
Chemokine-Derived Peptides: Novel Antimicrobial and Antineoplasic Agents

Abstract
:1. Introduction
2. Chemokines and Microbial Infections
2.1. Antibacterial Activities
2.2. Anti-Viral and Anti-Parasite Activities
3. Chemokine-Derived Peptides as Antimicrobials

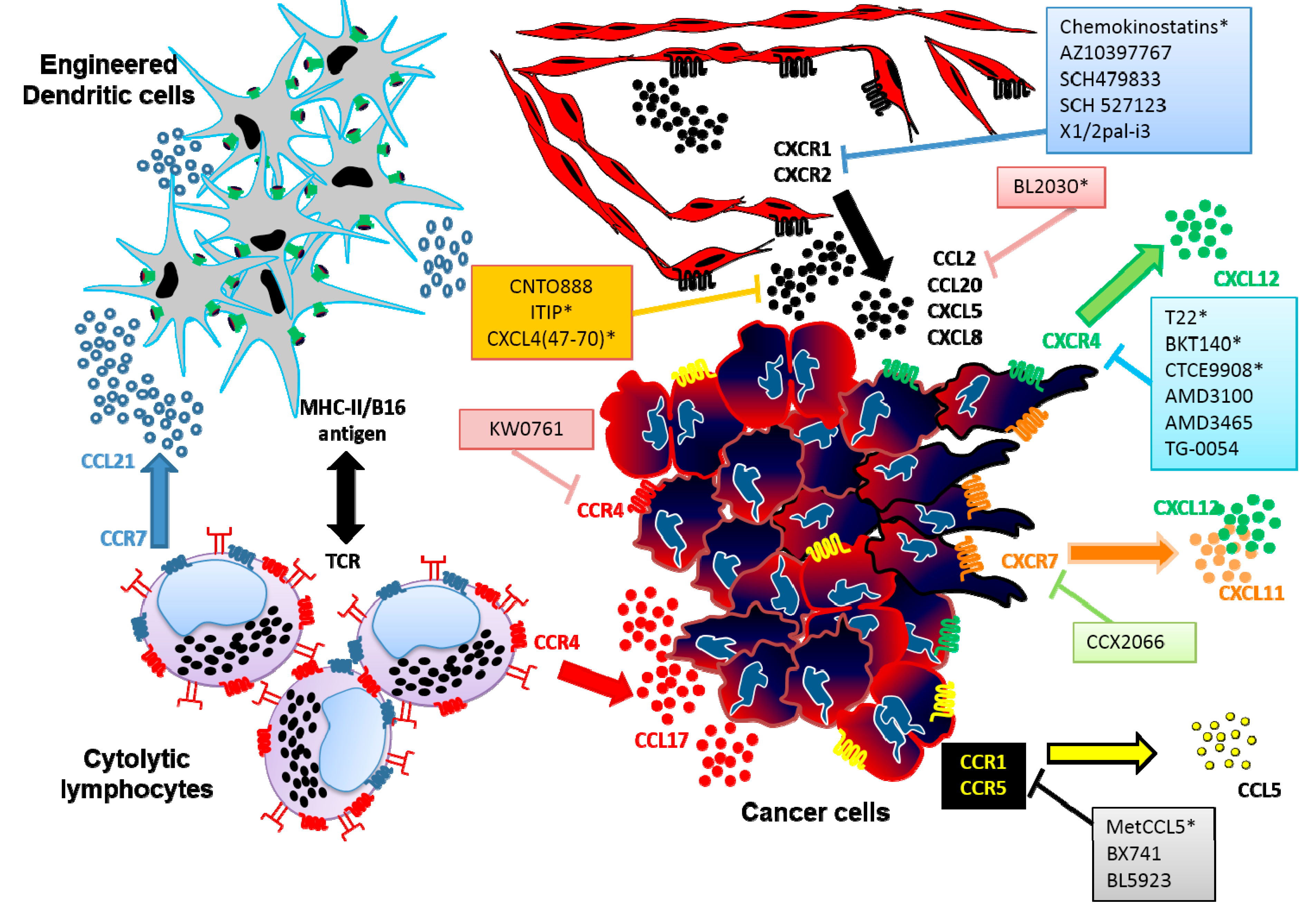{kind=link}
| CDP | Chemokine | Region | Biological Target | References |
|---|---|---|---|---|
| vMIP-II r1-21 | vMIP-II | N-terminal | Inhibition of HIV-1 entry | [45] |
| MIP-3α51–70 | CCL20/MIP-3α | C-terminal | E. coli (ML35) | [23,51] |
| MIP-3α59–70 | CCL20/MIP-3α | C-terminal | E. coli (ML35) | [52] |
| TC-1 | CXCL7/NAP-2 | C-terminal | E. coli (ML35) B. subtilis (ATCC6633) S. aureus (42D) C. neoformans (CI) C. glabrata (CI) | [51] |
| IL-881–99 | CXCL8/IL-8 | C-terminal | E. coli (MG1655) S. enterica (MS10) K. pneumoniae (CCUG 49243) H. pylori (SS1) S. pyogenes (CCUG 49246) | [58] |
| CCL1357–75 CDAP-4 | CCL13/MCP-4 | C-terminal | E. coli (ATCC25922) S. typhimurium (ATCC 14028) S. typhi (ATCC 9993) K. pneumonia (PIMM-UNAM28) P. aeruginosa (ATCC15692) P. aeruginosa (CI) | [59] |
| RP-1 | CXCL4/PF4 | C-terminal | L. major L. infantum chagasi (MHOM/BR/00/1669) L. braziliensis (CI) | [60] |
| PMX207, PMX1207 | CXCL4/PF4 | C-terminal | P. falciparum (3D7) E. coli (ATCC 25922) S. aureus (ATCC 27660) E. faecalis (ATCC 29212) P. aeruginosa (ATCC 10145) K. pneumoniae (ATCC 13883) | [61] |
4. Chemokines, Chemokine-Derived Peptides and Cancer
4.1. Paradoxical Roles of Chemokines in Cancer
4.2. Inflammation and Cancer
4.3. Angiogenesis Regulation
4.4. Tumor Progression
4.5. Chemokine-Derived Peptides with Anti-Tumour Activity

| Chemokine | Receptor Inhibitor | Receptor Antagonist | CDP | Other Strategies | Biological Effects and Clinical Trials |
|---|---|---|---|---|---|
| CCL2 CCL5 CCL20 | Carlumab (CNTO888) [116,117,118,119] Anti-CCL20 [115] | BL5923 [135] BX-471 [134] | Met-CCL5 [136] | BL2030 (a soluble CCR2 receptor fragment) [120] | Regulate leukocyte infiltration Inhibition of TAM formation Improve TLS formation CNTO888 (Phase II) BL5923 and BX471 (Preclinical) BL2030 (Preclinical data) |
| CCL4 CCL17 CCL22 | Mogamulizumab (KW-0761) [137,138] | - | - | Cytotoxic lymphocytes with CCR4 chimeric receptor | Modulation of tumor immune response Inhibition of migration of CCR4+ regulatory T cells Improvement of TLS formation Regulation of leukocyte infiltration KW-0761 (Phase II) |
| CCL21 CCL19 CXCL13 | - | - | - | Enhanced production | Improvement of TLS formation [148] |
| CXCL1 CXCL2 CXCL3 CXCL5 CXCL6 CXCL7 CXCL8 | Anti-CXCL5, Anti-CXCL8 [114] | SCH-479833 Navirixin (SCH-527123) AZ10397767 [150,151,153] | Chemokinostatins Pepducin X1/2pal-i3 [88,139,140,141,142,143,152] | - | Inhibition of tumor growth and survival Anti-angiogenesis SCH527123 (Phase I and Phase II) SCH-479833, AZ10397767, X1/2pal-i3 and chemokinostatins (Preclinical) |
| CXCL4L1 | - | - | CXCL4L1/PF4var47–70 [149] | - | Anti-angiogenesis Preclinical studies |
| CXCL4 CXCL9 CXCL10 CXCL11 CXCL13 CXCL14 | AMG487 [132] | - | CXCL4/PF4(47–70) [140] Chimeric ITIP [154] | - | Tumor growth inhibition Anti-angiogenesis AMG487, ITIP and PF447–70 (Preclinical) |
| CXCL12 CCL19 CCL21 | AMD3465 CCX2066 [131,132] | AMD3100 (Plerixafor) MSX-122 (Metastatix) POL-6326 TG-0054 (Burixafor) [123,124,125,126] | T22, BKT140 (4F-benzoyl-TN14003), CTCE-9908 [127,128,129,130] | Aptamer NOX-A12 (Spiegelmer) [129] Carrier for gene delivery CXCR4: Nter-KPVSLSYR [133] | Inhibition of tumor growth Decrease metastasis Anti-angiogenesis AMD3100 (FDA approved) MSX-122 (Phase I) POL6326 (Phase II) TG-0054 (Phase II) NOX-A12 (Phase II) BKT140 (Phase I) CTCE-9908 (FDA approved) AMD3465 (Preclinical) CCX2066 (Preclinical) |
| CXCL16 | - | - | - | Enhanced production | NK and T lymphocytes recruitment, Antitumoral activities |
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Rossi, D.; Zlotnik, A. The biology of chemokines and their receptors. Annu. Rev. Immunol. 2000, 18, 217–242. [Google Scholar] [CrossRef] [PubMed]
- Griffith, J.W.; Sokol, C.L.; Luster, A.D. Chemokines and chemokine receptors: Positioning cells for host defense and immunity. Annu. Rev. Immunol. 2014, 32, 659–702. [Google Scholar] [CrossRef] [PubMed]
- Bachelerie, F.; Ben-Baruch, A.; Burkhardt, A.M.; Combadiere, C.; Farber, J.M.; Graham, G.J.; Horuk, R.; Sparre-Ulrich, A.H.; Locati, M.; Luster, A.D.; et al. International Union of Basic and Clinical Pharmacology. [corrected]. LXXXIX. Update on the extended family of chemokine receptors and introducing a new nomenclature for atypical chemokine receptors. Pharmacol. Rev. 2014, 66, 1–79. [Google Scholar] [CrossRef] [PubMed]
- Zweemer, A.J.M.; Toraskar, J.; Heitman, L.H.; IJzerman, A.P. Bias in chemokine receptor signalling. Trends Immunol. 2014, 35, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Rollins, B.J. Chemokines. Blood 1997, 90, 909–928. [Google Scholar] [PubMed]
- Biragyn, A.; Surenhu, M.; Yang, D.; Ruffini, P.A.; Haines, B.A.; Klyushnenkova, E.; Oppenheim, J.J.; Kwak, L.W. Mediators of innate immunity that target immature, but not mature, dendritic cells induce antitumor immunity when genetically fused with nonimmunogenic tumor antigens. J. Immunol. 2001, 167, 6644–6653. [Google Scholar] [CrossRef] [PubMed]
- Balkwill, F. Cancer and the chemokine network. Nat. Rev. Cancer 2004, 4, 540–550. [Google Scholar] [CrossRef] [PubMed]
- Orimo, A.; Gupta, P.B.; Sgroi, D.C.; Arenzana-Seisdedos, F.; Delaunay, T.; Naeem, R.; Carey, V.J.; Richardson, A.L.; Weinberg, R.A. Stromal fibroblasts present in invasive human breast carcinomas promote tumor growth and angiogenesis through elevated SDF-1/CXCL12 secretion. Cell 2005, 121, 335–348. [Google Scholar] [CrossRef] [PubMed]
