
Structure and Principal Components Analyses Reveal an Intervarietal Fusion in Malaysian Mistletoe Fig (Ficus deltoidea Jack) Populations

Abstract
:1. Introduction
2. Results and Discussion
2.1. ANOVA and Mean Comparison of the Morphological Characteristics

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of Variation | DF | Mean of Squares | |||||||
|---|---|---|---|---|---|---|---|---|---|
| LL | LW | LA | PL | GS | FL | FD | FS | ||
| Between groups | 29 | 27.88 ** | 38.45 ** | 4122.83 ** | 13.62 ** | 4408.66 ** | 2.62 ** | 1.5 ** | 1.08 ** |
| Within groups | 270 | 0.29 | 0.27 | 48.74 | 0.22 | 54.27 | 0.02 | 0.00 | 0.02 |
| Accessions | LL | LW | LA | PL | GS | FL | FD | FS |
|---|---|---|---|---|---|---|---|---|
| ANG | 4.89 ± 0.72 j−o | 2.03 ± 0.22 m | 7.57 ± 1.35 mn | 0.54 ± 0.17 jk | 12.72 ± 3.34 j−n | 0.64 ± 0.08 l−p | 0.48 ± 0.07 k−o | 0.77 ± 0.14 e−h |
| FDAT1 | 4.34 ± 0.32 n | 2.06 ± 0.20 m | 7.59 ± 1.01 mn | 0.60 ± 0.15 jk | 11.50 ± 2.46 l−n | 0.69 ± 0.05 lm | 0.53 ± 0.10 k−m | 0.76 ± 0.08 e−h |
| FDAT2 | 5.77 ± 0.44 jk | 2.02 ± 0.21 m | 8.77 ± 1.42 mn | 0.68 ± 0.17 jk | 14.60 ± 1.26 j−n | 0.66 ± 0.04 l−n | 0.43 ± 0.04 no | 0.88 ± 0.06 ef |
| FDAT3 | 5.26 ± 0.45 k–m | 1.93 ± 0.15 m | 7.74 ± 1.16 mn | 0.35 ± 0.07 l | 8.80 ± 2.15 n | 0.55 ± 0.06 n−p | 0.45 ± 0.06 m−o | 0.64 ± 0.05 h−j |
| FDAK4 | 4.29 ± 0.23 n | 1.81 ± 0.20 m | 7.53 ± 1.23 mn | 0.44 ± 0.07 jk | 13.30 ± 2.98 k−n | 0.62 ± 0.06 l−o | 0.57 ± 0.03 kl | 0.75 ± 0.10 e−h |
| FDAT5 | 4.19 ± 0.19 no | 2.27 ± 0.08 m | 5.74 ± 0.50 mn | 0.47 ± 0.11 jk | 11.90 ± 2.51 l−n | 0.58 ± 0.10 m−o | 0.41 ± 0.02 o | 0.78 ± 0.14 e−h |
| FDAT6 | 5.48 ± 0.35 kl | 2.06 ± 0.17 m | 8.06 ± 0.36 n | 0.66 ± 0.16 jk | 16.20 ± 2.90 j−n | 0.71 ± 0.05 lm | 0.50 ± 0.03 mn | 0.82 ± 0.24 e−g |
| BIL | 3.41 ± 0.21 p | 2.10 ± 0.21 m | 3.77 ± 0.63 mn | 0.36 ± 0.07 k | 13.50 ± 2.27 k−n | 0.45 ± 0.06 p | 0.32 ± 0.04 p | 0.42 ± 0.13 kl |
| FDBK7 | 3.41 ± 0.21 p | 2.10 ± 0.21 m | 3.77 ± 0.63 mn | 0.36 ± 0.07 k | 13.50 ± 2.27 k−n | 0.45 ± 0.06 p | 0.32 ± 0.04 p | 0.42 ± 0.13 kl |
| DEL | 4.08 ± 0.68 m−p | 3.29 ± 0.32 kl | 8.12 ± 1.78 mn | 0.72 ± 0.20 i−k | 10.97 ± 2.86 k−n | 0.65 ± 0.10 l−p | 0.61 ± 0.10 j−l | 0.50 ± 0.12 i−l |
| FDDP8 | 3.75 ± 0.30 op | 3.27 ± 0.33 l | 7.09 ± 1.31 n | 0.57 ± 0.12 jk | 10.50 ± 3.06 mn | 0.71 ± 0.07 lm | 0.56 ± 0.05 kl | 0.59 ± 0.08 ij |
| FDDJ9 | 4.84 ± 0.52 m | 3.27 ± 0.28 l | 9.78 ± 0.87 mn | 0.91 ± 0.18 ij | 9.40 ± 2.01 mn | 0.72 ± 0.03 l | 0.68 ± 0.10 j | 0.53 ± 0.07 jk |
| FDDJ10 | 3.65 ± 0.36 p | 3.34 ± 0.37 kl | 7.49 ± 1.75 mn | 0.69 ± 0.12 jk | 13.00 ± 2.31 k−n | 0.52 ± 0.03 op | 0.59 ± 0.08 k | 0.37 ± 0.07 l |
| TRE | 6.33 ± 0.86 b−l | 3.86 ± 0.55 i−l | 22.13 ± 6.38 j−m | 2.20 ± 0.22 b−h | 22.97 ± 7.89 b−m | 1.40 ± 0.84 e−k | 0.97 ± 0.10 d−i | 1.20 ± 0.34 a−g |
| FDTT11 | 5.40 ± 0.50 kl | 4.06 ± 0.53 ij | 21.07 ± 4.53 k | 1.39 ± 0.47 h | 21.60 ± 7.26 h−j | 1.62 ± 0.20 ef | 0.98 ± 0.10 f−h | 1.43 ± 0.20 b |
| FDTT12 | 5.52 ± 0.36 kl | 3.72 ± 0.24 j−l | 19.63 ± 3.67 kl | 2.49 ± 0.47 ef | 25.10 ± 7.16 g−i | 1.60 ± 0.10 ef | 0.94 ± 0.07 g−i | 1.21 ± 0.21 c |
| FDTT13 | 6.61 ± 0.45 g–i | 3.62 ± 0.45 j−l | 22.61 ± 4.46 jk | 2.03 ± 0.62 g | 18.50 ± 5.38 i−l | 1.35 ± 0.29 ij | 1.05 ± 0.08 ef | 0.84 ± 0.09 ef |
| FDTT14 | 6.96 ± 0.46 f–h | 4.03 ± 0.42 ij | 26.01 ± 4.99 h−k | 1.88 ± 0.45 g | 24.40 ± 3.75 g−i | 1.25 ± 0.09 jk | 0.94 ± 0.12 g−i | 1.19 ± 0.15 cd |
