
Lack of Globulin Synthesis during Seed Development Alters Accumulation of Seed Storage Proteins in Rice

Abstract
:1. Introduction
2. Results
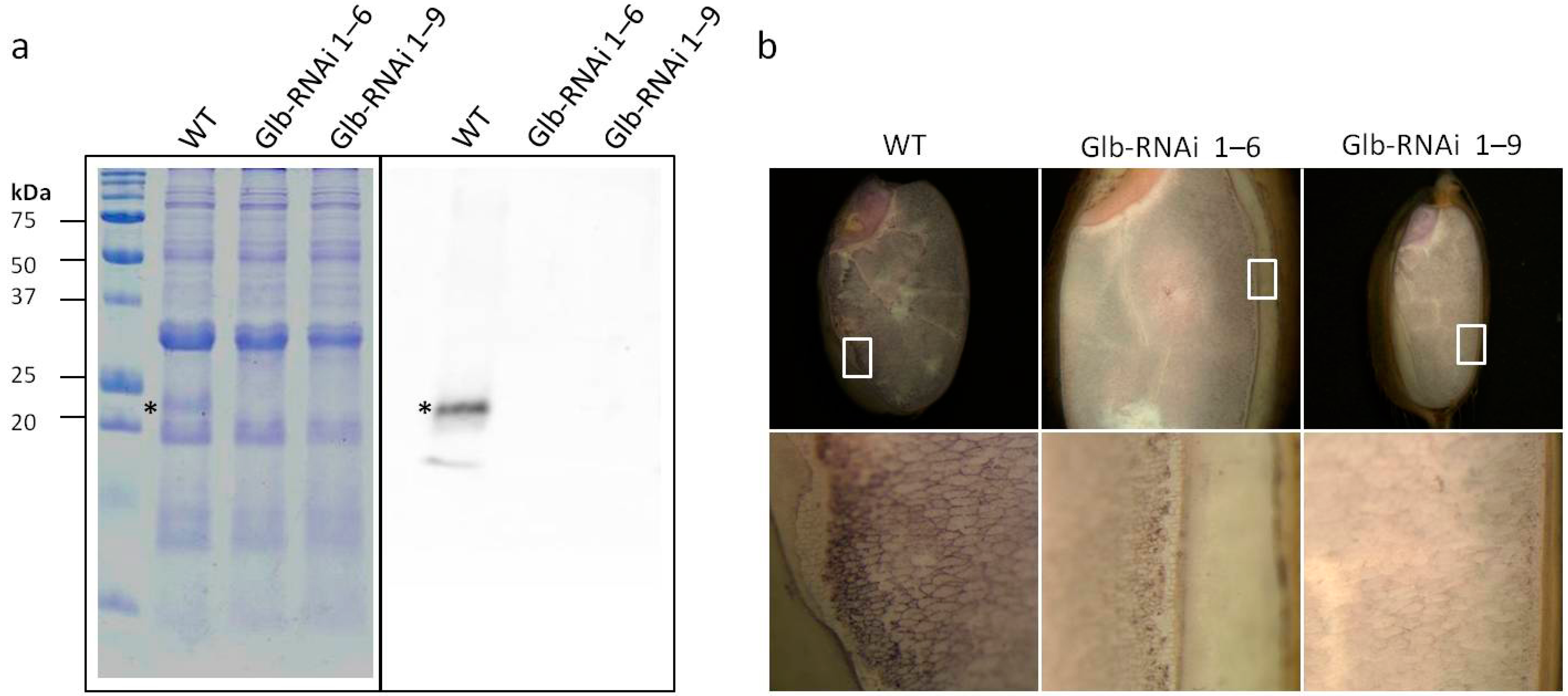
2.1. Globulin Deficiency in Glb-RNAi Seeds

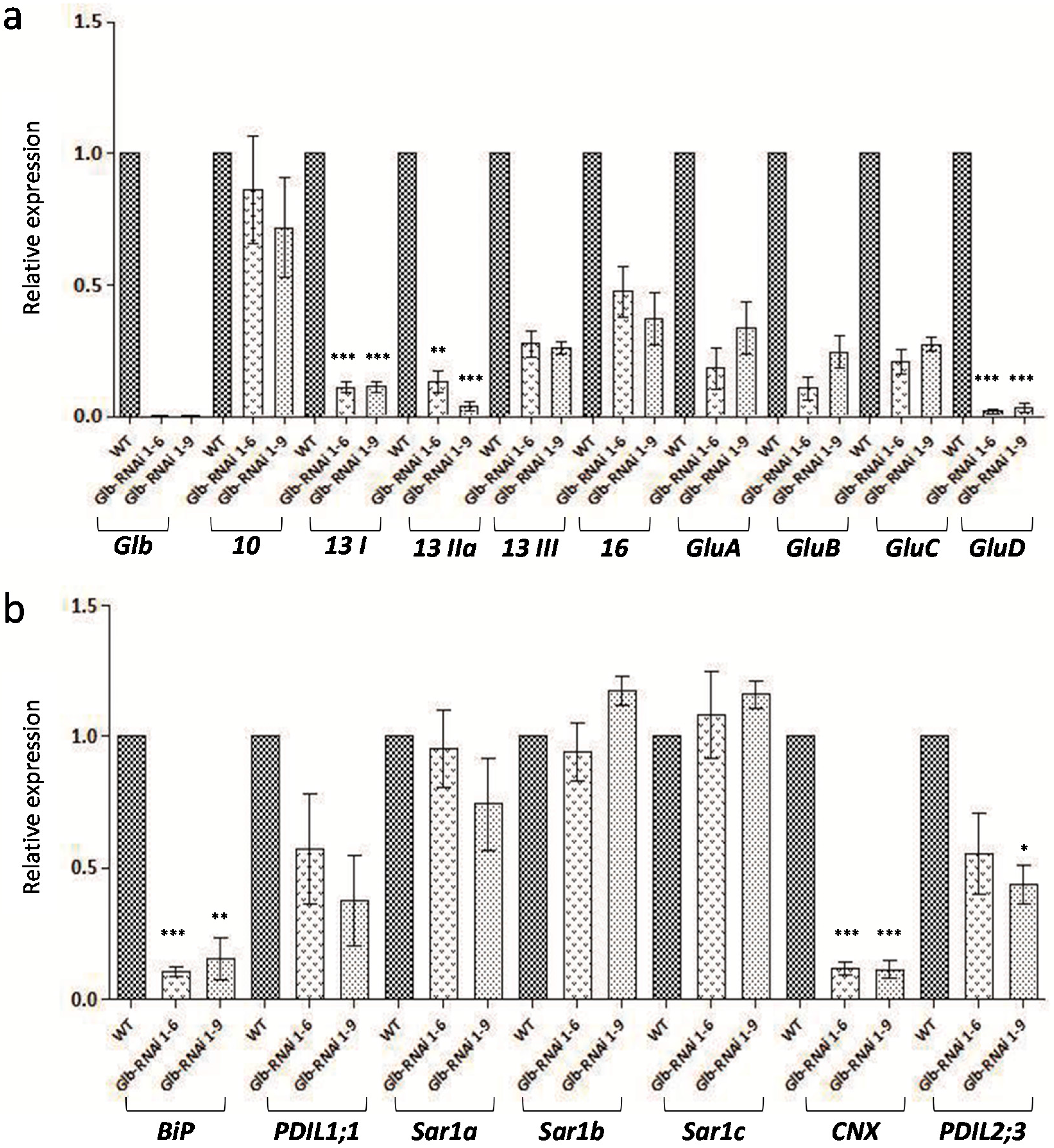
2.2. Analysis of Storage Protein Gene Expression Levels in Developing Seeds

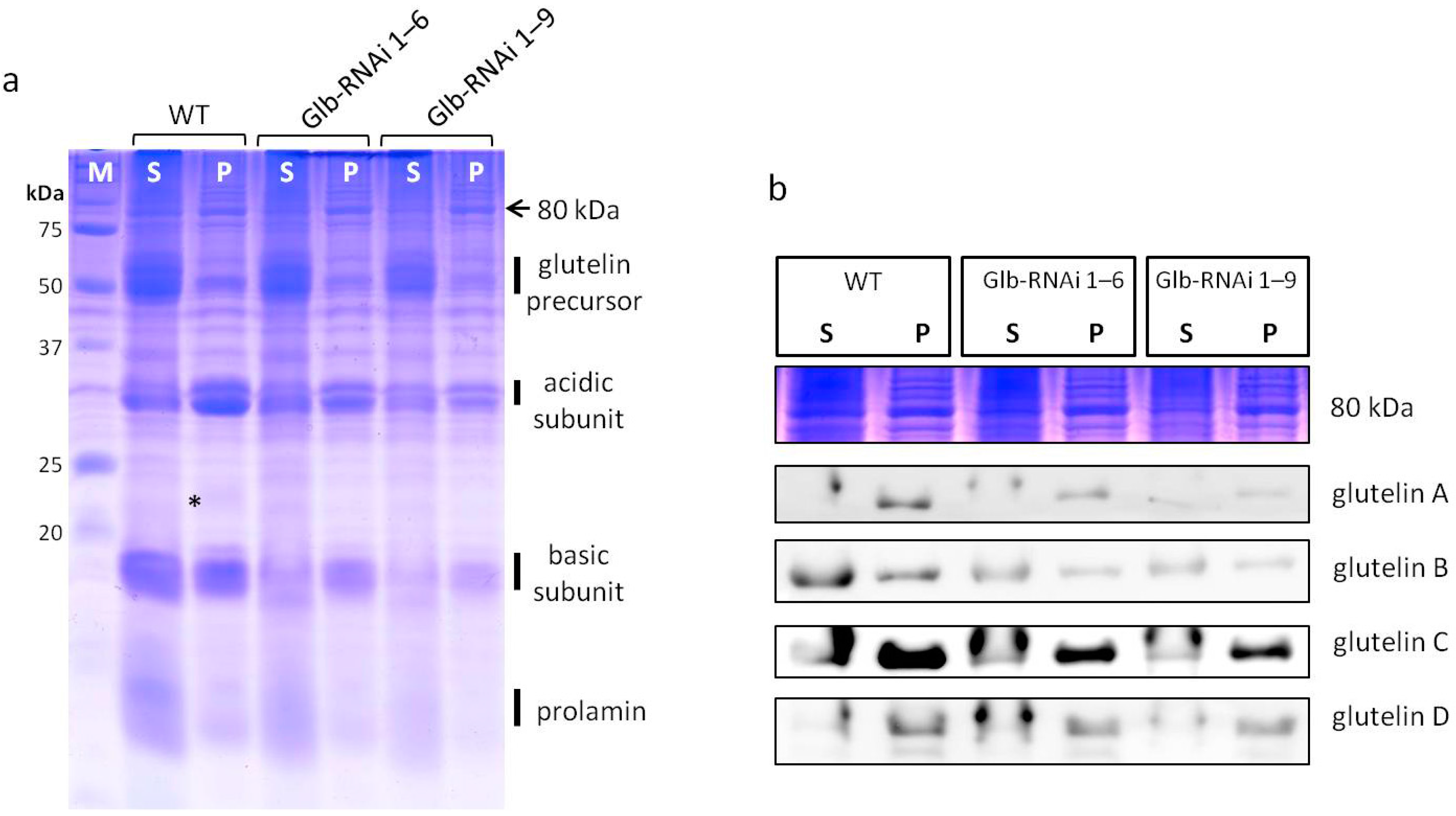
2.3. Analysis of Protein Accumulation in Mature Seeds

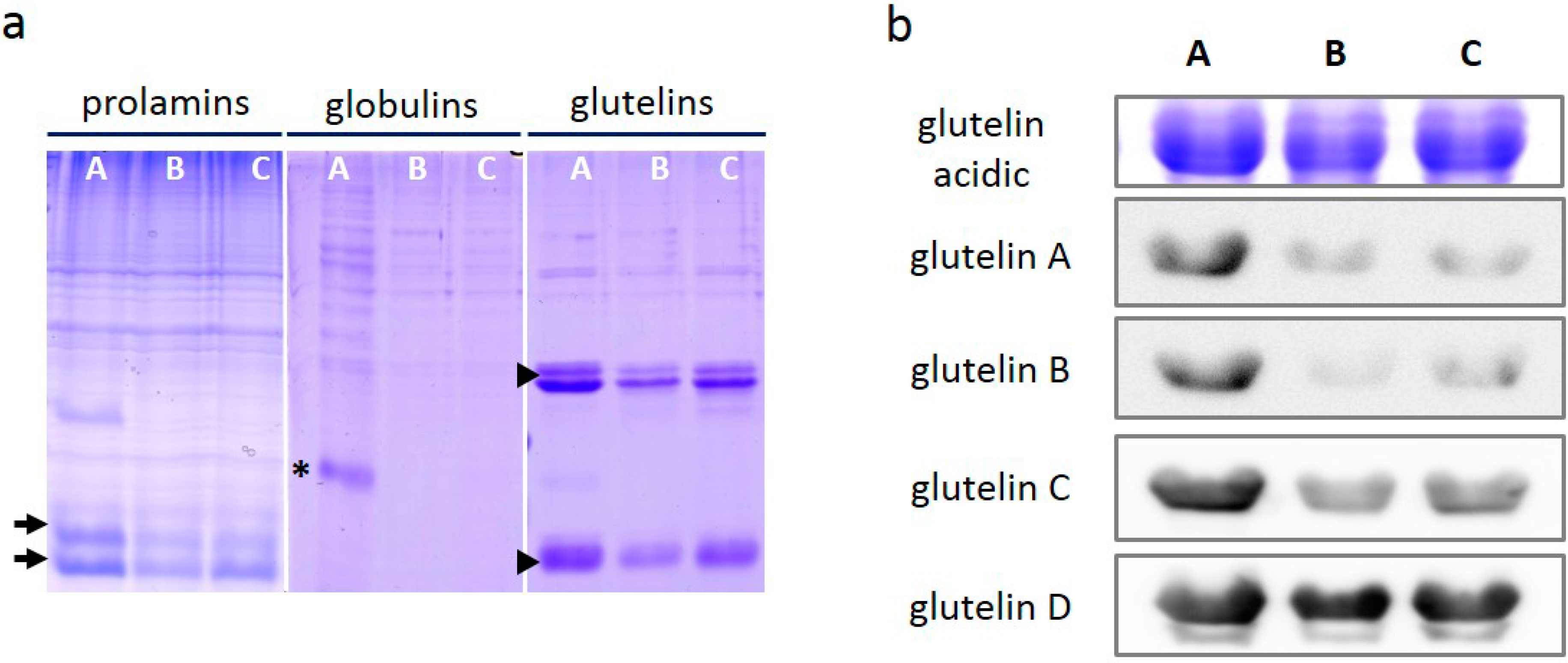
2.4. Comparative Solubility of Glutelins from Mature Seeds

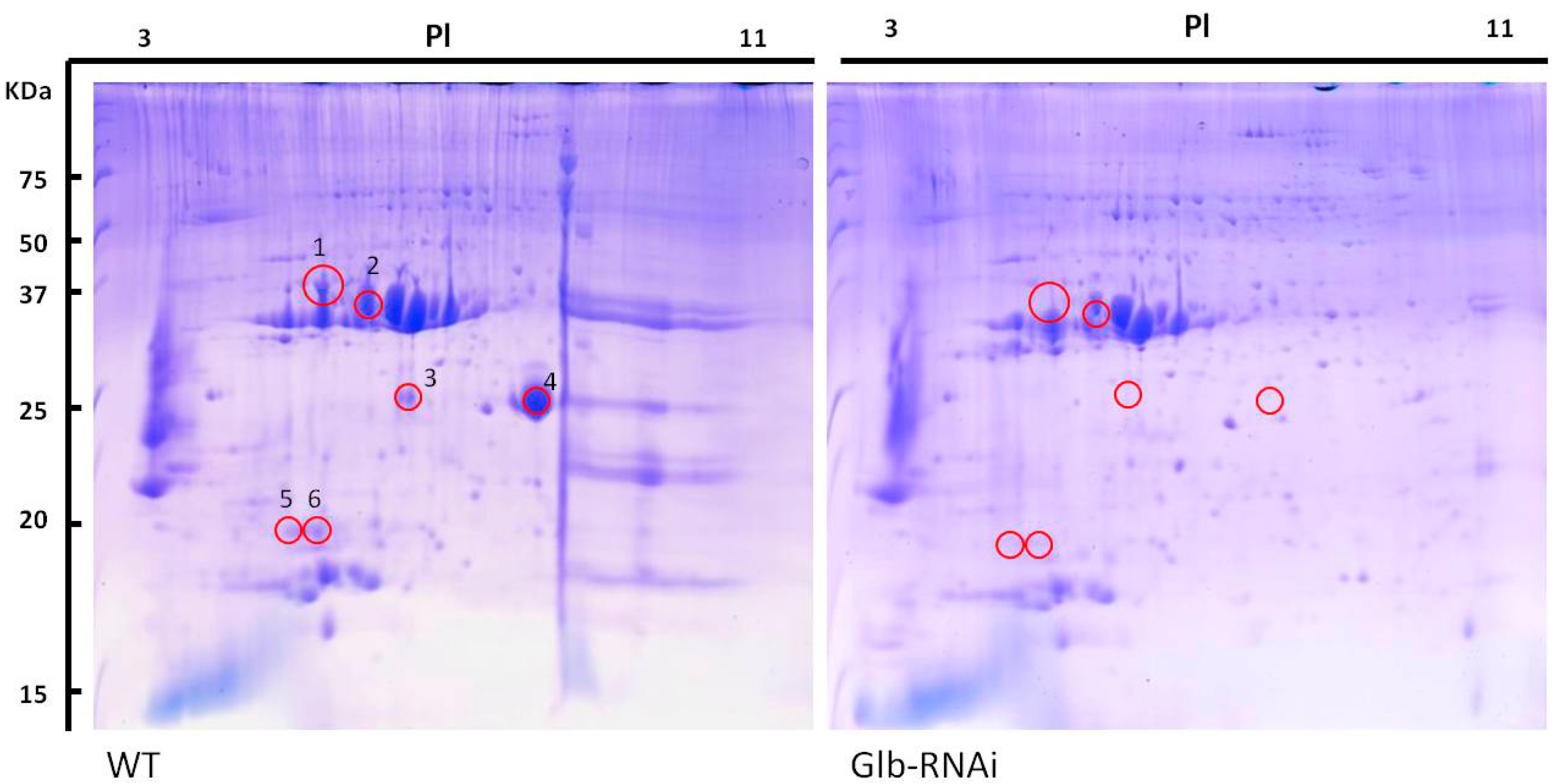
2.5. Protein Profiles in Globulin-Deficient Seeds

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spot # | Identified Protein | Coverage a | Theor. Mw (kDa) b | Theor. Pi c | Locus No. | Function |
|---|---|---|---|---|---|---|
| 1 | Glutelin type-B1 | 29.35 | 54.1 | 9.22 | Os02g0249800 | Storage protein |
| Glutelin, provisional | 21.14 | 53.5 | 9.03 | Os02g0249600 | Storage protein | |
| Hypothetical protein; similar to glutelin type-B2 precursor | 17.82 | 54.0 | 9.22 | Os02g0248800 | Storage protein | |
| Glutelin type-B5 | 24.04 | 35.6 | 7.06 | Os02g0268300 | Storage protein | |
| Hypothetical protein; similar to aspartic proteinase oryzasin 1 precursor | 7.66 | 55.8 | 7.05 | Os01g0663400 | Proteinase | |