- Miao, Z.; Luker, K.E.; Summers, B.C.; Berahovich, R.; Bhojani, M.S.; Rehemtulla, A.; Kleer, C.G.; Essner, J.J.; Nasevicius, A.; Luker, G.D.; et al. CXCR7 (RDC1) promotes breast and lung tumor growth in vivo and is expressed on tumor-associated vasculature. Proc. Natl. Acad. Sci. USA 2007, 104, 15735–15740. [Google Scholar] [CrossRef] [PubMed]
- Strieter, R.M.; Polverini, P.J.; Kunkel, S.L.; Arenberg, D.A.; Burdick, M.D.; Kasper, J.; Dzuiba, J.; van Damme, J.; Walz, A.; Marriott, D.; et al. The functional role of the ELR motif in CXC chemokine-mediated angiogenesis. J. Biol. Chem. 1995, 270, 27348–27357. [Google Scholar] [CrossRef] [PubMed]
- Strieter, R.M.; Burdick, M.D.; Gomperts, B.N.; Belperio, J.A.; Keane, M.P. CXC chemokines in angiogenesis. Cytokine Growth Factor Rev. 2005, 16, 593–609. [Google Scholar] [CrossRef] [PubMed]
- Schall, T.J.; Proudfoot, A.E. Overcoming hurdles in developing successful drugs targeting chemokine receptors. Nat. Rev. Immunol. 2011, 11, 355–363. [Google Scholar] [CrossRef] [PubMed]
- Territo, M.C.; Ganz, T.; Selsted, M.E.; Lehrer, R. Monocyte-chemotactic activity of defensins from human neutrophils. J. Clin. Investig. 1989, 84, 2017–2020. [Google Scholar] [CrossRef] [PubMed]
- Pereira, H.A.; Shafer, W.M.; Pohl, J.; Martin, L.E.; Spitznagel, J.K. CAP37, a human neutrophil-derived chemotactic factor with monocyte specific activity. J. Clin. Investig. 1990, 85, 1468–1476. [Google Scholar] [CrossRef] [PubMed]
- Chertov, O.; Michiel, D.F.; Xu, L.; Wang, J.M.; Tani, K.; Murphy, W.J.; Longo, D.L.; Taub, D.D.; Oppenheim, J.J. Identification of defensin-1, defensin-2, and CAP37/azurocidin as T-cell chemoattractant proteins released from interleukin-8-stimulated neutrophils. J. Biol. Chem. 1996, 271, 2935–2940. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Chertov, O.; Bykovskaia, S.N.; Chen, Q.; Buffo, M.J.; Shogan, J.; Anderson, M.; Schroder, J.M.; Wang, J.M.; Howard, O.M.; et al. Beta-defensins: Linking innate and adaptive immunity through dendritic and T cell CCR6. Science 1999, 286, 525–528. [Google Scholar] [CrossRef] [PubMed]
- Perez-Canadillas, J.M.; Zaballos, A.; Gutierrez, J.; Varona, R.; Roncal, F.; Albar, J.P.; Marquez, G.; Bruix, M. NMR solution structure of murine CCL20/MIP-3alpha, a chemokine that specifically chemoattracts immature dendritic cells and lymphocytes through its highly specific interaction with the beta-chemokine receptor CCR6. J. Biol. Chem. 2001, 276, 28372–28379. [Google Scholar] [CrossRef] [PubMed]
- Hoover, D.M.; Boulegue, C.; Yang, D.; Oppenheim, J.J.; Tucker, K.; Lu, W.; Lubkowski, J. The structure of human macrophage inflammatory protein-3alpha/CCL20. Linking antimicrobial and CC chemokine receptor-6-binding activities with human beta-defensins. J. Biol. Chem. 2002, 277, 37647–37654. [Google Scholar] [CrossRef] [PubMed]
- Krijgsveld, J.; Zaat, S.A.; Meeldijk, J.; van Veelen, P.A.; Fang, G.; Poolman, B.; Brandt, E.; Ehlert, J.E.; Kuijpers, A.J.; Engbers, G.H.; et al. Thrombocidins, microbicidal proteins from human blood platelets, are C-terminal deletion products of CXC chemokines. J. Biol. Chem. 2000, 275, 20374–20381. [Google Scholar] [CrossRef] [PubMed]
- Cole, A.M.; Ganz, T.; Liese, A.M.; Burdick, M.D.; Liu, L.; Strieter, R.M. Cutting edge: IFN-inducible ELR− CXC chemokines display defensin-like antimicrobial activity. J. Immunol 2001, 167, 623–627. [Google Scholar] [CrossRef] [PubMed]
- Hieshima, K.; Ohtani, H.; Shibano, M.; Izawa, D.; Nakayama, T.; Kawasaki, Y.; Shiba, F.; Shiota, M.; Katou, F.; Saito, T.; et al. CCL28 has dual roles in mucosal immunity as a chemokine with broad-spectrum antimicrobial activity. J. Immunol. 2003, 170, 1452–1461. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Wilson, E. The antimicrobial activity of CCL28 is dependent on C-terminal positively-charged amino acids. Eur. J. Immunol. 2010, 40, 186–196. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Chen, Q.; Hoover, D.M.; Staley, P.; Tucker, K.D.; Lubkowski, J.; Oppenheim, J.J. Many chemokines including CCL20/MIP-3alpha display antimicrobial activity. J. Leukoc. Biol. 2003, 74, 448–455. [Google Scholar] [CrossRef] [PubMed]
- Zasloff, M. Antimicrobial peptides of multicellular organisms. Nature 2002, 415, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Lai, Y.; Gallo, R.L. AMPed up immunity: How antimicrobial peptides have multiple roles in immune defense. Trends Immunol. 2009, 30, 131–141. [Google Scholar] [CrossRef] [PubMed]
- Yung, S.C.; Murphy, P.M. Antimicrobial chemokines. Front. Immunol. 2012, 3, 276. [Google Scholar] [CrossRef] [PubMed]
- Wolf, M.; Moser, B. Antimicrobial activities of chemokines: Not just a side-effect? Front. Immunol. 2012, 3, 213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collin, M.; Linge, H.M.; Bjartell, A.; Giwercman, A.; Malm, J.; Egesten, A. Constitutive expression of the antibacterial CXC chemokine GCP-2/CXCL6 by epithelial cells of the male reproductive tract. J. Reprod. Immunol. 2008, 79, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Linge, H.M.; Collin, M.; Nordenfelt, P.; Morgelin, M.; Malmsten, M.; Egesten, A. The human CXC chemokine granulocyte chemotactic protein 2 (GCP-2)/CXCL6 possesses membrane-disrupting properties and is antibacterial. Antimicrob. Agents Chemother. 2008, 52, 2599–2607. [Google Scholar] [CrossRef] [PubMed]
- Maerki, C.; Meuter, S.; Liebi, M.; Muhlemann, K.; Frederick, M.J.; Yawalkar, N.; Moser, B.; Wolf, M. Potent and broad-spectrum antimicrobial activity of CXCL14 suggests an immediate role in skin infections. J. Immunol. 2009, 182, 507–514. [Google Scholar] [CrossRef] [PubMed]