| FDTT15 | 7.20 ± 0.41 ef | 3.84 ± 0.25 jk | 28.21 ± 3.97 h–j | 3.08 ± 0.33 b–d | 34.50 ± 6.65 ef | 1.40 ± 0.10 hi | 0.87 ± 0.03 i | 1.18 ± 0.06 cd |
| FDTT16 | 6.89 ± 0.43 f–h | 4.51 ± 0.45 i | 23.71 ± 7.92 i−k | 3.10 ± 0.38 b−d | 20.10 ± 6.74 h−k | 1.41 ± 0.09 g−i | 0.94 ± 0.10 g−i | 1.72 ± 0.33 a |
| FDTK17 | 5.75 ± 0.82 bjk | 3.23 ± 0.48 l | 13.63 ± 2.40 lm | 1.44 ± 0.84 h | 16.60 ± 2.76 j−m | 1.16 ± 0.10 k | 1.09 ± 0.05 de | 0.82 ± 0.08 e−g |
| KUN | 7.56 ± 1.35 a−m | 6.41 ± 1.21 a−h | 48.36 ± 15.44 a−i | 2.67 ±1.05 a−i | 49.88 ± 19.93 a−h | 1.71 ± 0.25 a−k | 1.29 ± 0.26 a−i | 0.97 ± 0.31 a−i |
| FDKT18 | 5.08 ± 0.39 lm | 5.51 ± 0.42 fg | 30.37 ± 2.62 g−i | 1.21 ± 0.33 hi | 26.80 ± 7.00 gh | 1.53 ± 0.09 fg | 0.91 ± 0.05 hi | 1.06 ± 0.15 d |
| FDKT19 | 6.52 ± 0.54 hi | 5.55 ± 0.57 fg | 35.96 ± 7.25 fg | 3.48 ± 0.64 b | 31.50 ± 5.36 fg | 1.28 ± 0.14 jk | 1.29 ± 0.09 c | 1.72 ± 0.10 a |
| FDKT20 | 6.12 ± 0.68 ij | 5.02 ± 0.80 h | 32.69 ± 4.50 f−h | 2.01 ± 0.33 g | 30.90 ± 11.27 fg | 1.68 ± 0.13 de | 1.00 ± 0.09 fg | 0.90 ± 0.17 e |
| FDKT21 | 7.10 ± 0.56 e–g | 6.01 ± 0.77 ef | 44.23 ± 9.52 c−e | 2.92 ± 0.64 c−e | 63.30 ± 8.96 b | 1.81 ± 0.17 bc | 1.28 ± 0.10 c | 1.28±0.22 c |
| FDKT22 | 8.38 ± 0.64 bc | 8.40 ± 0.50 a | 68.04 ± 10.18 a | 3.26 ± 0.58 bc | 61.80 ± 11.92 b | 1.80 ± 0.10 b−d | 1.63 ± 0.14 a | 0.90 ± 0.08 e |
| FDKK23 | 6.59 ± 0.76 g–i | 5.32 ± 0.54 gh | 39.07 ± 7.85 ef | 1.24 ± 0.26 hi | 44.00 ± 6.72 cd | 1.70 ± 0.40 c−e | 0.98 ± 0.16 f−h | 0.89 ± 0.15 e |
| FDKT24 | 9.31 ± 0.56 a | 5.98 ± 0.45 ef | 65.01 ± 8.29 ab | 2.65 ± 0.87 de | 63.70 ± 6.78 b | 2.10 ± 0.16 a | 1.67 ± 0.14 a | 0.81 ± 0.08 e−g |
| FDKT25 | 8.57 ± 0.73 b | 5.85 ± 0.59 f | 42.83 ± 14.07 de | 4.66 ± 0.55 a | 45.10 ± 4.25 cd | 1.70 ± 0.10 c−e | 1.52 ± 0.10 f−h | 1.21 ± 0.12 c |
| FDKT26 | 8.02 ± 0.54 cd | 6.75 ± 0.66 cd | 59.50 ± 13.49 b | 3.08 ± 0.56 b−d | 92.70 ± 20.15 a | 1.61 ± 0.11 ef | 1.45 ± 0.11 b | 0.68 ± 0.23 g−i |
| FDKT27 | 8.71 ± 1.06 b | 6.45 ± 1.34 de | 50.79 ± 15.98 c | 2.98 ± 0.64 cd | 46.50 ± 12.96 c | 1.50 ± 0.02 f−h | 1.12 ± 0.04 de | 0.73 ± 0.12 f−h |
| FDKT28 | 8.82 ± 0.77 b | 7.08 ± 0.76 c | 48.83 ± 6.63 cd | 2.49 ± 0.49 ef | 38.70 ± 9.04 de | 1.92 ± 0.17 b | 1.26 ± 0.09 c | 0.75 ± 0.14 e−h |
| FDKK29 | 7.59 ± 0.81 de | 7.61 ± 1.02 b | 46.84 ± 7.63 cd | 2.08 ± 0.61 fg | 43.00 ± 2.75 cd | 1.89 ± 0.03 b | 1.47 ± 0.04 b | 0.81 ± 0.08 e−g |
| FDKK30 | 7.54 ± 0.44 de | 7.61 ± 0.32 b | 64.51 ± 12.05 ab | 2.68 ± 0.72 de | 60.50 ± 11.16 b | 1.69 ± 0.09 c−e | 1.16 ± 0.09 a | 0.85 ± 0.12 ef |

2.2. Principal Components Analyses (PCA) Using Morphological Data
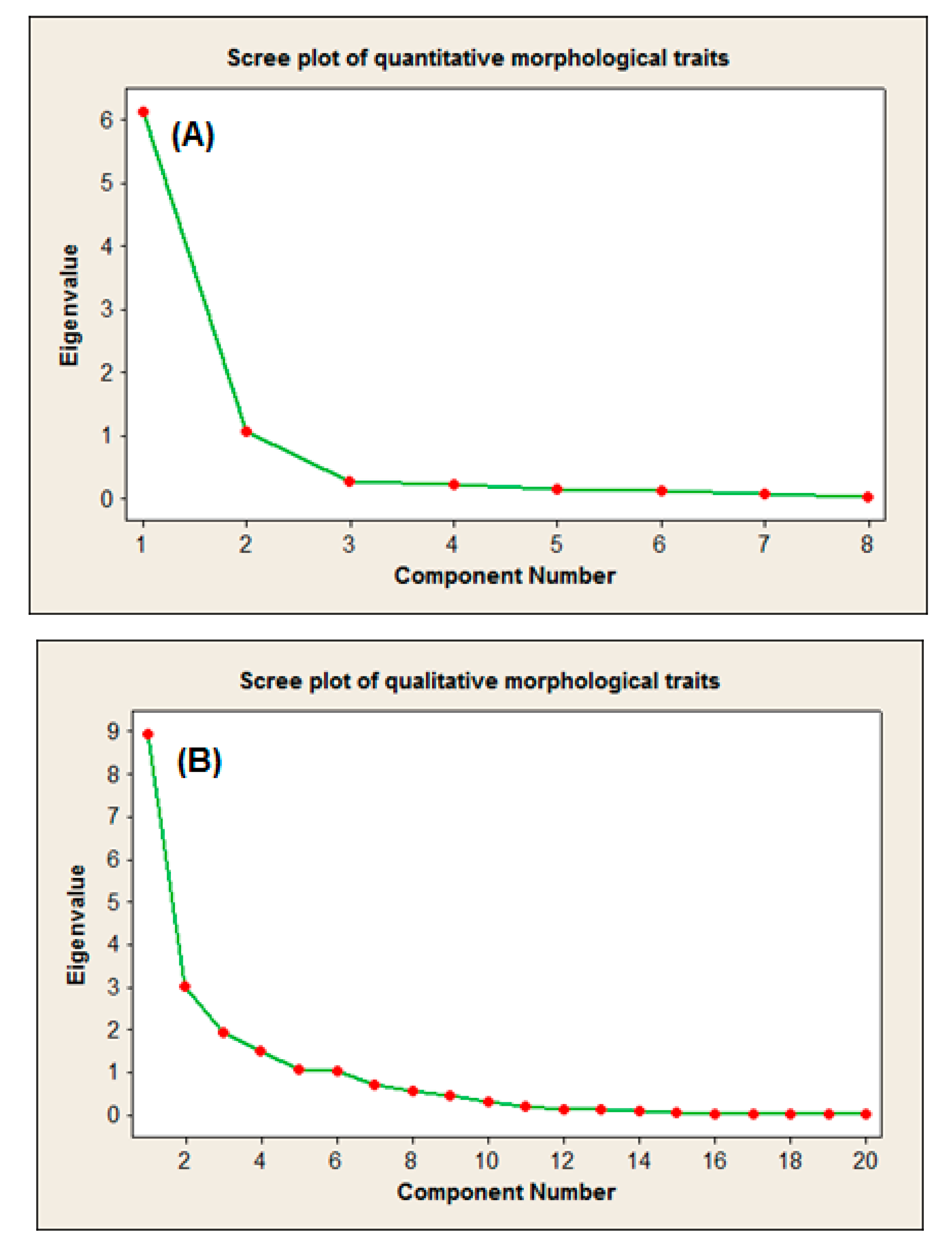
2.2.1. Scree Plot-Based Representation of the PCA
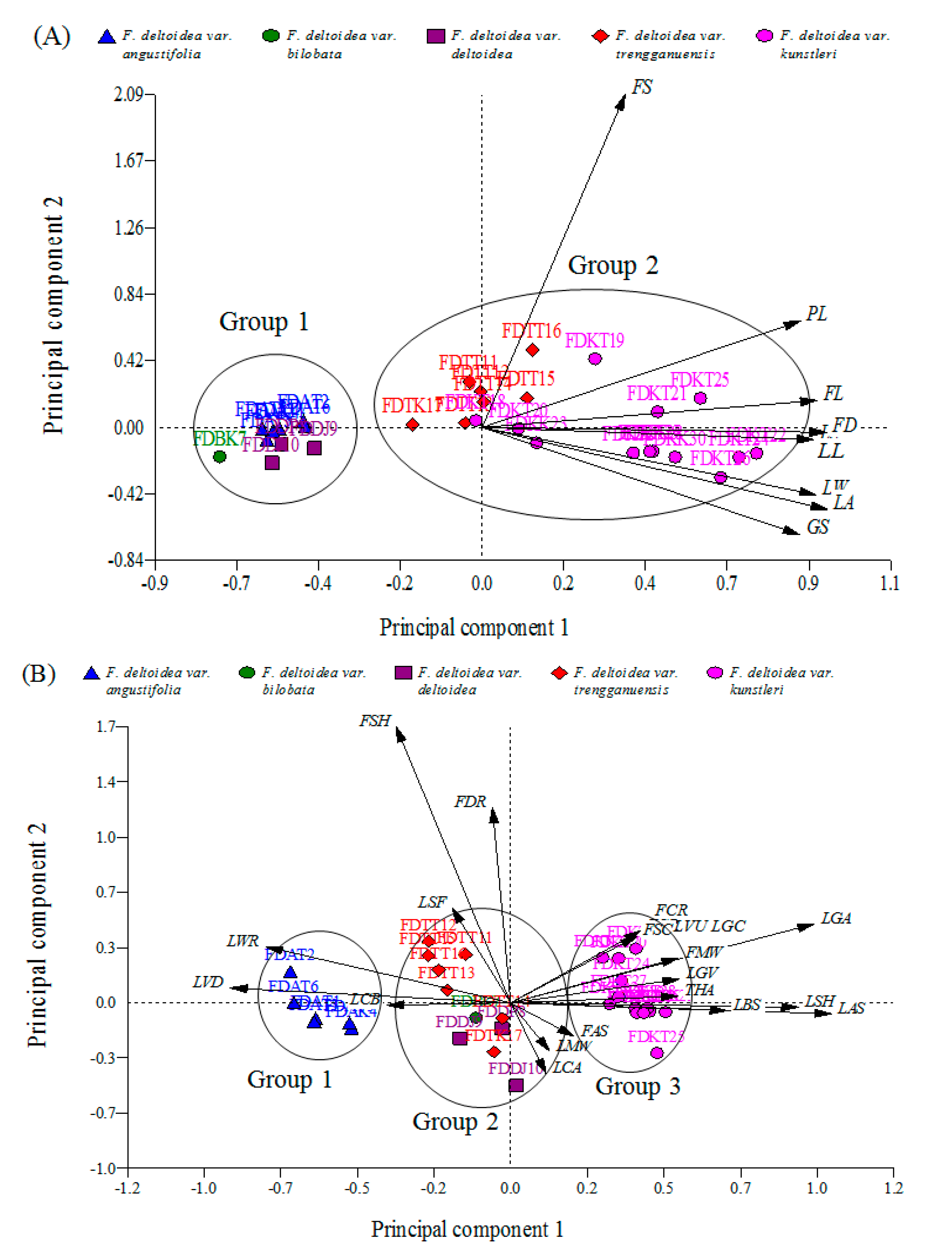
2.2.2. Biplot-Based Representation of the PCA

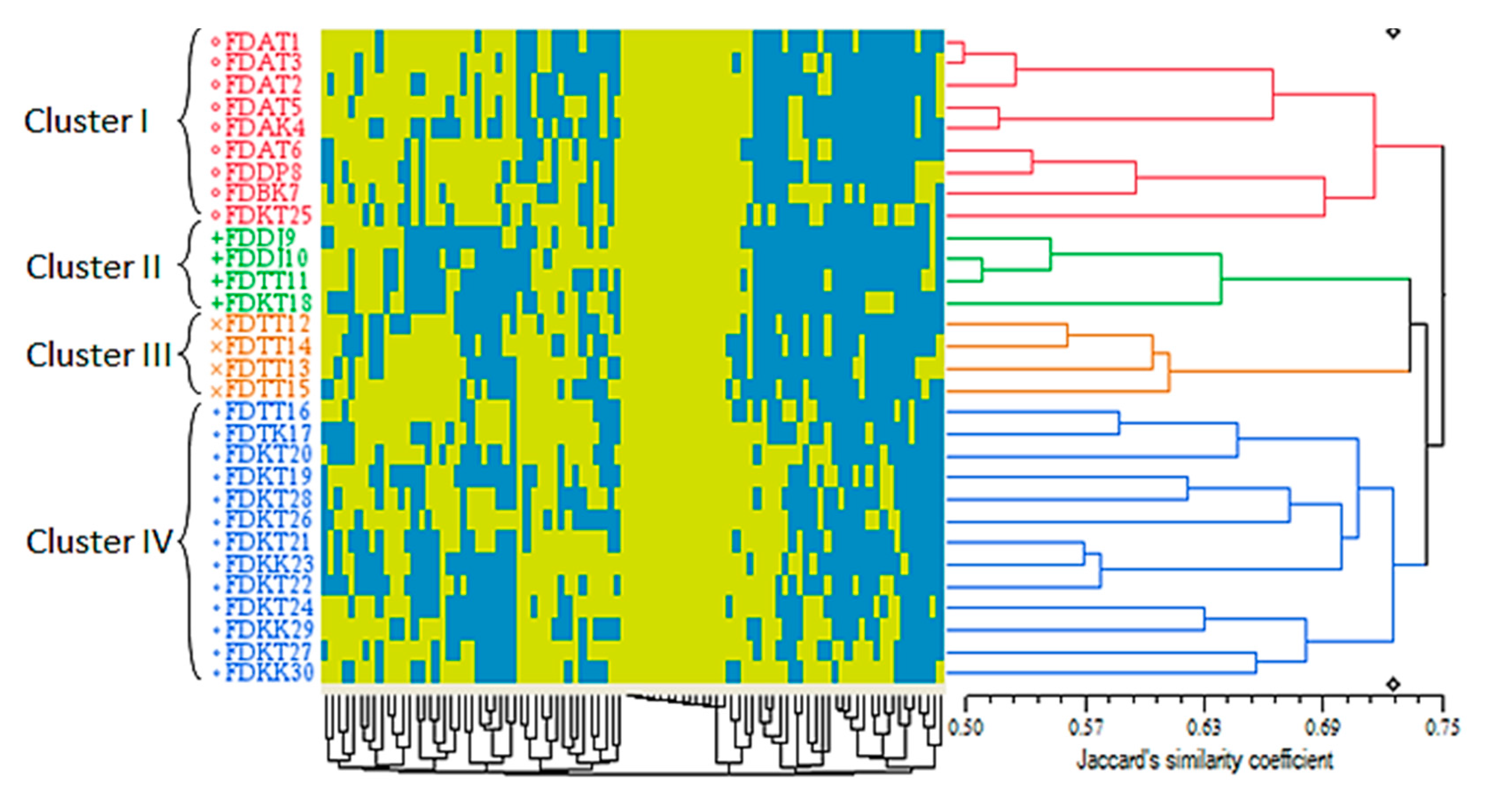
2.3. Morphological and Molecular Variation of the F. deltoidea Varieties Based on the Cluster Analysis
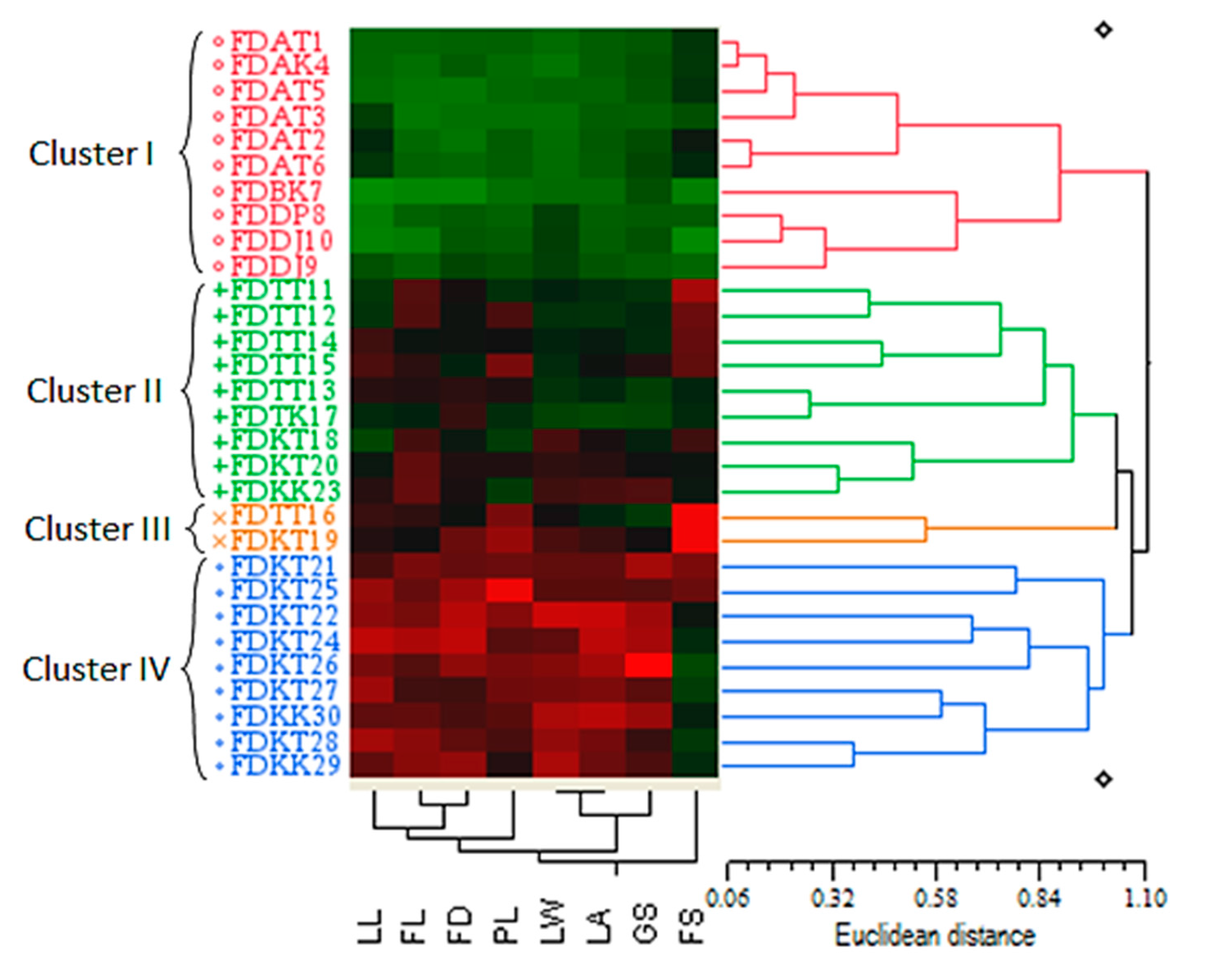
2.3.1. Cluster Analysis Based on the Quantitative Morphological Markers

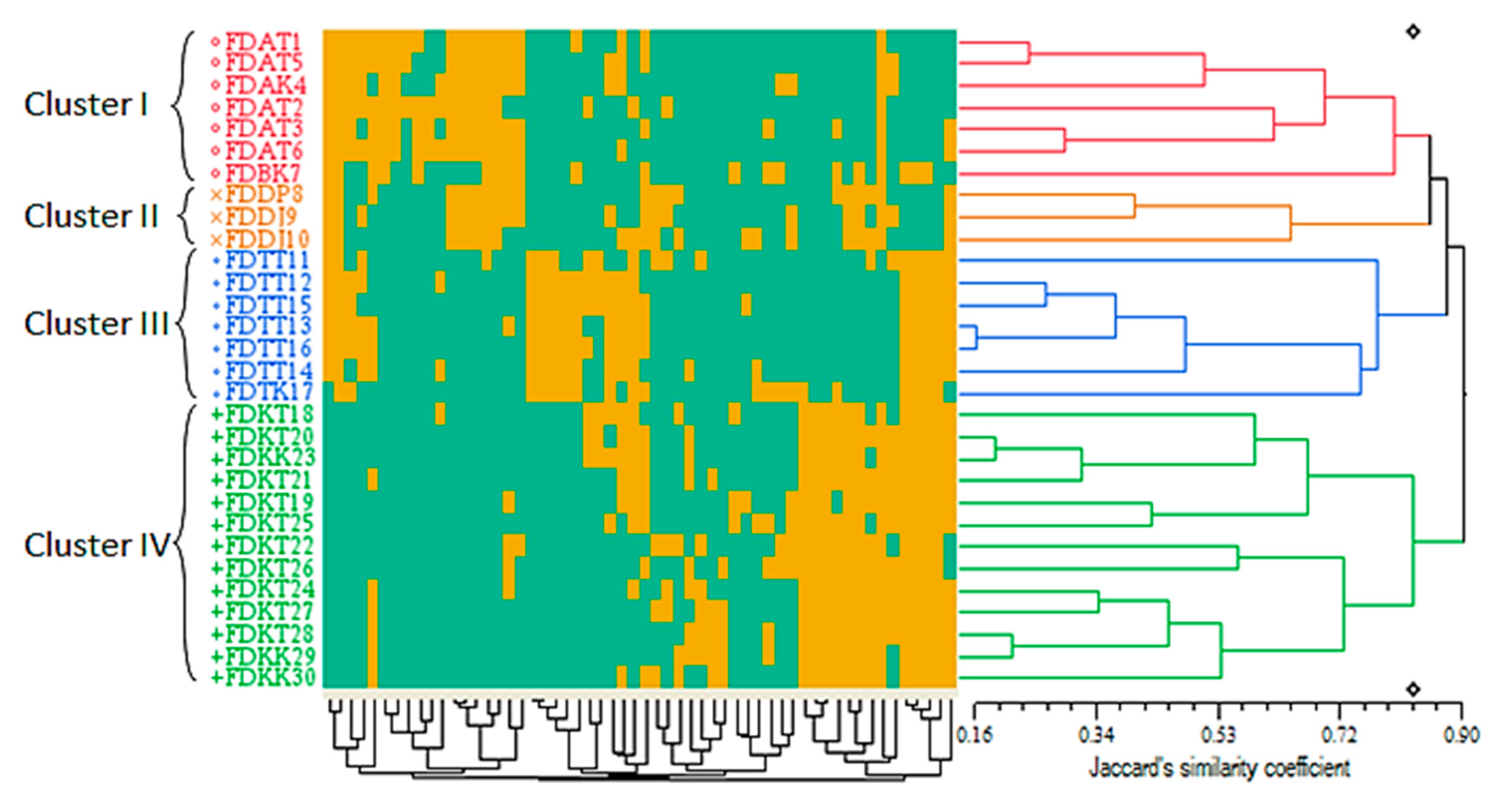
2.3.2. Cluster Analysis Based on the Qualitative Morphological Markers

2.3.3. Cluster Analysis Based on the ISSR Markers

2.3.4. Correlation of Similarity Matrices between Morphology and ISSR Marker Systems
