CrossRef] [PubMed]
- Stohs, S.J.; Bagchi, D. Oxidative mechanisms in the toxicity of metal ions. Free Radic. Biol. Med. 1995, 18, 321–336. [Google Scholar] [CrossRef]
- Ray, D.E.; Fry, J.R. A reassessment of the neurotoxicity of pyrethroid insecticides. Pharmacol. Ther. 2006, 111, 174–193. [Google Scholar] [CrossRef] [PubMed]
- Drysdale, G.R.; Cohn, M. Mode of action of 2,4-dinitrophenol in uncoupling oxidative phosphorylation. J. Biol. Chem. 1958, 233, 1574–1577. [Google Scholar] [PubMed]
- Huggett, D.B.; Brooks, B.W.; Peterson, B.; Foran, C.M.; Schlenk, D. Toxicity of select beta adrenergic receptor-blocking pharmaceuticals (b-blockers) on aquatic organisms. Arch. Environ. Contam. Toxicol. 2002, 43, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Poynton, H.C.; Taylor, N.S.; Hicks, J.; Colson, K.; Chan, S.; Clark, C.; Scanlan, L.; Loguinov, A.V.; Vulpe, C.; Viant, M.R. Metabolomics of microliter hemolymph samples enables an improved understanding of the combined metabolic and transcriptional responses of Daphnia magna to cadmium. Environ. Sci. Technol. 2011, 45, 3710–3717. [Google Scholar] [CrossRef] [PubMed]
- Nagato, E.G.; D’eon, J.C.; Lankadurai, B.P.; Poirier, D.G.; Reiner, E.J.; Simpson, A.J.; Simpson, M.J. 1H NMR-based metabolomics investigation of Daphnia magna responses to sub-lethal exposure to arsenic, copper and lithium. Chemosphere 2013, 93, 331–337. [Google Scholar] [CrossRef] [PubMed]
- Ho, D.H.; Burggren, W.W. Epigenetics and transgenerational transfer: A physiological perspective. J. Exp. Biol. 2010, 213, 3–16. [Google Scholar] [CrossRef] [PubMed]
- Vandegehuchte, M.B.; Janssen, C.R. Epigenetics and its implications for ecotoxicology. Ecotoxicology 2011, 20, 607–624. [Google Scholar] [CrossRef] [PubMed]
- Jaenisch, R.; Bird, A. Epigenetic regulation of gene expression: How the genome integrates intrinsic and environmental signals. Nat. Genet. 2003, 33, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Santos, K.F.; Mazzola, T.N.; Carvalho, H.F. The prima donna of epigenetics: The regulation of gene expression by DNA methylation. Braz. J. Med. Biol. Res. 2005, 38, 1531–1541. [Google Scholar] [CrossRef] [PubMed]
- Fuks, F. DNA methylation and histone modifications: Teaming up to silence genes. Curr. Opin. Genet. Dev. 2005, 15, 490–495. [Google Scholar] [CrossRef] [PubMed]
- Lennartsson, A.; Ekwall, K. Histone modification patterns and epigenetic codes. Biochim. Biophys. Acta 2009, 1790, 863–868. [Google Scholar] [CrossRef] [PubMed]
- Lippman, Z.; Martienssen, R. The role of RNA interference in heterochromatic silencing. Nature 2004, 431, 364–370. [Google Scholar] [CrossRef] [PubMed]
- Carthew, R.W.; Sontheimer, E.J. Origins and Mechanisms of miRNAs and siRNAs. Cell 2009, 136, 642–655. [Google Scholar] [CrossRef] [PubMed]
- Reamon-Buettner, S.M.; Mutschler, V.; Borlak, J. The next innovation cycle in toxicogenomics: Environmental epigenetics. Mutat. Res. 2008, 659, 158–165. [Google Scholar] [CrossRef] [PubMed]