- Hevezi, P.; Moyer, B.D.; Lu, M.; Gao, N.; White, E.; Echeverri, F.; Kalabat, D.; Soto, H.; Laita, B.; Li, C.; et al. Genome-wide analysis of gene expression in primate taste buds reveals links to diverse processes. PLoS ONE 2009, 4, e6395. [Google Scholar] [CrossRef] [PubMed]
- Meuter, S.; Moser, B. Constitutive expression of CXCL14 in healthy human and murine epithelial tissues. Cytokine 2008, 44, 248–255. [Google Scholar] [CrossRef] [PubMed]
- Frick, I.M.; Nordin, S.L.; Baumgarten, M.; Morgelin, M.; Sorensen, O.E.; Olin, A.I.; Egesten, A. Constitutive and inflammation-dependent antimicrobial peptides produced by epithelium are differentially processed and inactivated by the commensal Finegoldia magna and the pathogen Streptococcus pyogenes. J. Immunol. 2011, 187, 4300–4309. [Google Scholar] [CrossRef] [PubMed]
- Cederlund, A.; Gudmundsson, G.H.; Agerberth, B. Antimicrobial peptides important in innate immunity. FEBS J. 2011, 278, 3942–3951. [Google Scholar] [CrossRef] [PubMed]
- Burkhardt, A.M.; Tai, K.P.; Flores-Guiterrez, J.P.; Vilches-Cisneros, N.; Kamdar, K.; Barbosa-Quintana, O.; Valle-Rios, R.; Hevezi, P.A.; Zuniga, J.; Selman, M.; et al. CXCL17 is a mucosal chemokine elevated in idiopathic pulmonary fibrosis that exhibits broad antimicrobial activity. J. Immunol. 2012, 188, 6399–6406. [Google Scholar] [CrossRef] [PubMed]
- Crawford, M.A.; Zhu, Y.; Green, C.S.; Burdick, M.D.; Sanz, P.; Alem, F.; O’Brien, A.D.; Mehrad, B.; Strieter, R.M.; Hughes, M.A. Antimicrobial effects of interferon-inducible CXC chemokines against Bacillus anthracis spores and bacilli. Infect. Immun. 2009, 77, 1664–1678. [Google Scholar] [CrossRef] [PubMed]
- Crawford, M.A.; Burdick, M.D.; Glomski, I.J.; Boyer, A.E.; Barr, J.R.; Mehrad, B.; Strieter, R.M.; Hughes, M.A. Interferon-inducible CXC chemokines directly contribute to host defense against inhalational anthrax in a murine model of infection. PLoS Pathog. 2010, 6, e1001199. [Google Scholar] [CrossRef] [PubMed]
- Crawford, M.A.; Lowe, D.E.; Fisher, D.J.; Stibitz, S.; Plaut, R.D.; Beaber, J.W.; Zemansky, J.; Mehrad, B.; Glomski, I.J.; Strieter, R.M.; et al. Identification of the bacterial protein FtsX as a unique target of chemokine-mediated antimicrobial activity against Bacillus anthracis. Proc. Natl. Acad. Sci. USA 2011, 108, 17159–17164. [Google Scholar] [CrossRef] [PubMed]
- Broder, C.C.; Collman, R.G. Chemokine receptors and HIV. J. Leukoc. Biol. 1997, 62, 20–29. [Google Scholar] [PubMed]
- Berger, E.A.; Murphy, P.M.; Farber, J.M. Chemokine receptors as HIV-1 coreceptors: Roles in viral entry, tropism, and disease. Annu. Rev. Immunol. 1999, 17, 657–700. [Google Scholar] [CrossRef] [PubMed]
- Yang, O.O.; Garcia-Zepeda, E.A.; Walker, B.D.; Luster, A.D. Monocyte chemoattractant protein-2 (CC chemokine ligand 8) inhibits replication of human immunodeficiency virus type 1 via CC chemokine receptor 5. J. Infect. Dis. 2002, 185, 1174–1178. [Google Scholar] [CrossRef] [PubMed]
- Choi, W.T.; An, J. Biology and clinical relevance of chemokines and chemokine receptors CXCR4 and CCR5 in human diseases. Exp. Biol. Med. 2011, 236, 637–647. [Google Scholar] [CrossRef] [PubMed]
- Kledal, T.N.; Rosenkilde, M.M.; Coulin, F.; Simmons, G.; Johnsen, A.H.; Alouani, S.; Power, C.A.; Luttichau, H.R.; Gerstoft, J.; Clapham, P.R.; et al. A broad-spectrum chemokine antagonist encoded by Kaposi’s sarcoma-associated herpesvirus. Science 1997, 277, 1656–1659. [Google Scholar] [CrossRef] [PubMed]
- Moore, J.P.; Trkola, A.; Dragic, T. Co-receptors for HIV-1 entry. Curr. Opin. Immunol. 1997, 9, 551–562. [Google Scholar] [CrossRef]
- Zhou, N.; Luo, Z.; Luo, J.; Hall, J.W.; Huang, Z. A novel peptide antagonist of CXCR4 derived from the N-terminus of viral chemokine vMIP-II. Biochemistry 2000, 39, 3782–3787. [Google Scholar] [CrossRef] [PubMed]
- Zhou, N.; Luo, Z.; Luo, J.; Fan, X.; Cayabyab, M.; Hiraoka, M.; Liu, D.; Han, X.; Pesavento, J.; Dong, C.Z.; et al. Exploring the stereochemistry of CXCR4-peptide recognition and inhibiting HIV-1 entry with d-peptides derived from chemokines. J. Biol. Chem. 2002, 277, 17476–17485. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Duggineni, S.; Espitia, S.; Richman, D.D.; An, J.; Huang, Z. A synthetic bivalent ligand of CXCR4 inhibits HIV infection. Biochem. Biophys. Res. Commun. 2013, 435, 646–650. [Google Scholar] [CrossRef] [PubMed]
- Detheux, M.; Ständker, L.; Vakili, J.; Münch, J.; Forssmann, U.; Adermann, K.; Pöhlmann, S.; Vassart, G.; Kirchhoff, F.; Parmentier, M.; et al. Natural proteolytic processing of hemofiltrate Cc chemokine 1 generates a potent Cc chemokine receptor (Ccr)1 and Ccr5 agonist with anti-HIV properties. J. Exp. Med. 2000, 192, 1501–1508. [Google Scholar] [CrossRef] [PubMed]
- Love, M.S.; Millholland, M.G.; Mishra, S.; Kulkarni, S.; Freeman, K.B.; Pan, W.; Kavash, R.W.; Costanzo, M.J.; Jo, H.; Daly, T.M.; et al. Platelet factor 4 activity against P. falciparum and its translation to nonpeptidic mimics as antimalarials. Cell Host Microbe 2012, 12, 815–823. [Google Scholar] [CrossRef] [PubMed]
- Sobirk, S.K.; Morgelin, M.; Egesten, A.; Bates, P.; Shannon, O.; Collin, M. Human chemokines as antimicrobial peptides with direct parasiticidal effect on Leishmania mexicana in vitro. PLoS ONE 2013, 8, e58129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwakman, P.H.; Krijgsveld, J.; de Boer, L.; Nguyen, L.T.; Boszhard, L.; Vreede, J.; Dekker, H.L.; Speijer, D.; Drijfhout, J.W.; te Velde, A.A.; et al. Native thrombocidin-1 and unfolded thrombocidin-1 exert antimicrobial activity via distinct structural elements. J. Biol. Chem. 2011, 286, 43506–43514. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.T.; Chan, D.I.; Boszhard, L.; Zaat, S.A.; Vogel, H.J. Structure-function studies of chemokine-derived carboxy-terminal antimicrobial peptides. Biochim. Biophys. Acta 2010, 1798, 1062–1072. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.T.; Kwakman, P.H.; Chan, D.I.; Liu, Z.; de Boer, L.; Zaat, S.A.; Vogel, H.J. Exploring platelet chemokine antimicrobial activity: Nuclear magnetic resonance backbone dynamics of NAP-2 and TC-1. Antimicrob. Agents Chemother. 2011, 55, 2074–2083. [Google Scholar] [CrossRef] [PubMed]