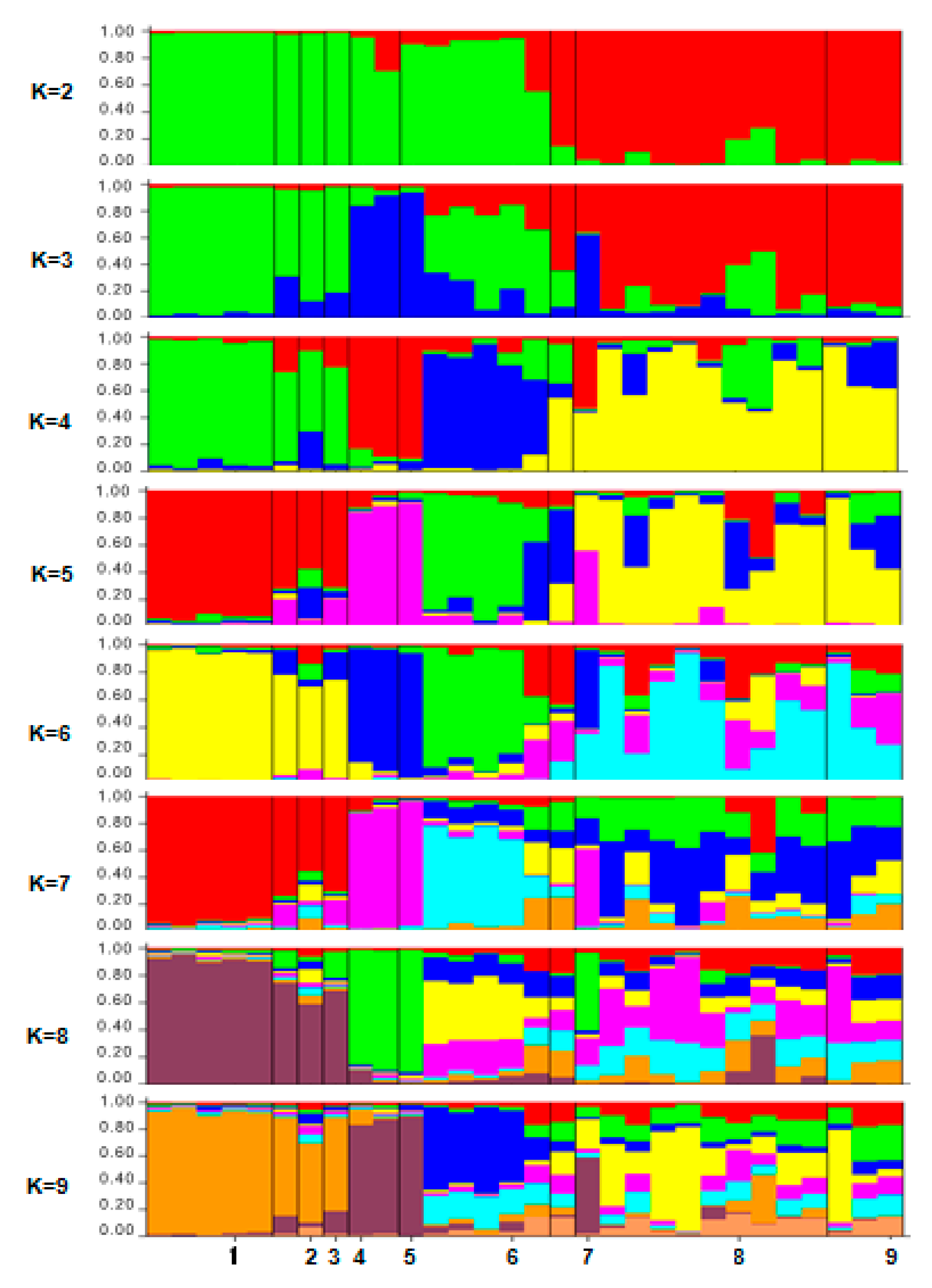
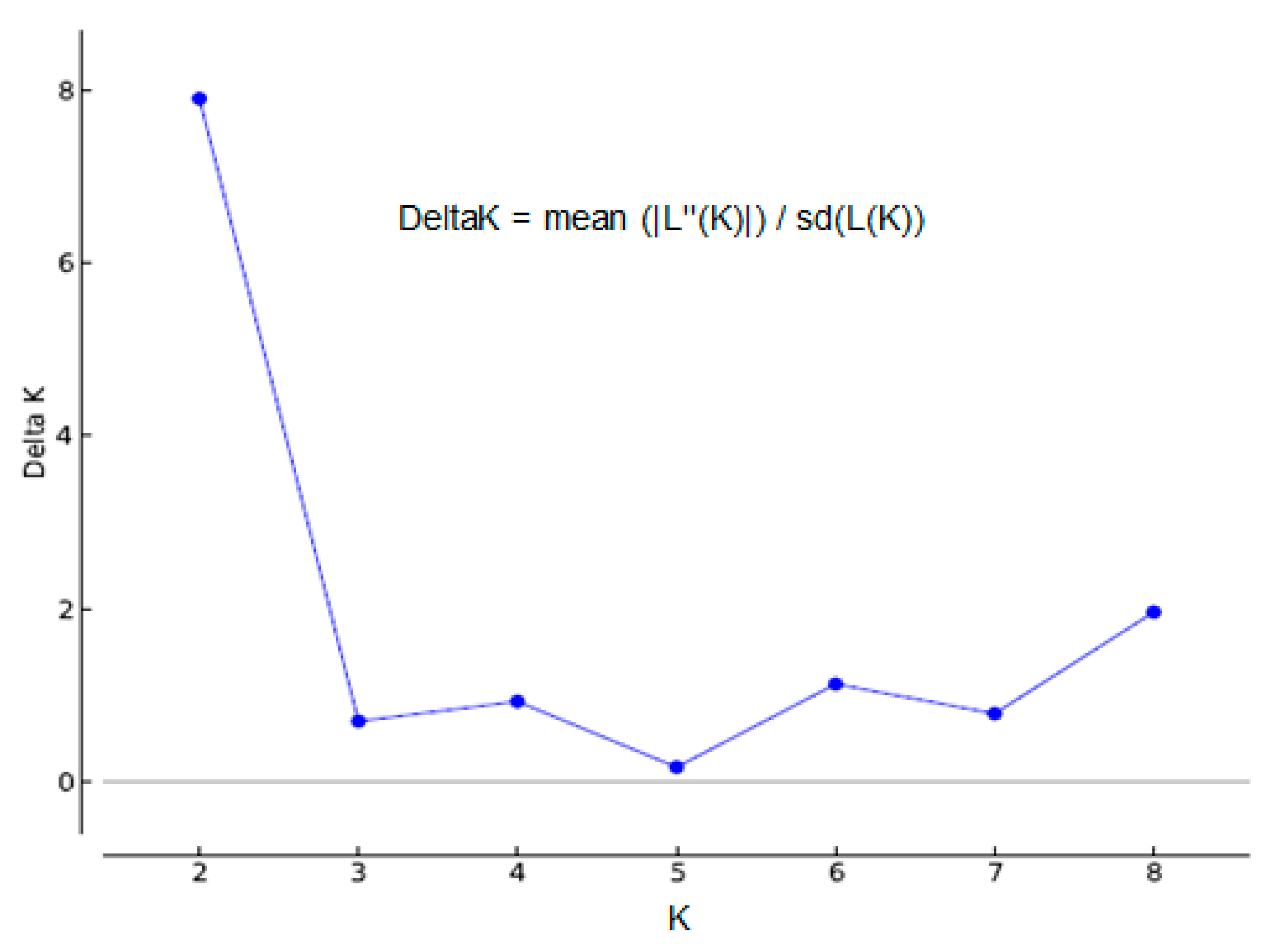
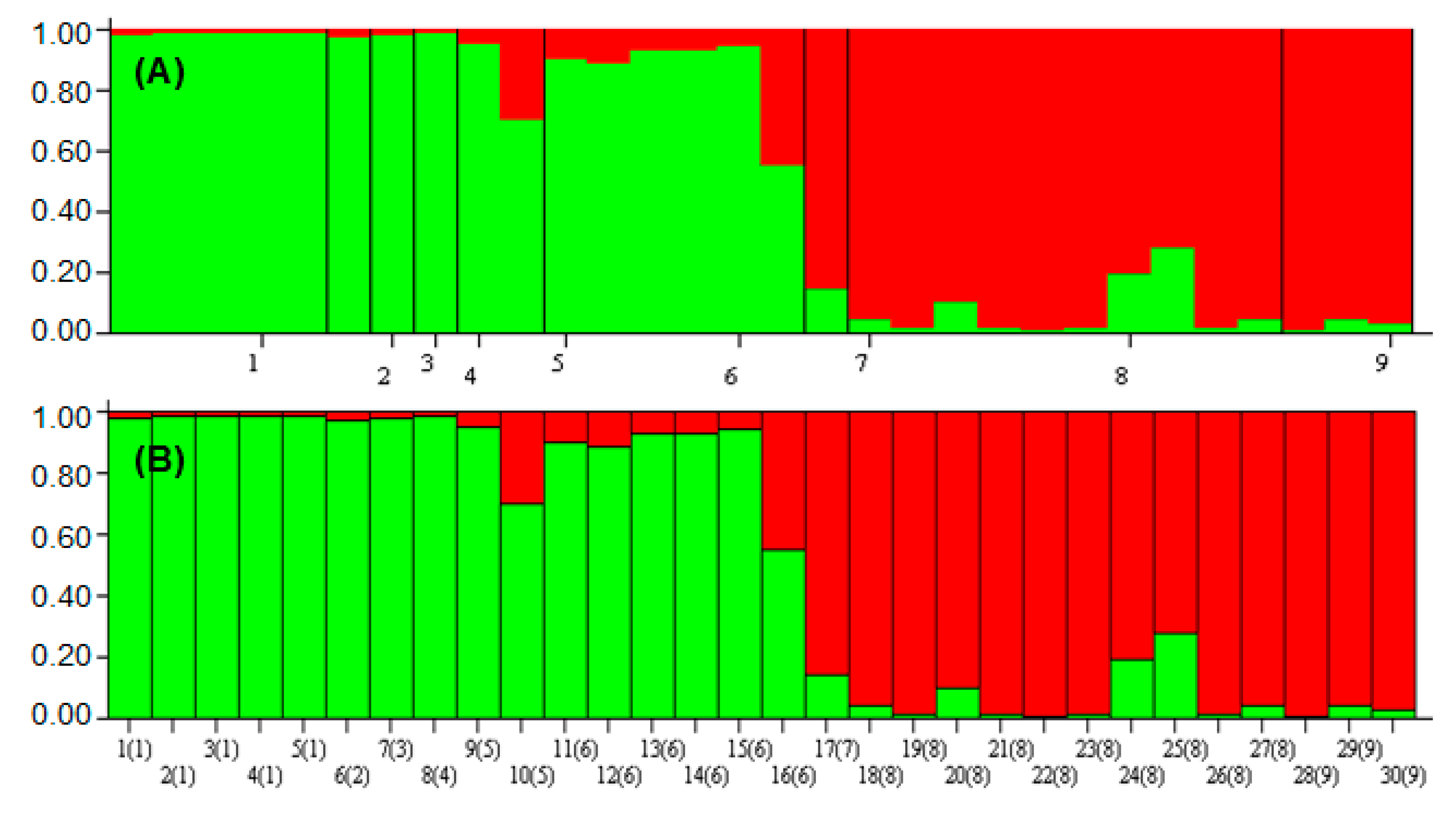
2.4. ISSR-PCR and Population Structure
| Primer | Sequence | Tm (°C) | Size Range (bp) | NAL * | NPL ** | Shannon Index | Polymorphism (%) |
|---|---|---|---|---|---|---|---|
| ISSR16 | (CAC)3GC | 38.0 | 174–1551 | 14 | 10 | 0.361 ± 0.288 | 71.4 |
| I841 | (GA)8CC | 50.3 | 420–1926 | 14 | 12 | 0.445 ± 0.243 | 85.7 |
| ISSR25 | (AC)8GA | 48.0 | 155–1524 | 11 | 10 | 0.559 ± 0.202 | 90.9 |
| UBC815 | (CT)8G | 47.0 | 218–1155 | 11 | 9 | 0.411 ± 0.254 | 81.8 |
| UBC816 | (CA)8T | 50.0 | 158–1341 | 14 | 11 | 0.390 ± 0.277 | 78.5 |
| UBC823 | (TC)8C | 48.0 | 173–1239 | 15 | 13 | 0.473 ± 0.233 | 86.7 |
| UBC806 | (TA)8G | 27.8 | 166–1564 | 10 | 9 | 0.504 ± 0.212 | 90.0 |
| ISSR20 | (AC)7TA | 45.8 | 700–1792 | 6 | 0 | – | 0 |
| ISSR30 | (AC)8 | 43.4 | 385–2544 | 11 | 0 | – | 0 |
| 10 | – | – | 155–2544 | 106/15.14 | 74/8.2 | 0.449 ± 0.247 | 65.0 |



2.5. Evolutionary Hypothesis and Mating System of F. deltoidea
3. Experimental Section
3.1. Plant Materials

3.2. Ethics Statement
| Population Code * | Accession | Botanical Name | Origin | Latitude | Longitude |
|---|---|---|---|---|---|
| 1 | FDAT1 | F. deltoidea var. angustifolia | Terengganu | 5° 39.086 N | 102° 84.198 E |
| 1 | FDAT2 | F. deltoidea var. angustifolia | Terengganu | 5° 60.775 N | 102° 74.873 E |
| 1 | FDAT3 | F. deltoidea var. angustifolia | Terengganu | 5° 39.086 N | 102° 84.198 E |
| 2 | FDAK4 | F. deltoidea var. angustifolia | Kelantan | 5° 97.854 N | 102° 42.650 E |
| 1 | FDAT5 | F. deltoidea var. angustifolia | Terengganu | 5° 60.775 N | 102° 74.873 E |
| 1 | FDAT6 | F. deltoidea var. angustifolia | Terengganu | 5° 52.281 N | 102° 93.927 E |
| 3 | FDBK7 | F. deltoidea var. bilobata | Kelantan | 5° 97.854 N | 102° 42.650 E |
| 4 | FDDP8 | F. deltoidea var. deltoidea | Pahang | 2° 90.220 N | 102° 89.700 E |
| 5 | FDDJ9 | F. deltoidea var. deltoidea | Johor | 1° 89.186 N | 102° 76.717 E |
| 5 | FDDJ10 | F. deltoidea var. deltoidea | Johor | 1° 93.317 N | 103° 18.551 E |
| 6 | FDTT11 | F. deltoidea var. trengganuensis | Terengganu | 5° 39.086 N | 102° 84.198 E |
| 6 | FDTT12 | F. deltoidea var. trengganuensis | Terengganu | 5° 39.086 N | 102° 84.198 E |
| 6 | FDTT13 | F. deltoidea var. trengganuensis | Terengganu | 5° 39.086 N | 102° 84.198 E |
| 6 | FDTT14 | F. deltoidea var. trengganuensis | Terengganu | 5° 39.086 N | 102° 84.198 E |
| 6 | FDTT15 | F. deltoidea var. trengganuensis | Terengganu | 5° 52.281 N | 102° 93.927 E |
| 6 | FDTT16 | F. deltoidea var. trengganuensis | Terengganu | 5° 60.775 N | 102° 74.873 E |
| 7 | FDTK17 | F. deltoidea var. trengganuensis | Kelantan | 5° 97.854 N | 102° 42.650 E |
| 8 | FDKT18 | F. deltoidea var. kunstleri | Terengganu | 5° 39.086 N | 102° 84.198 E |
| 8 | FDKT19 | F. deltoidea var. kunstleri | Terengganu | 5° 52.281 N | 102° 93.927 E |
| 8 | FDKT20 | F. deltoidea var. kunstleri | Terengganu | 5° 39.086 N | 102° 84.198 E |
| 8 | FDKT21 | F. deltoidea var. kunstleri | Terengganu | 5° 60.775 N | 102° 74.873 E |
| 8 | FDKT22 | F. deltoidea var. kunstleri | Terengganu | 5° 39.086 N | 102° 84.198 E |
| 9 | FDKK23 | F. deltoidea var. kunstleri | Kelantan | 5° 97.854 N | 102° 42.650 E |
| 8 | FDKT24 | F. deltoidea var. kunstleri | Terengganu | 5° 39.086 N | 102° 84.198 E |