| Blast and wounding induced mitogen-activated protein kinase | 5.22 | 55.2 | 7.94 | Os06g0708000 | Kinase | |
| 2 | Hypothetical protein; glutelin provisional | 40.38 | 35.6 | 7.06 | Os02g0268100 | Storage protein |
| Glutelin type-B1 | 18.03 | 54.1 | 9.22 | Os02g0249800 | Storage protein | |
| Glutelin, provisional | 10.53 | 51.5 | 8.56 | Os10g0400200 | Storage protein | |
| Glutelin, provisional | 9.25 | 51.2 | 8.68 | Os01g0762500 | Storage protein | |
| Transposon protein, putative | 3.57 | 96.8 | 6.35 | Os03g0380600 | Plant transposase | |
| Hypothetical protein; bHLH-MYC and R2R3-MYB transcription factors | 8.91 | 37.5 | 6.98 | Os06g0233800 | Transcription factor | |
| 3 | Glutelin, provisional | 11.01 | 51.2 | 8.68 | Os01g0762500 | Storage protein |
| Cupin family protein | 6.55 | 61.4 | 7.53 | Os03g0336100 | Cupin family protein | |
| Globulin | 4.60 | 19.8 | 6.96 | Os05g0499100 | Storage protein | |
| Lysosomal thiol reductase family protein | 8.20 | 28.6 | 6.24 | Os03g0295800 | Reductase enzyme | |
| Hypothetical protein; universal stress protein-like | 14.12 | 18.2 | 6.71 | Os03g0750000 | Putative stress-related protein | |