- Yeaman, M.R.; Yount, N.Y.; Waring, A.J.; Gank, K.D.; Kupferwasser, D.; Wiese, R.; Bayer, A.S.; Welch, W.H. Modular determinants of antimicrobial activity in platelet factor-4 family kinocidins. Biochim. Biophys. Acta 2007, 1768, 609–619. [Google Scholar] [CrossRef] [PubMed]
- Yount, N.Y.; Gank, K.D.; Xiong, Y.Q.; Bayer, A.S.; Pender, T.; Welch, W.H.; Yeaman, M.R. Platelet microbicidal protein 1: Structural themes of a multifunctional antimicrobial peptide. Antimicrob. Agents Chemother. 2004, 48, 4395–4404. [Google Scholar] [CrossRef] [PubMed]
- Walz, A.; Baggiolini, M. Generation of the neutrophil-activating peptide NAP-2 from platelet basic protein or connective tissue-activating peptide III through monocyte proteases. J. Exp. Med. 1990, 171, 449–454. [Google Scholar] [CrossRef] [PubMed]
- Dankert, J.; Krijgsveld, J.; van Der Werff, J.; Joldersma, W.; Zaat, S.A. Platelet microbicidal activity is an important defense factor against viridans streptococcal endocarditis. J. Infect. Dis. 2001, 184, 597–605. [Google Scholar] [CrossRef] [PubMed]
- Bjorstad, A.; Fu, H.; Karlsson, A.; Dahlgren, C.; Bylund, J. Interleukin-8-derived peptide has antibacterial activity. Antimicrob. Agents Chemother. 2005, 49, 3889–3895. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Becerra, F.; Silva, D.A.; Dominguez-Ramirez, L.; Mendoza-Hernandez, G.; Lopez-Vidal, Y.; Soldevila, G.; Garcia-Zepeda, E.A. Analysis of the antimicrobial activities of a chemokine-derived peptide (CDAP-4) on Pseudomonas aeruginosa. Biochem. Biophys. Res. Commun. 2007, 355, 352–358. [Google Scholar] [CrossRef] [PubMed]
- Erfe, M.C.B.; David, C.V.; Huang, C.; Lu, V.; Maretti-Mira, A.C.; Haskell, J.; Bruhn, K.W.; Yeaman, M.R.; Craft, N. Efficacy of synthetic peptides RP-1 and AA-RP-1 against Leishmania species in vitro and in vivo. Antimicrob. Agents Chemother. 2012, 56, 658–665. [Google Scholar] [CrossRef] [PubMed]
- Bourbigot, S.; Dodd, E.; Horwood, C.; Cumby, N.; Fardy, L.; Welch, W.H.; Ramjan, Z.; Sharma, S.; Waring, A.J.; Yeaman, M.R.; et al. Antimicrobial peptide RP-1 structure and interactions with anionic vs. zwitterionic micelles. Biopolymers 2009, 91, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Linge, H.M.; Collin, M.; Giwercman, A.; Malm, J.; Bjartell, A.; Egesten, A. The antibacterial chemokine MIG/CXCL9 is constitutively expressed in epithelial cells of the male urogenital tract and is present in seminal plasma. J. Interferon Cytokine Res. 2008, 28, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Proost, P.; Struyf, S.; Schols, D.; Opdenakker, G.; Sozzani, S.; Allavena, P.; Mantovani, A.; Augustyns, K.; Bal, G.; Haemers, A.; et al. Truncation of macrophage-derived chemokine by CD26/ dipeptidyl-peptidase IV beyond Its predicted cleavage site affects chemotactic activity and CC chemokine receptor 4 interaction. J. Biol. Chem. 1999, 274, 3988–3993. [Google Scholar] [CrossRef] [PubMed]
- Yount, N.Y.; Waring, A.J.; Gank, K.D.; Welch, W.H.; Kupferwasser, D.; Yeaman, M.R. Structural correlates of antimicrobial efficacy in IL-8 and related human kinocidins. Biochim. Biophys. Acta 2007, 1768, 598–608. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, B.O.; Wu, Z.; Nuding, S.; Groscurth, S.; Marcinowski, M.; Beisner, J.; Buchner, J.; Schaller, M.; Stange, E.F.; Wehkamp, J. Reduction of disulphide bonds unmasks potent antimicrobial activity of human beta-defensin 1. Nature 2011, 469, 419–423. [Google Scholar] [CrossRef] [PubMed]
- Leibovich-Rivkin, T.; Liubomirski, Y.; Bernstein, B.; Meshel, T.; Ben-Baruch, A. Inflammatory factors of the tumor microenvironment induce plasticity in nontransformed breast epithelial cells: EMT, invasion, and collapse of normally organized breast textures. Neoplasia 2013, 15, 1330–1346. [Google Scholar] [CrossRef] [PubMed]
- Viola, A.; Sarukhan, A.; Bronte, V.; Molon, B. The pros and cons of chemokines in tumor immunology. Trends Immunol. 2012, 33, 496–504. [Google Scholar] [CrossRef] [PubMed]
- Zlotnik, A. Involvement of chemokine receptors in organ-specific metastasis. Contrib. Microbiol. 2006, 13, 191–199. [Google Scholar] [PubMed]
- Ben-Baruch, A. Organ selectivity in metastasis: regulation by chemokines and their receptors. Clin. Exp. Metastasis 2008, 25, 345–356. [Google Scholar] [CrossRef] [PubMed]
- Muller, A.; Homey, B.; Soto, H.; Ge, N.; Catron, D.; Buchanan, M.E.; McClanahan, T.; Murphy, E.; Yuan, W.; Wagner, S.N.; et al. Involvement of chemokine receptors in breast cancer metastasis. Nature 2001, 410, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Lorusso, G.; Ruegg, C. New insights into the mechanisms of organ-specific breast cancer metastasis. Semin. Cancer Biol. 2012, 22, 226–233. [Google Scholar] [CrossRef] [PubMed]
- Valdivia-Silva, J.E.; Franco-Barraza, J.; Silva, A.L.; Pont, G.D.; Soldevila, G.; Meza, I.; Garcia-Zepeda, E.A. Effect of pro-inflammatory cytokine stimulation on human breast cancer: Implications of chemokine receptor expression in cancer metastasis. Cancer Lett. 2009, 283, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Franco-Barraza, J.; Valdivia-Silva, J.E.; Zamudio-Meza, H.; Castillo, A.; Garcia-Zepeda, E.A.; Benitez-Bribiesca, L.; Meza, I. Actin cytoskeleton participation in the onset of IL-1beta induction of an invasive mesenchymal-like phenotype in epithelial MCF-7 cells. Arch. Med. Res. 2010, 41, 170–181. [Google Scholar] [CrossRef] [PubMed]