| 8 | FDKT25 | F. deltoidea var. kunstleri | Terengganu | 5° 39.086 N | 102° 84.198 E |
| 8 | FDKT26 | F. deltoidea var. kunstleri | Terengganu | 5° 39.086 N | 102° 84.198 E |
| 8 | FDKT27 | F. deltoidea var. kunstleri | Terengganu | 5° 60.775 N | 102° 74.873 E |
| 8 | FDKT28 | F. deltoidea var. kunstleri | Terengganu | 5° 39.086 N | 102° 84.198 E |
| 9 | FDKK29 | F. deltoidea var. kunstleri | Kelantan | 5° 97.854 N | 102° 42.650 E |
| 9 | FDKK30 | F. deltoidea var. kunstleri | Kelantan | 5° 97.854 N | 102° 42.650 E |
3.3. Morphological Characteristics
3.3.1. Quantitative Data Collection
3.3.2. Qualitative Data Collection
3.4. Molecular Characterization and Data Collection
| Characters | Abbreviation | Coding Description |
|---|---|---|
| Tree habit | THA | 1, erect; 2, leaning |
| Leaf shape * | LSH | 1, spatulate; 2, elliptic/oval; 3, obovate |
| Shape of leaf apex | LAS | 1, obtuse; 2, rounded; 3, truncate; 4, obcordate |
| Shape of the leaf base | LBS | 1, attenuate; 2, narrowly cuneate; 3, cuneate |
| Type of the leaf surface | LSF | 1, flat; 2, vein impressed; 3, margin dented |
| Maximum width of the leaf | LMW | 1, 1/3 to 2/3 from leaf base; 2, more than 2/3 from leaf base |
| Leaf length: width ratio * | LWR | 1, 1:1; 2, 1.5:1; 3, 2:1; 4, 3:1 |
| Color of the upper-leaf surface | LCA | 1, light green; 2, green; 3, dark green |
| Color of the underside-leaf surface | LCB | 1, yellow; 2, greenish yellow; 3, green |
| Gland color | LGC | 1, red; 2, purple or black |
| Amount of gland on leaf * | LAG | 1, one; 2, three to five; 3, more than five |
| Visible Glands on the upper-leaf surface | LGV | 1, non-visible; 2, visible |
| Leaf venation of the upper-leaf surface | LVU | 1, Unapparent; 2, Apparent |
| Vein began to diverge * | LVD | 1, less than 1/3 from leaf base; 2, 1/3 to less than 2/3 from leaf base; 3, 2/3 and more from leaf base |
| Leaf length | LL | Measured in cm |
| Leaf width | LW | Measured in cm |
| Leaf area | LA | Measured in cm2 |
| Petiole length | PL | Measured in cm |
| Amount of golden spots on leaf surface | GS | Number of golden spots |
| Fruit shape | FSH | 1, globose; 2, subglobose; 3, bell shape; 4, ovoid |
| Maximum width of fruit | FMW | 1, less than 1/3 from leaf base; 2, 1/3 to less than 2/3 from leaf base; 3, 2/3 and more from leaf base |
| Fruit apex shape | FAS | 1, acute; 2, round; 3, truncate; 4, cordate |
| Ratio of fruit length diameter | FDR | 1, 1:1; 2, 1.5:1; 3, 2:1 |
| Color of ripening fruit | FCR | 1, green to orange; 2, green to pink or red |
| Scale color | FSC | 1, orange; 2, red/brown |
| Fruit length | FL | Measured in cm |
| Fruit diameter * | FD | Measured in cm |
| Fruit stalk length | FS | Measured in cm |
3.4.1. DNA Extraction, Polymerase Chain Reaction (PCR) Protocols
3.4.2. Electrophoresis and Gel Visualization
3.5. Experimental Design and Statistical Analysis
3.5.1. PCA and Cluster Analysis Based on the Morphological Data
3.5.2. Cluster Analysis Based on the Molecular Data
3.5.3. STRUCTURE Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Kamarudin, K.R.; Rehan, A.M.; Hashim, R.; Usup, G. An update on diversity of sea cucumbers (Echinodermata: Holothuroidea) in Malaysia. Malays. Nat. J. 2010, 62, 315–334. [Google Scholar]