- Krieg, A.J.; Rankin, E.B.; Chan, D.; Razorenova, O.; Fernandez, S.; Giaccia, A.J. Regulation of the histone demethylase JMJD1A by hypoxia-inducible factor 1 alpha enhances hypoxic gene expression and tumor growth. Mol. Cell. Biol. 2010, 30, 344–353. [Google Scholar] [CrossRef] [PubMed]
- Vandegehuchte, M.B.; Vandenbrouck, T.; Coninck, D.D.; de Coen, W.M.; Janssen, C.R. Can metal stress induce transferable changes in gene transcription in Daphnia magna? Aquat. Toxicol. 2010, 97, 188–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aluru, N.; Karchner, S.I.; Hahn, M.E. Role of DNA methylation of AHR1 and AHR2 promoters in differential sensitivity to PCBs in Atlantic Killifish, Fundulus heteroclitus. Aquat. Toxicol. 2011, 101, 288–294. [Google Scholar] [CrossRef] [PubMed]
- Vandegehuchte, M.B.; Kyndt, T.; Vanholme, B.; Haegeman, A.; Gheysen, G.; Janssen, C.R. Occurrence of DNA methylation in Daphnia magna and influence of multigeneration Cd exposure. Environ. Int. 2009, 35, 700–706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Youngson, N.A.; Whitelaw, E. Transgenerational epigenetic effects. Annu. Rev. Genomics Hum. Genet. 2008, 9, 233–257. [Google Scholar] [CrossRef] [PubMed]
- Vandegehuchte, M.B.; Lemi’ere, F.; Vanhaecke, L.; Vanden Berghe, W.; Janssen, C.R. Direct and transgenerational impact on Daphnia magna of chemicals with a known effect on DNA methylation. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2010, 151, 278–285. [Google Scholar] [CrossRef] [PubMed]
- Vandegehuchte, M.B.; Vandenbrouck, T.; de Coninck, D.; de Coen, W.M.; Janssen, C.R. Gene transcription and higher-level effects of multigenerational Zn exposure in Daphnia magna. Chemosphere 2010, 80, 1014–1020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eads, B.D.; Colbourne, J.K.; Bohuski, E.; Andrews, J. Profiling sex-biased gene expression during parthenogenetic reproduction in Daphnia pulex. BMC Genomics 2007, 8, 464. [Google Scholar] [CrossRef] [PubMed]
- Agrawal1, A.A.; Laforsch, C.; Tollrian, R. Transgenerational induction of defences in animals and plants. Nature 1999, 401, 60–63. [Google Scholar] [CrossRef]
- Miyakawa, H; Imai, M.; Sugimoto, N.; Ishikawa, Y.; Ishikawa, A.; Ishigaki, H.; Okada, Y.; Miyazaki, S.; Koshikawa, S.; Cornette, R.; et al. Gene up-regulation in response to predator kairomones in the water flea, Daphnia pulex. BMC Dev. Biol. 2010, 10, 45. [Google Scholar]
- Vandegehuchte, M.B.; Lemière, F.; Janssen, C.R. Quantitative DNA-methylation in Daphnia magna and effects of multigeneration Zn exposure. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2009, 150, 343–348. [Google Scholar] [CrossRef] [PubMed]
- Soetaert, A.; Vandenbrouck, T.; van der Ven, K.; Maras, M.; van Remortel, P.; Blust, R.; de Coen, W.M. Molecular responses during cadmium-induced stress in Daphnia magna: Integration of differential gene expression with higher-level effects. Aquat. Toxicol. 2007, 83, 212–222. [Google Scholar] [CrossRef] [PubMed]
- Klaper, R.; Crago, J.; Barr, J.; Arndt, D.; Setyowati, K.; Chen, J. Toxicity biomarker expression in daphnids exposed to manufactured nanoparticles: Changes in toxicity with functionalization. Environ. Pollut. 2009, 157, 1152–1156. [Google Scholar] [CrossRef] [PubMed]