| Hypothetical protein; similar to FAD-dependent oxidoreductase family | 4.59 | 61.4 | 6.89 | Os08g0114300 | Reductase enzyme | |
| 4 | Globulin | 9.20 | 19.8 | 6.96 | Os05g0499100 | Storage protein |
| Hypothetical protein; manganese-superoxide dismutase precursor | 16.88 | 25.0 | 7.31 | Os05g0323900 | Superoxide dismutase family protein | |
| Cupin family protein | 3.89 | 61.4 | 7.53 | Os03g0336100 | Cupin family protein | |
| Glutelin, provisional | 5.51 | 51.2 | 8.68 | Os01g0762500 | Storage protein | |
| Pyruvate phosphate dikinase, provisional | 2.74 | 88.1 | 5.63 | Os05g0405000 | Kinase | |
| Hypothetical protein; similar to germin-like protein precursor | 13.26 | 19.6 | 8.72 | Os08g0189100 | Cupin family protein | |
| 5 | Alpha-amylase inhibitors, provisional | 16.25 | 17.3 | 8.03 | Os07g0215500 | Allergenic protein |
| Hypothetical protein; similar to prolamin | 42.38 | 17.0 | 8.72 | Os07g0219400 | Storage protein | |
| Alpha-amylase inhibitors, provisional | 10.19 | 17.0 | 8.03 | Os07g0214600 | Allergenic protein | |
| Hypothetical protein; similar to early nodulin-like protein | 6.74 | 19.3 | 8.34 | Os08g0273300 | Nodulin-like family protein | |
| 6 | Alpha-amylase inhibitors, provisional | 13.31 | 17.3 | 8.03 | Os07g0215500 | Allergenic protein |