- Hezri, A.A.; Hasan, M.N. Towards sustainable development? The evolution of environmental policy in Malaysia. Nat. Resour. Forum 2006, 30, 37–50. [Google Scholar] [CrossRef]
- Kochummen, K.M.; Rusea, G. Moraceae. Tree Flora Sabah Sarawak 2000, 3, 181–334. [Google Scholar]
- Bunawan, H.; Amin, N.M.; Bunawan, S.N.; Baharum, S.N.; Noor, N.M. Ficus Deltoidea Jack: A review on its phytochemical and pharmacological importance. Evid. Based Complement. Altern. Med. 2014, 2014, 902734. [Google Scholar] [CrossRef] [PubMed]
- Draman, S.; Aris, M.A.; Akter, S.F.U.; Azlina, H.; Nor, A.; Muzaffar, R.; Norazlanshah, H. Mas Cotek (Ficus deltoidea): A Possible supplement for type II diabetes: A pilot study. Pertanika J. Trop. Agric. Sci. 2012, 35, 93–102. [Google Scholar]
- Adam, Z.; Hamid, M.; Ismail, A.; Khamis, S. Effect of Ficus deltoidea aqueous extract on blood glucose level in normal and mild diabetic rats. Malays. J. Health Sci. 2007, 5, 9–16. [Google Scholar]
- Aminudin, N.; Sin, C.Y.; Chee, E.S.; Nee, K.I.; Renxin, L. Blood glucose lowering effect of Ficus deltoidea aqueous extract. Malays. J. Sci. 2007, 26, 73–78. [Google Scholar]
- Hakiman, M.; Maziah, M. Non-enzymatic and enzymatic antioxidant activities in aqueous extract of Ficus deltoidea accessions. J. Med. Plant Res. 2009, 3, 120–131. [Google Scholar]
- Misbah, H.; Azlina, A.A.; Aminudin, N. Antidiabetic and antioxidant properties of Ficus deltoidea fruit extracts and fractions. BMC Complement. Altern. Med. 2013, 13, 118. [Google Scholar] [CrossRef] [PubMed]
- Sulaiman, M.R.; Hussain, M.K.; Zakaria, Z.A.; Somchit, M.N.; Moin, S.; Mohamad, A.S.; Israf, D.A. Evaluation of the antinociceptive activity of Ficus deltoidea aqueous extract. Fitoterapia 2008, 79, 557–561. [Google Scholar] [CrossRef] [PubMed]
- Abdulla, M.A.; Ahmed, K.A.A.; Abu-Luhoom, F.M.; Muhanid, M. Role of Ficus deltoidea extract in the enhancement of wound healing in experimental rats. Biomed. Res. 2010, 21, 241–245. [Google Scholar]
- Berg, C.C. Flora Malesiana precursor for the treatment of Moraceae 3: Ficus subgenus Ficus. Blumea 2003, 48, 529–550. [Google Scholar] [CrossRef]
- Fatihah, H.N.N.; Mat, N.; Zaimah, A.R.; Zuhailah, M.N.; Norhaslinda, H. Morphological phylogenetic analysis of seven varieties of Ficus deltoidea Jack from the Malay Peninsula of Malaysia. PLoS ONE 2012, 7, e52441. [Google Scholar] [CrossRef] [PubMed]
- Berg, C.C.; Corner, E.J. Moraceae (Ficus). Ser. I Seed Plants Flora Malays. 2005, 17, 1–729. [Google Scholar]
- Mat, N.; Rosni, N.A.; Ab Rashid, N.Z.; Haron, N.; Nor, Z.M.; Nudin, N.F.H.; Yunus, A.G.; Ali, A.M. Leaf morphological variations and heterophylly in Ficus deltoidea Jack (Moraceae). Sains Malays. 2012, 41, 527–538. [Google Scholar]
- Valdiani, A.; Kadir, M.A.; Saad, M.S.; Talei, D.; Tan, S.G. Intra-specific hybridization: Generator of genetic diversification and heterosis in Andrographis paniculata Nees. A bridge from extinction to survival. Gene 2012, 505, 23–36. [Google Scholar] [CrossRef] [PubMed]
- Dehghan-Shoar, M.; Hampton, J.G.; Hill, M.J. Identifying and discriminating among Lucerne cultivars using plant morphological characters. N. Z. J. Agric. Res. 2005, 48, 271–276. [Google Scholar] [CrossRef]
- Bhore, S.J.; Nurul, A.H.; Shah, F.H. Genetic variability based on randomly amplified polymorphic DNA in mistletoe fig (Ficus deltoidea Jack) collected from Peninsular Malaysia. J. For. Sci. 2009, 25, 57–65. [Google Scholar]