- Valdivia-Silva, J.; Franco-Barraza, J.; Cukierman, E.; García-Zepeda, E.A. Novel insights Into the role of inflammation in promoting breast cancer development. In Breast Cancer—Focusing Tumor Microenvironment, Stem Cells and Metastasis; Gunduz, M., Gunduz, E., Eds.; InTech: Rijeka, Croatia, 2011; pp. 129–164. [Google Scholar]
- Karnoub, A.E.; Weinberg, R.A. Chemokine networks and breast cancer metastasis. Breast Dis. 2006, 26, 75–85. [Google Scholar] [PubMed]
- Zlotnik, A. Chemokines and cancer. Int. J. Cancer 2006, 119, 2026–2029. [Google Scholar] [CrossRef] [PubMed]
- Rahir, G.; Moser, M. Tumor microenvironment and lymphocyte infiltration. Cancer Immunol. Immunother. 2012, 61, 751–759. [Google Scholar] [CrossRef] [PubMed]
- Moser, B.; Willimann, K. Chemokines: Role in inflammation and immune surveillance. Ann. Rheum. Dis. 2004, 63 (Suppl. 2), ii84–ii89. [Google Scholar] [CrossRef] [PubMed]
- Koizumi, K.; Hojo, S.; Akashi, T.; Yasumoto, K.; Saiki, I. Chemokine receptors in cancer metastasis and cancer cell-derived chemokines in host immune response. Cancer Sci. 2007, 98, 1652–1658. [Google Scholar] [CrossRef] [PubMed]
- Fridlender, Z.G.; Sun, J.; Kim, S.; Kapoor, V.; Cheng, G.; Ling, L.; Worthen, G.S.; Albelda, S.M. Polarization of tumor-associated neutrophil phenotype by TGF-beta: “N1” vs. “N2” TAN. Cancer Cell 2009, 16, 183–194. [Google Scholar] [CrossRef] [PubMed]
- Maruyama, T.; Kono, K.; Izawa, S.; Mizukami, Y.; Kawaguchi, Y.; Mimura, K.; Watanabe, M.; Fujii, H. CCL17 and CCL22 chemokines within tumor microenvironment are related to infiltration of regulatory T cells in esophageal squamous cell carcinoma. Dis. Esophagus 2010, 23, 422–429. [Google Scholar] [CrossRef] [PubMed]
- Mizukami, H.; Shirahata, A.; Goto, T.; Sakata, M.; Saito, M.; Ishibashi, K.; Kigawa, G.; Nemoto, H.; Sanada, Y.; Hibi, K. PGP9.5 methylation as a marker for metastatic colorectal cancer. Anticancer Res. 2008, 28, 2697–2700. [Google Scholar] [PubMed]
- Musha, H.; Ohtani, H.; Mizoi, T.; Kinouchi, M.; Nakayama, T.; Shiiba, K.; Miyagawa, K.; Nagura, H.; Yoshie, O.; Sasaki, I. Selective infiltration of CCR5+CXCR3+ T lymphocytes in human colorectal carcinoma. Int. J. Cancer 2005, 116, 949–956. [Google Scholar] [CrossRef] [PubMed]
- Hojo, S.; Koizumi, K.; Tsuneyama, K.; Arita, Y.; Cui, Z.; Shinohara, K.; Minami, T.; Hashimoto, I.; Nakayama, T.; Sakurai, H.; et al. High-level expression of chemokine CXCL16 by tumor cells correlates with a good prognosis and increased tumor-infiltrating lymphocytes in colorectal cancer. Cancer Res. 2007, 67, 4725–4731. [Google Scholar] [CrossRef] [PubMed]
- Li, A.; Varney, M.L.; Valasek, J.; Godfrey, M.; Dave, B.J.; Singh, R.K. Autocrine role of interleukin-8 in induction of endothelial cell proliferation, survival, migration and MMP-2 production and angiogenesis. Angiogenesis 2005, 8, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Kryczek, I.; Lange, A.; Mottram, P.; Alvarez, X.; Cheng, P.; Hogan, M.; Moons, L.; Wei, S.; Zou, L.; Machelon, V.; et al. Cxcl12 and vascular endothelial growth factor synergistically induce neoangiogenesis in human ovarian cancers. Cancer Res. 2005, 65, 465–472. [Google Scholar] [PubMed]
- Vandercappellen, J.; van Damme, J.; Struyf, S. The role of CXC chemokines and their receptors in cancer. Cancer Lett. 2008, 267, 226–244. [Google Scholar] [CrossRef] [PubMed]
- Maurer, A.M.; Zhou, B.; Han, Z.C. Roles of platelet factor 4 in hematopoiesis and angiogenesis. Growth Factors 2006, 24, 242–252. [Google Scholar] [CrossRef] [PubMed]
- Wislez, M.; Fujimoto, N.; Izzo, J.G.; Hanna, A.E.; Cody, D.D.; Langley, R.R.; Tang, H.; Burdick, M.D.; Sato, M.; Minna, J.D.; et al. High expression of ligands for chemokine receptor CXCR2 in alveolar epithelial neoplasia induced by oncogenic kras. Cancer Res. 2006, 66, 4198–4207. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.Q.; Tsuda, Y.; Han, M.; Xu, D.H.; Kanagawa, N.; Hatanaka, Y.; Tani, Y.; Mizuguchi, H.; Tsutsumi, Y.; Mayumi, T.; et al. NK cells are migrated and indispensable in the anti-tumor activity induced by CCL27 gene therapy. Cancer Immunol. Immunother. 2009, 58, 291–299. [Google Scholar] [CrossRef] [PubMed]
- Tessema, M.; Klinge, D.M.; Yingling, C.M.; Do, K.; van Neste, L.; Belinsky, S.A. Re-expression of CXCL14, a common target for epigenetic silencing in lung cancer, induces tumor necrosis. Oncogene 2010, 29, 5159–5170. [Google Scholar] [CrossRef] [PubMed]
- Villablanca, E.J.; Raccosta, L.; Zhou, D.; Fontana, R.; Maggioni, D.; Negro, A.; Sanvito, F.; Ponzoni, M.; Valentinis, B.; Bregni, M.; et al. Tumor-mediated liver X receptor-alpha activation inhibits CC chemokine receptor-7 expression on dendritic cells and dampens antitumor responses. Nat. Med. 2010, 16, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Zlotnik, A.; Burkhardt, A.M.; Homey, B. Homeostatic chemokine receptors and organ-specific metastasis. Nat. Rev. Immunol. 2011, 11, 597–606. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, D.; Zhao, J. The Role of chemokine receptor CXCR4 in breast cancer metastasis. Am. J. Cancer Res. 2013, 3, 46–57. [Google Scholar] [PubMed]
- Ehtesham, M.; Mapara, K.Y.; Stevenson, C.B.; Thompson, R.C. CXCR4 mediates the proliferation of glioblastoma progenitor cells. Cancer Lett. 2009, 274, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Verbeke, H.; Struyf, S.; Laureys, G.; van Damme, J. The expression and role of CXC chemokines in colorectal cancer. Cytokine Growth Factor Rev. 2011, 22, 345–358. [Google Scholar] [CrossRef] [PubMed]
- Mohle, R.; Failenschmid, C.; Bautz, F.; Kanz, L. Overexpression of the chemokine receptor CXCR4 in B cell chronic lymphocytic leukemia is associated with increased functional response to stromal cell-derived factor-1 (SDF-1). Leukemia 1999, 13, 1954–1959. [Google Scholar] [CrossRef] [PubMed]