- Heckmann, L.H.; Sibly, R.M.; Connon, R.; Hooper, H.L.; Hutchinson, T.H.; Maund, S.J.; Hill, C.J.; Bouetard, A.; Callaghan, A. Systems biology meets stress ecology: Linking molecular and organismal stress responses in Daphnia magna. Genome. Biol. 2008, 9, R40. [Google Scholar] [CrossRef] [PubMed]
- Merzendorfer, H.; Zimoch, L. Chitin metabolism in insects: Structure, function and regulation of chitin synthases and chitinases. J. Exp. Biol. 2003, 206, 4393–4412. [Google Scholar] [CrossRef] [PubMed]
- Hennig, H.F. Metal-binding proteins as metal pollution indicators. Environ. Health Perspect. 1986, 65, 175–187. [Google Scholar] [PubMed]
- Gaetke, L.M.; Chow, C.K. Copper toxicity, oxidative stress, and antioxidant nutrients. Toxicology 2003, 189, 147–163. [Google Scholar] [CrossRef]
- Wang, Y.J.; Lee, C.C.; Chang, W.C.; Liou, H.B.; Ho, Y.S. Oxidative stress and liver toxicity in rats and human hepatoma cell line induced by pentachlorophenol and its major metabolite tetrachlorohydroquinone. Toxicol. Lett. 2001, 122, 157–169. [Google Scholar] [CrossRef]
- Xie, F.; Koziar, S.A.; Lampi, M.A.; Dixon, D.G.; Norwood, W.P.; Borgmann, U.; Huang, X.-D.; Greenberg, B.M. Assessment of the toxicity of mixtures of copper, 9,10-phenanthrenequinone, and phenanthrene to Daphnia magna: Evidence for a reactive oxygen mechanism. Environ. Toxicol. Chem. 2006, 25, 613–622. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.K.; Song, J.W.; Park, J.D.; Choi, B.S. Copper induces the accumulation of amyloid-beta in the brain. Mol. Cell. Toxicol. 2013, 9, 57–66. [Google Scholar] [CrossRef]
- Park, H.G.; Yeo, M.K. Effects of TiO2 nanoparticles and nanotubes on zebrafish caudal fin regeneration. Mol. Cell. Toxicol. 2013, 9, 375–383. [Google Scholar] [CrossRef]
- Park, H.G.; Kim, J.I.; Kang, M.; Yeo, M.K. The effect of metal-doped TiO2 nanoparticles on zebrafish embryogenesis. Mol. Cell. Toxicol. 2014, 10, 293–301. [Google Scholar] [CrossRef]
- Ote, M.; Mita, K.; Kawasaki, H.; Daimon, T.; Kobayashi, M.; Shimada, T. Identification of molting fluid carboxypeptidase A (MF-CPA) in Bombyx mori. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2005, 141, 314–322. [Google Scholar] [CrossRef] [PubMed]
- Kato, Y.; Tokishita, S.; Ohta, T.; Yamagata, H. A vitellogenin chain containing a superoxide dismutase-like domain is the major component of yolk proteins in cladoceran crustacean Daphnia magna. Gene 2004, 334, 157–165. [Google Scholar] [CrossRef] [PubMed]
- OECD/OCDE 202, OECD Guideline for Testing of Chemicals; Daphnia sp. Acute Immobilisation Test. Available online: http://www.oecd-ilibrary.org/environment/test-no-202-daphnia-sp-acute-immobilisation-test_9789264069947-en (accessed on 28 May 2015).
- OECD/OCDE 211, OECD Guideline for Testing of Chemicals; Daphnia magna Reproduction Test. Available online: http://www.oecd-ilibrary.org/environment/test-no-211-daphnia-magna-reproduction-test_9789264185203-en (accessed on 28 May 2015).