- Foll, M.; Gaggiotti, O. Identifying the environmental factors that determine the genetic structure of populations. Genetics 2006, 174, 875–891. [Google Scholar] [CrossRef] [PubMed]
- Valdiani, A.; Talei, D.; Javanmard, A.; Tan, S.G.; Kadir, M.A.; Maziah, M. Morpho-molecular analysis as a prognostic model for repulsive feedback of “Andrographis paniculata” to allogamy. Gene 2014, 542, 156–167. [Google Scholar] [CrossRef] [PubMed]
- Corner, E. The complexity of Ficus deltoidea; A recent invasion of the Sunda Shelf. Philos. Trans. R. Soc. 1969, 256, 281–317. [Google Scholar] [CrossRef]
- Went, F.W. The effect of temperature on plant growth. Annu. Rev. Plant Physiol. 1953, 4, 347–362. [Google Scholar] [CrossRef]
- Morison, J.I.L.; Morecroft, M.D. Plant Growth and Climate Change; Blackwell Publishing Ltd.: Oxford, UK, 2006. [Google Scholar]
- Talei, D.; Valdiani, A.; Khanif, Y.M.; Abdullah, M.P. Estimation of salt tolerance in Andrographis paniculata accessions using multivariate regression model. Euphytica 2013, 189, 147–160. [Google Scholar] [CrossRef]
- Iezzoni, A.F.; Pritts, M.P. Applications of principal components analysis to horticultural research. Hortic. Sci. 1991, 26, 334–338. [Google Scholar]
- Mars, M.; Marrakchi, M. Diversity of pomegranate (Punica granatum L.) germplasm in Tunisia. Genet. Resour. Crop Evol. 1999, 46, 461–467. [Google Scholar] [CrossRef]
- Upadhyaya, H.D.; Gowda, C.L.; Buhariwalla, H.K.; Crouch, J.H. Efficient use of crop germplasm resources: Identifying useful germplasm for crop improvement through core and mini-core collections and molecular marker approaches. Plant Genet. Resour. Character Util. 2006, 4, 25–35. [Google Scholar] [CrossRef]
- Goodman, M.M. Distance analysis in biology. Syst. Zool. 1972, 21, 174–186. [Google Scholar] [CrossRef]
- Schut, J.W.; Qi, X.; Stam, P. Association between relationship measures based on AFLP markers, pedigree data and morphological traits in barley. Theor. Appl. Genet. 1997, 95, 1161–1168. [Google Scholar] [CrossRef]
- Papadopoulou, K.; Ehaliotis, C.; Tourna, M.; Kastanis, P.; Karydis, I.; Zervakis, G. Genetic relatedness among dioecious Ficus carica L. cultivars by random amplified polymorphic DNA analysis, and evaluation of agronomic and morphological characters. Genetica 2002, 114, 183–194. [Google Scholar] [CrossRef] [PubMed]
- Saddoud, O.; Baraket, G.; Chatti, K.; Trifi, M.; Marrakchi, M.; Salhi-Hannachi, A.; Mars, M. Morphological variability of Fig (Ficus carica L.) cultivars. Int. J. Fruit Sci. 2008, 8, 35–51. [Google Scholar] [CrossRef]
- Podgornik, M.; Vuka, I.; Vrhovnik, I.; Mavsar, D. A survey and morphological evaluation of fig (Ficus carica L.) genetic resources from Slovenia. Sci. Hortic. 2010, 125, 380–389. [Google Scholar] [CrossRef]
- Hardig, T.M.; Brunsfeld, S.J.; Fritz, R.S.; Morgan, M.; Orians, C.M. Morphological and molecular evidence for hybridization and introgression in a willow (Salix) hybrid zone. Mol. Ecol. 2000, 9, 9–24. [Google Scholar] [CrossRef] [PubMed]
- Ackermann, R.R.; Bishop, J.M. Morphological and molecular evidence reveals recent hybridization between gorilla taxa. Evolution 2010, 64, 271–290. [Google Scholar] [CrossRef] [PubMed]