- Yun, J.A.; Kim, H.C.; Kim, S.H.; Cho, Y.B.; Yun, S.H.; Lee, W.Y.; Chun, H.K. Prognostic significance of perineural invasion in stage IIA colon cancer. ANZ J. Surg. 2014. [Google Scholar] [CrossRef] [PubMed]
- Kawada, K.; Hosogi, H.; Sonoshita, M.; Sakashita, H.; Manabe, T.; Shimahara, Y.; Sakai, Y.; Takabayashi, A.; Oshima, M.; Taketo, M.M. Chemokine receptor CXCR3 promotes colon cancer metastasis to lymph nodes. Oncogene 2007, 26, 4679–4688. [Google Scholar] [CrossRef] [PubMed]
- Saur, D.; Seidler, B.; Schneider, G.; Algul, H.; Beck, R.; Senekowitsch-Schmidtke, R.; Schwaiger, M.; Schmid, R.M. CXCR4 expression increases liver and lung metastasis in a mouse model of pancreatic cancer. Gastroenterology 2005, 129, 1237–1250. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Loberg, R.; Taichman, R.S. The pivotal role of CXCL12 (SDF-1)/CXCR4 axis in bone metastasis. Cancer Metastasis Rev. 2006, 25, 573–587. [Google Scholar] [CrossRef] [PubMed]
- Ghadjar, P.; Rubie, C.; Aebersold, D.M.; Keilholz, U. The chemokine CCL20 and its receptor CCR6 in human malignancy with focus on colorectal cancer. Int. J. Cancer 2009, 125, 741–745. [Google Scholar] [CrossRef] [PubMed]
- Raynaud, C.M.; Mercier, O.; Dartevelle, P.; Commo, F.; Olaussen, K.A.; de Montpreville, V.; Andre, F.; Sabatier, L.; Soria, J.C. Expression of chemokine receptor CCR6 as a molecular determinant of adrenal metastatic relapse in patients with primary lung cancer. Clin. Lung Cancer 2010, 11, 187–191. [Google Scholar] [CrossRef] [PubMed]
- Klein, A.; Sagi-Assif, O.; Izraely, S.; Meshel, T.; Pasmanik-Chor, M.; Nahmias, C.; Couraud, P.O.; Erez, N.; Hoon, D.S.; Witz, I.P. The metastatic microenvironment: Brain-derived soluble factors alter the malignant phenotype of cutaneous and brain-metastasizing melanoma cells. Int. J. Cancer 2012, 131, 2509–2518. [Google Scholar] [CrossRef] [PubMed]
- Amersi, F.F.; Terando, A.M.; Goto, Y.; Scolyer, R.A.; Thompson, J.F.; Tran, A.N.; Faries, M.B.; Morton, D.L.; Hoon, D.S. Activation of CCR9/CCL25 in cutaneous melanoma mediates preferential metastasis to the small intestine. Clin. Cancer Res. 2008, 14, 638–645. [Google Scholar] [CrossRef] [PubMed]
- Letsch, A.; Keilholz, U.; Schadendorf, D.; Assfalg, G.; Asemissen, A.M.; Thiel, E.; Scheibenbogen, C. Functional CCR9 expression is associated with small intestinal metastasis. J. Investig. Dermatol. 2004, 122, 685–690. [Google Scholar] [CrossRef] [PubMed]
- Meijer, J.; Ogink, J.; Roos, E. Effect of the chemokine receptor CXCR7 on proliferation of carcinoma cells in vitro and in vivo. Br. J. Cancer 2008, 99, 1493–1501. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.; Schuster, R.; Stringer, K.F.; Waltz, S.E.; Lentsch, A.B. The duffy antigen/receptor for chemokines (DARC) regulates prostate tumor growth. FASEB J. 2006, 20, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Bonecchi, R.; Savino, B.; Borroni, E.M.; Mantovani, A.; Locati, M. Chemokine decoy receptors: Structure-function and biological properties. Curr. Top. Microbiol. Immunol. 2010, 341, 15–36. [Google Scholar] [PubMed]
- Bobisse, S.; Rondina, M.; Merlo, A.; Tisato, V.; Mandruzzato, S.; Amendola, M.; Naldini, L.; Willemsen, R.A.; Debets, R.; Zanovello, P.; et al. Reprogramming T lymphocytes for melanoma adoptive immunotherapy by T-cell receptor gene transfer with lentiviral vectors. Cancer Res. 2009, 69, 9385–9394. [Google Scholar] [CrossRef] [PubMed]
- Baratelli, F.; Takedatsu, H.; Hazra, S.; Peebles, K.; Luo, J.; Kurimoto, P.S.; Zeng, G.; Batra, R.K.; Sharma, S.; Dubinett, S.M.; et al. Pre-clinical characterization of GMP grade CCL21-gene modified dendritic cells for application in a phase I trial in non-small cell lung cancer. J. Transl. Med. 2008, 6, 38. [Google Scholar] [CrossRef] [PubMed]
- Di Stasi, A.; de Angelis, B.; Rooney, C.M.; Zhang, L.; Mahendravada, A.; Foster, A.E.; Heslop, H.E.; Brenner, M.K.; Dotti, G.; Savoldo, B. T lymphocytes coexpressing CCR4 and a chimeric antigen receptor targeting CD30 have improved homing and antitumor activity in a Hodgkin tumor model. Blood 2009, 113, 6392–6402. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.; Lazennec, G. Chemokines: Novel targets for breast cancer metastasis. Cancer Metastasis Rev. 2007, 26, 401–420. [Google Scholar] [CrossRef] [PubMed]
- Arenberg, D.A.; Keane, M.P.; DiGiovine, B.; Kunkel, S.L.; Morris, S.B.; Xue, Y.Y.; Burdick, M.D.; Glass, M.C.; Iannettoni, M.D.; Strieter, R.M. Epithelial-neutrophil activating peptide (ENA-78) is an important angiogenic factor in non-small cell lung cancer. J. Clin. Investig. 1998, 102, 465–472. [Google Scholar] [CrossRef] [PubMed]
- Beider, K.; Abraham, M.; Begin, M.; Wald, H.; Weiss, I.D.; Wald, O.; Pikarsky, E.; Abramovitch, R.; Zeira, E.; Galun, E.; et al. Interaction between CXCR4 and CCL20 pathways regulates tumor growth. PLoS ONE 2009, 4, e5125. [Google Scholar] [CrossRef] [PubMed]
- Loberg, R.D.; Ying, C.; Craig, M.; Day, L.L.; Sargent, E.; Neeley, C.; Wojno, K.; Snyder, L.A.; Yan, L.; Pienta, K.J. Targeting CCL2 with systemic delivery of neutralizing antibodies induces prostate cancer tumor regression in vivo. Cancer Res. 2007, 67, 9417–9424. [Google Scholar] [CrossRef] [PubMed]
- Garber, K. First results for agents targeting cancer-related inflammation. J. Natl. Cancer Inst. 2009, 101, 1110–1112. [Google Scholar] [CrossRef] [PubMed]
- Sandhu, S.K.; Papadopoulos, K.; Fong, P.C.; Patnaik, A.; Messiou, C.; Olmos, D.; Wang, G.; Tromp, B.J.; Puchalski, T.A.; Balkwill, F.; et al. A first-in-human, first-in-class, phase I study of carlumab (CNTO 888), a human monoclonal antibody against CC-chemokine ligand 2 in patients with solid tumors. Cancer Chemother. Pharmacol. 2013, 71, 1041–1050. [Google Scholar] [CrossRef] [PubMed]