| Putative prolamin | 8.15 | 17.0 | 9.17 | Os05g0329200 | Storage protein | |
| Prolamin | 5.31 | 17.1 | 9.38 | Os07g0219400 | Storage protein | |
| Alpha-amylase inhibitors, provisional | 3.81 | 17.0 | 8.03 | Os07g0214600 | Allergenic protein |

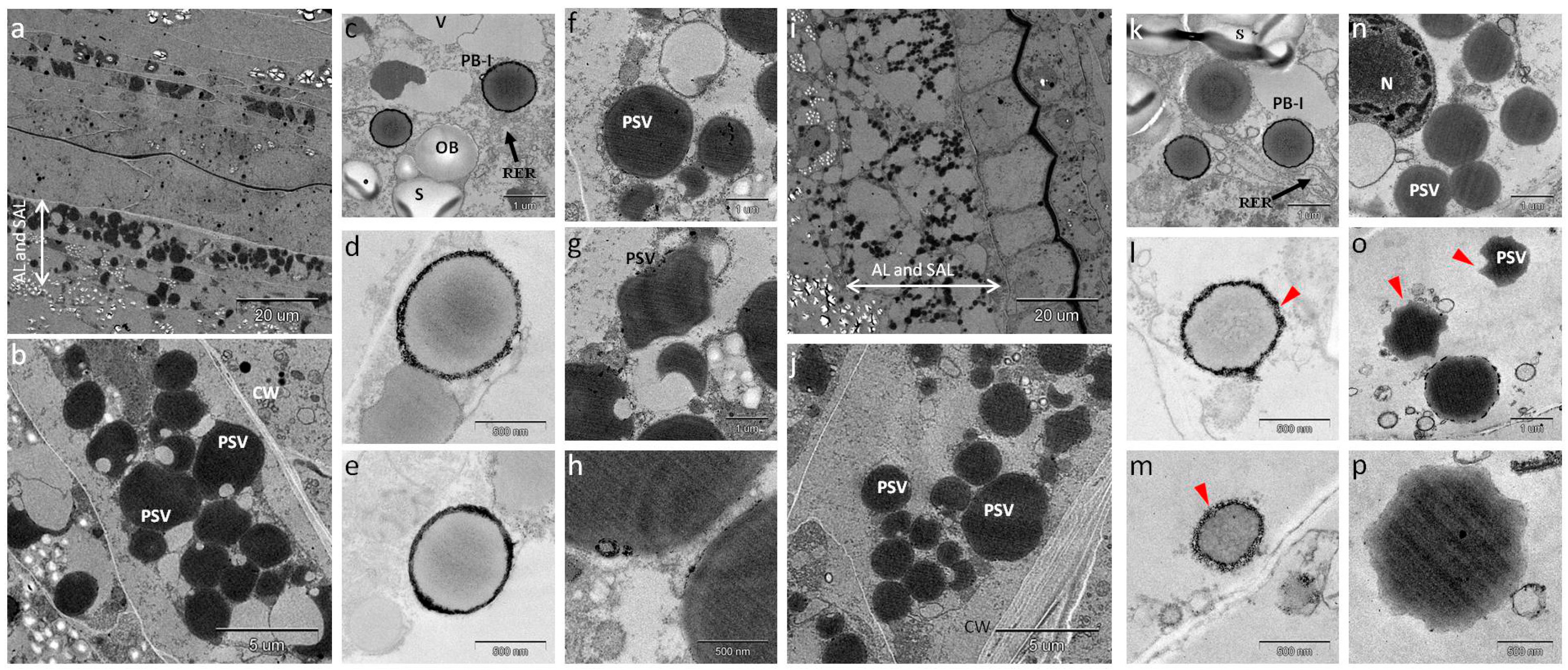
2.6. Observation of Storage Protein Organelles in Immature Seeds Compared with Mature Seeds

3. Discussion
4. Experimental Section
4.1. Plasmid Preparation
4.2. Plant Transformation
4.3. In Situ Western Hybridization
4.4. Sodium Dodecyl Sulfate (SDS)-Polyacrylamide Gel Electrophoresis (PAGE) and Immunoblotting
4.5. RNA Extraction and Gene Expression Analysis
4.6. Seed Storage Proteins (SSP) Fractionation
4.7. Two-Dimensional Gel Electrophoresis (2-DE)
4.8. Microscopic Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Shewry, P.R.; Halford, N.G. Cereal seed storage proteins: Structures, properties and role in grain utilization. J. Exp. Bot. 2002, 53, 947–958. [Google Scholar] [CrossRef] [PubMed]
- Okita, T.W.; Hwang, Y.S.; Hnilo, J.; Kim, W.T.; Aryan, A.P.; Larson, R.; Krishnan, H. Structure and expression of the rice glutelin multigene family. J. Biol. Chem. 1989, 264, 12573–12581. [Google Scholar] [PubMed]