- Ruas, P.M.; Ruas, C.F.; Rampim, L.; Carvalho, V.P.; Ruas, E.A.; Sera, T. Genetic relationship in Coffea species and parentage determination of interspecific hybrids using ISSR (Inter-Simple Sequence Repeat) markers. Genet. Mol. Biol. 2003, 26, 319–327. [Google Scholar] [CrossRef]
- Harper, F.M.; Hart, M.W. Morphological and phylogenetic evidence for hybridization and introgression in a sea star secondary contact zone. Invertebr. Biol. 2007, 126, 373–384. [Google Scholar] [CrossRef]
- Neophytou, C. Bayesian clustering analyses for genetic assignment and study of hybridization in oaks: Effects of asymmetric phylogenies and asymmetric sampling scheme. Tree Genet. Genomes 2014, 10, 273–285. [Google Scholar] [CrossRef]
- Liu, J.; Shi, S.; Chang, E.; Yang, W.; Jiang, Z. Genetic diversity of the critically endangered thuja sutchuenensis revealed by ISSR markers and the implications for conservation. Int. J. Mol. Sci. 2013, 14, 14860–14871. [Google Scholar] [CrossRef] [PubMed]
- Jaccard, P. A comparative study of the floral distribution in Alps and Jura. Bull. Walden Soc. Nat. Sci. 1901, 37, 547–579. [Google Scholar]
- Tairo, F.; Mneney, E.; Kullaya, A. Morphological and agronomical characterization of sweet potato (Ipomoea batatas (L.) Lam.) germplasm collection from Tanzania. Afr. J. Plant Sci. 2008, 2, 77–85. [Google Scholar]
- Sokal, R.R.; Micheners, C.D. A statistical method for evaluating systematic relationships. Univ. Kans. Sci. Bull. 1958, 38, 1409–1438. [Google Scholar]
- Dalirsefat, S.B.; da Silva Meyer, A.; Mirhoseini, S.Z. Comparison of similarity coefficients used for cluster analysis with amplified fragment length polymorphism markers in the silkworm, Bombyx mori. J. Insect Sci. 2009, 9, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Mantel, N. The detection of disease clustering and generalized regression approach. Cancer Res. 1967, 27, 209–220. [Google Scholar] [PubMed]
- Gao, X.Y.; Zhi, X.Y.; Li, H.W.; Klenk, H.P.; Li, W.J. Comparative genomics of the bacterial genus streptococcus illuminates evolutionary implications of species groups. PLoS ONE 2014, 9, e101229. [Google Scholar] [CrossRef] [PubMed]
- Earl, D.A.; von Holdt, B.M. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Egger, B.; Sefc, K.M.; Makasa, L.; Sturmbauer, C.; Salzburger, W. Introgressive hybridization between color morphs in a population of cichlid fishes twelve years after human-induced secondary admixis. J. Hered. 2012, 103, 515–522. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, B.W. Host specificity of fig wasps (Agaonidae). Evolution 1970, 24, 680–691. [Google Scholar]
- Yu, H.; Compton, S.G. Moving your sons to safety: Galls containing male fig wasps expand into the centre of figs, away from enemies. PLoS ONE 2012, 7, e30833. [Google Scholar] [CrossRef] [PubMed]
- Starr, F.; Starr, K.; Loope, L. Ficus deltoidea, Mistletoe Fig (Moraceae); United States Geological Survey-Biological Resources Division Haleakala Field Station: Maui, HI, USA, 2003. [Google Scholar]
- Harrison, R.D. Figs and Fig Wasps: An Intricate Interaction. In Proceeding of the International Field Biology Course, Sarawak, Malaysia, 15 July–14 August 2004; pp. 16–17.
- McKey, D. Population biology of figs: Applieations for conservation. Experientia 1989, 45, 661–673. [Google Scholar] [CrossRef]
- Poore, M.E.D. Studies in Malaysian rain forest I. The forest on triassic sediments in Jengka Jorest Reserve. J. Ecol. 1986, 56, 143–196. [Google Scholar] [CrossRef]