- Rozel, S.; Galban, C.J.; Nicolay, K.; Lee, K.C.; Sud, S.; Neeley, C.; Snyder, L.A.; Chenevert, T.L.; Rehemtulla, A.; Ross, B.D.; et al. Synergy between anti-CCL2 and docetaxel as determined by DW-MRI in a metastatic bone cancer model. J. Cell. Biochem. 2009, 107, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Izhak, L.; Wildbaum, G.; Weinberg, U.; Shaked, Y.; Alami, J.; Dumont, D.; Friedman, B.; Stein, A.; Karin, N. Predominant expression of CCL2 at the tumor site of prostate cancer patients directs a selective loss of immunological tolerance to CCL2 that could be amplified in a beneficial manner. J. Immunol. 2010, 184, 1092–1101. [Google Scholar] [CrossRef] [PubMed]
- Domanska, U.M.; Kruizinga, R.C.; Nagengast, W.B.; Timmer-Bosscha, H.; Huls, G.; de Vries, E.G.; Walenkamp, A.M. A review on CXCR4/CXCL12 axis in oncology: No place to hide. Eur. J. Cancer 2013, 49, 219–230. [Google Scholar] [CrossRef] [PubMed]
- Heveker, N.; Tissot, M.; Thuret, A.; Schneider-Mergener, J.; Alizon, M.; Roch, M.; Marullo, S. Pharmacological properties of peptides derived from stromal cell-derived factor 1: Study on human polymorphonuclear cells. Mol. Pharmacol. 2001, 59, 1418–1425. [Google Scholar] [PubMed]
- Wong, D.; Korz, W. Translating an Antagonist of Chemokine Receptor CXCR4: From bench to bedside. Clin. Cancer Res. 2008, 14, 7975–7980. [Google Scholar] [CrossRef] [PubMed]
- Duda, D.G.; Kozin, S.V.; Kirkpatrick, N.D.; Xu, L.; Fukumura, D.; Jain, R.K. CXCL12 (SDF1alpha)-CXCR4/CXCR7 pathway inhibition: an emerging sensitizer for anticancer therapies? Clin. Cancer Res. 2011, 17, 2074–2080. [Google Scholar] [CrossRef] [PubMed]
- Broxmeyer, H.E.; Orschell, C.M.; Clapp, D.W.; Hangoc, G.; Cooper, S.; Plett, P.A.; Liles, W.C.; Li, X.; Graham-Evans, B.; Campbell, T.B.; et al. Rapid mobilization of murine and human hematopoietic stem and progenitor cells with AMD3100, a CXCR4 antagonist. J. Exp. Med. 2005, 201, 1307–1318. [Google Scholar] [CrossRef] [PubMed]
- De Nigris, F.; Schiano, C.; Infante, T.; Napoli, C. CXCR4 inhibitors: tumor vasculature and therapeutic challenges. Recent Pat. Anticancer Drug Discov. 2012, 7, 251–264. [Google Scholar] [CrossRef] [PubMed]
- Kwong, J.; Kulbe, H.; Wong, D.; Chakravarty, P.; Balkwill, F. An antagonist of the chemokine receptor CXCR4 induces mitotic catastrophe in ovarian cancer cells. Mol. Cancer Ther. 2009, 8, 1893–1905. [Google Scholar] [CrossRef] [PubMed]
- Porvasnik, S.; Sakamoto, N.; Kusmartsev, S.; Eruslanov, E.; Kim, W.J.; Cao, W.; Urbanek, C.; Wong, D.; Goodison, S.; Rosser, C.J. Effects of CXCR4 antagonist CTCE-9908 on prostate tumor growth. Prostate 2009, 69, 1460–1469. [Google Scholar] [CrossRef] [PubMed]
- Tamamura, H.; Imai, M.; Ishihara, T.; Masuda, M.; Funakoshi, H.; Oyake, H.; Murakami, T.; Arakaki, R.; Nakashima, H.; Otaka, A.; et al. Pharmacophore identification of a chemokine receptor (CXCR4) antagonist, T22 ([Tyr5,12, Lys7]-polyphemusin II), which specifically blocks T cell-line-tropic HIV-1 infection. Bioorg. Med. Chem. 1998, 6, 1033–1041. [Google Scholar] [CrossRef]
- Peled, A.; Abraham, M.; Avivi, I.; Rowe, J.M.; Beider, K.; Wald, H.; Tiomkin, L.; Ribakovsky, L.; Riback, Y.; Ramati, Y.; et al. The high-affinity CXCR4 antagonist BKT140 is safe and induces a robust mobilization of human CD34+ cells in patients with multiple myeloma. Clin. Cancer Res. 2014, 20, 469–479. [Google Scholar] [CrossRef] [PubMed]
- Ling, X.; Spaeth, E.; Chen, Y.; Shi, Y.; Zhang, W.; Schober, W.; Hail, N., Jr.; Konopleva, M.; Andreeff, M. The CXCR4 antagonist AMD3465 regulates oncogenic signaling and invasiveness in vitro and prevents breast cancer growth and metastasis in vivo. PLoS ONE 2013, 8, e58426. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.K.; Arya, R.K.; Trivedi, A.K.; Sanyal, S.; Baral, R.; Dormond, O.; Briscoe, D.M.; Datta, D. Chemokine receptor trio: CXCR3, CXCR4 and CXCR7 crosstalk via CXCL11 and CXCL12. Cytokine Growth Factor Rev. 2013, 24, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Egorova, A.; Kiselev, A.; Hakli, M.; Ruponen, M.; Baranov, V.; Urtti, A. Chemokine-derived peptides as carriers for gene delivery to CXCR4 expressing cells. J. Gene Med. 2009, 11, 772–781. [Google Scholar] [CrossRef] [PubMed]
- Sebag, M. CCR1 blockade and myeloma bone disease. Blood 2012, 120, 1351–1352. [Google Scholar] [CrossRef] [PubMed]
- Kitamura, T.; Fujishita, T.; Loetscher, P.; Revesz, L.; Hashida, H.; Kizaka-Kondoh, S.; Aoki, M.; Taketo, M.M. Inactivation of chemokine (C-C motif) receptor 1 (CCR1) suppresses colon cancer liver metastasis by blocking accumulation of immature myeloid cells in a mouse model. Proc. Natl. Acad. Sci. USA 2010, 107, 13063–13068. [Google Scholar] [CrossRef] [PubMed]
- Robinson, S.C.; Scott, K.A.; Wilson, J.L.; Thompson, R.G.; Proudfoot, A.E.; Balkwill, F.R. A chemokine receptor antagonist inhibits experimental breast tumor growth. Cancer Res. 2003, 63, 8360–8365. [Google Scholar] [PubMed]
- Yamamoto, K.; Utsunomiya, A.; Tobinai, K.; Tsukasaki, K.; Uike, N.; Uozumi, K.; Yamaguchi, K.; Yamada, Y.; Hanada, S.; Tamura, K.; et al. Phase I study of KW-0761, a defucosylated humanized anti-CCR4 antibody, in relapsed patients with adult T-cell leukemia-lymphoma and peripheral T-cell lymphoma. J. Clin. Oncol. 2010, 28, 1591–1598. [Google Scholar] [CrossRef] [PubMed]
- Ogura, M.; Ishida, T.; Hatake, K.; Taniwaki, M.; Ando, K.; Tobinai, K.; Fujimoto, K.; Yamamoto, K.; Miyamoto, T.; Uike, N.; et al. Multicenter phase II study of mogamulizumab (KW-0761), a defucosylated anti-CC chemokine receptor 4 antibody, in patients With relapsed peripheral T-cell lymphoma and cutaneous T-cell lymphoma. J. Clin. Oncol. 2014, 32, 1157–1163. [Google Scholar] [CrossRef] [PubMed]