- Takaiwa, F.; Oono, K.; Wing, D.; Kato, A. Sequence of three members and expression of a new major subfamily of glutelin genes from rice. Plant Mol. Biol. 1991, 17, 875–885. [Google Scholar] [CrossRef] [PubMed]
- Kawakatsu, T.; Yamamoto, M.P.; Hirose, S.; Yano, M.; Takaiwa, F. Characterization of a new rice glutelin gene GluD-1 expressed in the starchy endosperm. J. Exp. Bot. 2008, 59, 4233–4245. [Google Scholar] [CrossRef] [PubMed]
- Qu, L.Q.; Takaiwa, F. Evaluation of tissue specificity and expression strength of rice seed component gene promoters in transgenic rice. Plant Biotechnol. J. 2004, 2, 113–125. [Google Scholar] [CrossRef] [PubMed]
- Qu, L.Q.; Xing, Y.P.; Liu, W.X.; Xu, X.P.; Song, Y.R. Expression pattern and activity of six glutelin gene promoters in transgenic rice. J. Exp. Bot. 2008, 59, 2417–2424. [Google Scholar] [CrossRef] [PubMed]
- Doroshenk, K.A.; Tian, L.; Crofts, A.J.; Kumamaru, T.; Okita, T.W. Characterization of RNA binding protein RBP-P reveals a possible role in rice glutelin gene expression and RNA localization. Plant Mol. Biol. 2014, 85, 381–394. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, M.; Kumamaru, T.; Satoh, H.; Iwata, N.; Omura, T.; Kasai, Z.; Tanaka, K. Purification of protein body-I of rice seed and its polypeptide composition. Plant Cell Physiol. 1987, 28, 1517–1527. [Google Scholar]