- Ranc, N.; Muños, S.; Santoni, S.; Causse, M. A clarified position for Solanum lycopersicum var. cerasiforme in the evolutionary history of tomatoes (solanaceae). BMC Plant Biol. 2008, 8, 130. [Google Scholar] [CrossRef] [PubMed]
- IPGRI; CIHEAM. Descriptors for Fig; International Plant Genetic Resources Institute, Rome Italy, and International Centre for Advanced Mediterranean Agronomic Studies; IPGRI: Paris, France, 2003. [Google Scholar]
- Pandey, S.K.; Singh, H. A simple, cost-effective method for leaf area estimation. J. Bot. 2011, 2011, 1–16. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L. Isolation of plant DNA from fresh tissue. Focus 1990, 12, 13–15. [Google Scholar]
- Rohlf, F.J. NTSYSpc: Numerical Taxonomy System. Ver. 2.1. Applied Biostatic, Exeter Software; Setauket: New York, NY, USA, 2000. [Google Scholar]
- SAS Institute Inc. JMP® 8 User Guide, 2nd ed.; SAS Institute Inc.: Cary, NC, USA, 2009. [Google Scholar]
- Hubisz, M.J.; Falush, D.; Stephens, M.; Pritchard, J.K. Inferring weak population structure with the assistance of sample group information. Mol. Ecol. Resour. 2009, 9, 1322–1332. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [PubMed]
- Falush, D.; Stephens, M.; Pritchard, J.K. Inference of population structure using multilocus genotype data: Dominant markers and null alleles. Mol. Ecol. Notes 2007, 7, 574–578. [Google Scholar] [CrossRef] [PubMed]
- Tacuatiá, L.O.; Eggers, L.; Kaltchuk-Santos, E.; Souza-Chies, T.T. Population genetic structure of Sisyrinchium micranthum Cav. (Iridaceae) in Itapuã State Park, Southern Brazil. Genet. Mol. Biol. 2012, 35, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Ge, Y.; Wang, W.; Yu, X.; Shen, X.; Liu, J.; Liu, X.; Tian, D.; Shen, F.; Yu, Y. Molecular characterization of cultivated bromeliad accessions with inter-simple sequence repeat (ISSR) markers. Int. J. Mol. Sci. 2012, 13, 6040–6052. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zimisuhara, B.; Valdiani, A.; Shaharuddin, N.A.; Qamaruzzaman, F.; Maziah, M. Structure and Principal Components Analyses Reveal an Intervarietal Fusion in Malaysian Mistletoe Fig (Ficus deltoidea Jack) Populations. Int. J. Mol. Sci. 2015, 16, 14369-14394. https://doi.org/10.3390/ijms160714369
Zimisuhara B, Valdiani A, Shaharuddin NA, Qamaruzzaman F, Maziah M. Structure and Principal Components Analyses Reveal an Intervarietal Fusion in Malaysian Mistletoe Fig (Ficus deltoidea Jack) Populations. International Journal of Molecular Sciences. 2015; 16(7):14369-14394. https://doi.org/10.3390/ijms160714369
Chicago/Turabian StyleZimisuhara, Birifdzi, Alireza Valdiani, Noor Azmi Shaharuddin, Faridah Qamaruzzaman, and Mahmood Maziah. 2015. "Structure and Principal Components Analyses Reveal an Intervarietal Fusion in Malaysian Mistletoe Fig (Ficus deltoidea Jack) Populations" International Journal of Molecular Sciences 16, no. 7: 14369-14394. https://doi.org/10.3390/ijms160714369
APA StyleZimisuhara, B., Valdiani, A., Shaharuddin, N. A., Qamaruzzaman, F., & Maziah, M. (2015). Structure and Principal Components Analyses Reveal an Intervarietal Fusion in Malaysian Mistletoe Fig (Ficus deltoidea Jack) Populations. International Journal of Molecular Sciences, 16(7), 14369-14394. https://doi.org/10.3390/ijms160714369