- McLachlan, J.A. Environmental signaling: What embryos and evolution teach us about endocrine disrupting chemicals. Endocr. Rev. 2001, 22, 319–341. [Google Scholar] [CrossRef] [PubMed]
- Iguchi, T.; Sumi, M.; Tanabe, S. Endocrine disruptor issues in Japan. Congen. Anorm. 2002, 42, 106–119. [Google Scholar] [CrossRef]
- Comber, M.H.I.; Williams, T.D.; Stewart, K.M. The effects of nonylphenol on Daphnia magna. Water Res. 1993, 27, 273–276. [Google Scholar] [CrossRef]
- Tatarazako, N.; Takao, Y.; Kishi, K.; Onikura, N.; Arizono, K.; Iguchi, T. Styrene dimers and trimers affect reproduction of daphnid (Ceriodaphnia dubia). Chemosphere 2002, 48, 597–601. [Google Scholar] [CrossRef]
- Tatarazako, N.; Oda, S.; Watanabe, H.; Morita, M.; Iguchi, T. Juvenile hormone agonists affect the occurrence of male Daphnia. Chemosphere 2003, 53, 827–833. [Google Scholar] [CrossRef]
- Schultz, T.W.; Freeman, S.R.; Dumont, J.N. Uptake, depuration, and distribution of selenium in Daphnia and its effects on survival and ultrastructure. Arch. Environ. Contam. Toxicol. 1980, 9, 23–40. [Google Scholar] [CrossRef] [PubMed]
- Cooper, N.L.; Bidwell, J.R.; Kumar, A. Toxicity of copper, lead, and zinc mixtures to Ceriodaphniadubia and Daphnia carinata. Ecotoxicol. Environ. Saf. 2009, 72, 1523–1528. [Google Scholar] [CrossRef] [PubMed]
- Jurado, E.; Fernández-Serrano, M.; Núñez Olea, J.; Lechuga, M.; Jiménez, J.L.; Ríos, F. Acute toxicity of alkylpolyglucosides to Vibrio fischeri, Daphnia magna and microalgae: A comparative study. Bull. Environ. Contam. Toxicol. 2012, 88, 290–295. [Google Scholar] [CrossRef] [PubMed]
- Collard, H.J.; Ji, K.; Lee, S.; Liu, X.; Kang, S.; Kho, Y.; Ahn, B.; Ryu, J.; Lee, J.; Choi, K. Toxicity and endocrine disruption in zebrafish (Danio rerio) and two freshwater invertebrates (Daphnia magna and Moina macrocopa) after chronic exposure to mefenamic acid. Ecotoxicol. Environ. Saf. 2013, 94, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.M.; Wang, W.X. Importance of surface coatings and soluble silver in silver nanoparticles toxicity to Daphnia magna. Nanotoxicology 2012, 6, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Newton, K.M.; Puppala, H.L.; Kitchens, C.L.; Colvin, V.L.; Klaine, S.J. Silver nanoparticle toxicity to Daphnia magna is a function of dissolved silver concentration. Environ. Toxicol. Chem. 2013, 32, 2356–2364. [Google Scholar] [CrossRef] [PubMed]
- Blinova, I.; Niskanen, J.; Kajankari, P.; Kanarbik, L.; Käkinen, A.; Tenhu, H.; Penttinen, O.P.; Kahru, A. Toxicity of two types of silver nanoparticles to aquatic crustaceans Daphnia magna and Thamnocephalus platyurus. Environ. Sci. Pollut. Res. Int. 2013, 20, 3456–3463. [Google Scholar] [CrossRef] [PubMed]
- Jo, H.J.; Choi, J.W.; Lee, S.H.; Hong, S.W. Acute toxicity of Ag and CuO nanoparticle suspensions against Daphnia magna: The importance of their dissolved fraction varying with preparation methods. J. Hazard. Mater 2012, 227–228, 301–308. [Google Scholar] [CrossRef] [PubMed]