- Mitsukawa, N.; Konishi, R.; Kidzu, K.; Ohtsuki, K.; Masumura, T.; Tanaka, K. Amino acid sequencing and cDNA cloning of rice seed storage proteins, the 13 kDa prolamins, extracted from type I protein bodies. Plant Biotechnol. 1999, 16, 103–113. [Google Scholar] [CrossRef]
- Saito, Y.; Shigemitsu, T.; Yamasaki, R.; Sasou, A.; Goto, F.; Kishida, K.; Kuroda, M.; Tanaka, K.; Morita, S.; Satoh, S.; et al. Formation mechanism of the internal structure of type I protein bodies in rice endosperm: Relationship between the localization of prolamin species and the expression of individual genes. Plant J. 2012, 70, 1043–1055. [Google Scholar] [CrossRef] [PubMed]
- Nagamine, A.; Matsusaka, H.; Ushijima, T.; Kawagoe, Y.; Ogawa, M.; Okita, T.W.; Kumamaru, T. A role for the cysteine-rich 10 kDa prolamin in protein body I formation in rice. Plant Cell Physiol. 2011, 52, 1003–1016. [Google Scholar] [CrossRef] [PubMed]
- Shorrosh, B.S.; Wen, L.; Zen, K.C.; Huang, J.K.; Pan, J.S.; Hermodson, M.A.; Tanaka, K.; Muthukrishnan, S.; Reeck, G.R. A novel cereal storage protein: Molecular genetics of the 19 kDa globulin of rice. Plant Mol. Biol. 1992, 18, 151–154. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, H.B.; Pueppke, S.G. Nucleotide sequence of an abundant rice seed globulin: Homology with the high molecular weight glutelins of wheat, rye and triticale. Biochem. Biophys. Res. Commun. 1993, 193, 460–466. [Google Scholar] [CrossRef] [PubMed]
- Nakase, M.; Hotta, H.; Adachi, T.; Aoki, N.; Nakamura, R.; Masumura, T.; Tanaka, K.; Matsuda, T. Cloning of the rice seed α-globulin-encoding gene: Sequence similarity of the 5′-flanking region to those of the genes encoding wheat high-molecular-weight glutenin and barley D hordein. Gene 1996, 170, 223–236. [Google Scholar] [CrossRef]
- Gu, Y.Q.; Wanjugi, H.; Coleman-Derr, D.; Kong, X.; Anderson, O.D. Conserved globulin gene across eight grass genomes identify fundamental units of the loci encoding seed storage proteins. Funct. Integr. Genomics 2010, 10, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Yoon, U.H.; Lee, J.; Lee, G.S.; Kim, Y.M.; Hahn, J.H.; Ji, H.S.; Yoon, S.W.; Lee, J.Y.; Kim, T.H. Current status of structural and expression analysis of seed storage protein genes in japonica rice. Korean J. Soc. Int. Agric. 2012, 24, 369–379. [Google Scholar]