- Rosca, E.V.; Lal, B.; Koskimaki, J.E.; Popel, A.S.; Laterra, J. Collagen IV and CXC chemokine-derived antiangiogenic peptides suppress glioma xenograft growth. Anticancer Drugs 2012, 23, 706–712. [Google Scholar] [CrossRef] [PubMed]
- Karagiannis, E.D.; Popel, A.S. Novel anti-angiogenic peptides derived from ELR-containing CXC chemokines. J. Cell. Biochem. 2008, 104, 1356–1363. [Google Scholar] [CrossRef] [PubMed]
- Koskimaki, J.E.; Karagiannis, E.D.; Rosca, E.V.; Vesuna, F.; Winnard, P.T., Jr.; Raman, V.; Bhujwalla, Z.M.; Popel, A.S. Peptides derived from type IV collagen, CXC chemokines, and thrombospondin-1 domain-containing proteins inhibit neovascularization and suppress tumor growth in MDA-MB-231 breast cancer xenografts. Neoplasia 2009, 11, 1285–1291. [Google Scholar] [CrossRef] [PubMed]
- Vandercappellen, J.; van Damme, J.; Struyf, S. The role of the CXC chemokines platelet factor-4 (CXCL4/PF-4) and its variant (CXCL4L1/PF-4var) in inflammation, angiogenesis and cancer. Cytokine Growth Factor Rev. 2011, 22, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Huang, H. Platelet factor-4 (CXCL4/PF-4): An angiostatic chemokine for cancer therapy. Cancer Lett. 2013, 331, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Kuo, J.-H.; Chen, Y.-P.; Liu, J.-S.; Dubrac, A.; Quemener, C.; Prats, H.; Bikfalvi, A.; Wu, W.-G.; Sue, S.-C. Alternative C-terminal helix orientation alters chemokine function: Structure of the anti-angiogenic chemokin, CXCL4L1. J. Biol. Chem. 2013, 288, 13522–13533. [Google Scholar] [CrossRef] [PubMed]
- Dubrac, A.; Quemener, C.; Lacazette, E.; Lopez, F.; Zanibellato, C.; Wu, W.-G.; Bikfalvi, A.; Prats, H. Functional divergence between 2 chemokines is conferred by single amino acid change. Blood 2010, 116, 4703–4711. [Google Scholar] [CrossRef] [PubMed]
- Struyf, S.; Burdick, M.D.; Proost, P.; van Damme, J.; Strieter, R.M. Platelets release CXCL4L1, a nonallelic variant of the chemokine platelet factor-4/CXCL4 and potent inhibitor of angiogenesis. Circ. Res. 2004, 95, 855–857. [Google Scholar] [CrossRef] [PubMed]
- Struyf, S.; Salogni, L.; Burdick, M.D.; Vandercappellen, J.; Gouwy, M.; Noppen, S.; Proost, P.; Opdenakker, G.; Parmentier, M.; Gerard, C.; et al. Angiostatic and chemotactic activities of the CXC chemokine CXCL4L1 (platelet factor-4 variant) are mediated by CXCR3. Blood 2011, 117, 480–488. [Google Scholar] [CrossRef] [PubMed]
- De Chaisemartin, L.; Goc, J.; Damotte, D.; Validire, P.; Magdeleinat, P.; Alifano, M.; Cremer, I.; Fridman, W.H.; Sautes-Fridman, C.; Dieu-Nosjean, M.C. Characterization of chemokines and adhesion molecules associated with T cell presence in tertiary lymphoid structures in human lung cancer. Cancer Res. 2011, 71, 6391–6399. [Google Scholar] [CrossRef] [PubMed]
- Vandercappellen, J.; Liekens, S.; Bronckaers, A.; Noppen, S.; Ronsse, I.; Dillen, C.; Belleri, M.; Mitola, S.; Proost, P.; Presta, M.; et al. The COOH-terminal peptide of platelet factor-4 variant (CXCL4L1/PF-4var47–70) strongly inhibits angiogenesis and suppresses B16 melanoma growth in vivo. Mol. Cancer Res. 2010, 8, 322–334. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Singh, A.P.; Sharma, B.; Owen, L.B.; Singh, R.K. CXCL8 and its cognate receptors in melanoma progression and metastasis. Future Oncol. 2010, 6, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Varney, M.L.; Singh, S.; Li, A.; Mayer-Ezell, R.; Bond, R.; Singh, R.K. Small molecule antagonists for CXCR2 and CXCR1 inhibit human colon cancer liver metastases. Cancer Lett. 2011, 300, 180–188. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Tressel, S.L.; Kaimal, R.; Balla, M.; Lam, F.H.; Covic, L.; Kuliopulos, A. Identification of a metalloprotease-chemokine signaling system in the ovarian cancer microenvironment: implications for antiangiogenic therapy. Cancer Res. 2010, 70, 5880–5890. [Google Scholar] [CrossRef] [PubMed]
- Seaton, A.; Maxwell, P.J.; Hill, A.; Gallagher, R.; Pettigrew, J.; Wilson, R.H.; Waugh, D.J. Inhibition of constitutive and cxc-chemokine-induced NF-κB activity potentiates ansamycin-based HSP90-inhibitor cytotoxicity in castrate-resistant prostate cancer cells. Br. J. Cancer 2009, 101, 1620–1629. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Yang, X.; Xu, W.; Li, K.; Chu, Y.; Xiong, S. Integrating individual functional moieties of CXCL10 and CXCL11 into a novel chimeric chemokine leads to synergistic antitumor effects: A strategy for chemokine-based multi-target-directed cancer therapy. Cancer Immunol. Immunother. 2010, 59, 1715–1726. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valdivia-Silva, J.; Medina-Tamayo, J.; Garcia-Zepeda, E.A. Chemokine-Derived Peptides: Novel Antimicrobial and Antineoplasic Agents. Int. J. Mol. Sci. 2015, 16, 12958-12985. https://doi.org/10.3390/ijms160612958
Valdivia-Silva J, Medina-Tamayo J, Garcia-Zepeda EA. Chemokine-Derived Peptides: Novel Antimicrobial and Antineoplasic Agents. International Journal of Molecular Sciences. 2015; 16(6):12958-12985. https://doi.org/10.3390/ijms160612958
Chicago/Turabian StyleValdivia-Silva, Julio, Jaciel Medina-Tamayo, and Eduardo A. Garcia-Zepeda. 2015. "Chemokine-Derived Peptides: Novel Antimicrobial and Antineoplasic Agents" International Journal of Molecular Sciences 16, no. 6: 12958-12985. https://doi.org/10.3390/ijms160612958
APA StyleValdivia-Silva, J., Medina-Tamayo, J., & Garcia-Zepeda, E. A. (2015). Chemokine-Derived Peptides: Novel Antimicrobial and Antineoplasic Agents. International Journal of Molecular Sciences, 16(6), 12958-12985. https://doi.org/10.3390/ijms160612958