- Fouqueray, M.; Dufils, B.; Vollat, B.; Chaurand, P.; Botta, C.; Abacci, K.; Labille, J.; Rose, J.; Garric, J. Effects of aged TiO2 nanomaterial from sunscreen on Daphnia magna exposed by dietary route. Environ. Pollut. 2012, 163, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Heinlaan, M.; Ivask, A.; Blinova, I.; Dubourguier, H.C.; Kahru, A. Toxicity of nanosized and bulk ZnO, CuO and TiO2 to bacteria Vibrio fischeri and crustaceans Daphnia magna and Thamnocephalus platyurus. Chemosphere 2008, 71, 1308–1316. [Google Scholar] [CrossRef] [PubMed]
- Lubbock, J. An account of the two methods of reproduction in Daphnia, and of the structure of the ephippium. Philos. Trans. R. Soc. Lond. 1857, 147, 79–100. [Google Scholar] [CrossRef]
- Zaffagnini, F.; Peters, R.H.; de Bernardi, R. Reproduction in Daphnia. Mem. Ist. Ital. Idrobiol. 1987, 45, 245–284. [Google Scholar]
- Kleiven, O.T.; Larsson, P.; Oikos, A.H. Sexual reproduction in Daphnia magna requires three stimuli. OIKOS 1992, 65, 197–206. [Google Scholar] [CrossRef]
- Thompson, J.D. Phenotypic plasticity as a component of evolutionary change. Trends Evol. Ecol. 1991, 6, 246–249. [Google Scholar] [CrossRef]
- Depledge, M.H. Genotypic toxicity: Implications for individuals and populations. Environ. Health Perspect. 1994, 102, 101–104. [Google Scholar] [CrossRef] [PubMed]
- Shugart, L.R.; Theodorakis, C. Genetic ecotoxicology: The genotypic diversity approach. Comp. Biochem. Physiol. 1996, 113, 273–276. [Google Scholar] [CrossRef]
- Moens, L.N.; van der Ven, K.; van Remortel, P.; del-Favero, J.; de Coen, W.M. Expression profiling of endocrine-disrupting compounds using a customized Cyprinus carpio cDNA microarray. Toxicol. Sci. 2006, 93, 298–310. [Google Scholar] [CrossRef] [PubMed]
- Moens, L.N.; Smolders, R.; van der Ven, K.; van Remortel, P.; del-Favero, J.; de Coen, W.M. Effluent impact assessment using microarray-based analysis in common carp: A systems toxicology approach. Chemosphere 2007, 67, 2293–2304. [Google Scholar] [CrossRef] [PubMed]
- Koedrith, P.; Boonprasert, R.; Kwon, J.Y.; Kim, I.S.; Seo, Y.R. Recent toxicological investigations of metal or metal oxide nanoparticles in mammalian models in vitro and in vivo: DNA damaging potential, and relevant physicochemical characteristics. Mol. Cell. Toxicol. 2014, 10, 107–126. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, H.J.; Koedrith, P.; Seo, Y.R. Ecotoxicogenomic Approaches for Understanding Molecular Mechanisms of Environmental Chemical Toxicity Using Aquatic Invertebrate, Daphnia Model Organism. Int. J. Mol. Sci. 2015, 16, 12261-12287. https://doi.org/10.3390/ijms160612261
Kim HJ, Koedrith P, Seo YR. Ecotoxicogenomic Approaches for Understanding Molecular Mechanisms of Environmental Chemical Toxicity Using Aquatic Invertebrate, Daphnia Model Organism. International Journal of Molecular Sciences. 2015; 16(6):12261-12287. https://doi.org/10.3390/ijms160612261
Chicago/Turabian StyleKim, Hyo Jeong, Preeyaporn Koedrith, and Young Rok Seo. 2015. "Ecotoxicogenomic Approaches for Understanding Molecular Mechanisms of Environmental Chemical Toxicity Using Aquatic Invertebrate, Daphnia Model Organism" International Journal of Molecular Sciences 16, no. 6: 12261-12287. https://doi.org/10.3390/ijms160612261