- Kumamaru, T.; Satoh, H.; Iwata, N.; Omura, T.; Ogawa, M.; Tanaka, K. Mutants for rice storage proteins: 1. Screening of mutants for rice storage proteins of protein bodies in the starchy endosperm. Theor. Appl. Genet. 1988, 76, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, M.; Kumamaru, T.; Satoh, H.; Omura, T.; Park, T.; Shintaku, K.; Baba, K. Mutants for rice storage proteins: 2. Isolation and characterization of protein bodies from rice mutants. Theor. Appl. Genet. 1989, 78, 305–310. [Google Scholar] [CrossRef] [PubMed]
- Ueda, Y.; Satoh-Cruz, M.; Matsusaka, H.; Takemoto-Kuno, Y.; Fukuda, M.; Okita, T.W.; Ogawa, M.; Satoh, H.; Kumamaru, T. Gene-gene interactions between mutants that accumulate abnormally high amounts of proglutelin in rice seed. Breed Sci. 2010, 60, 568–574. [Google Scholar] [CrossRef]
- Tian, L.; Dai, L.L.; Yin, Z.J.; Fukuda, M.; Kumamaru, T.; Dong, X.B.; Xu, X.P.; Qu, L.Q. Small GTPase Sar1 is crucial for proglutelin and α-globulin export from the endoplasmic reticulum in rice endosperm. J. Exp. Bot. 2013, 64, 2831–2845. [Google Scholar] [CrossRef] [PubMed]
- Takemoto, Y.; Coughlan, S.J.; Okita, T.W.; Satoh, H.; Ogawa, M.; Kumamaru, T. The rice mutant esp2 greatly accumulates the glutelin precursor and deletes the protein disulfide isomerase. Plant Physiol. 2002, 128, 1212–1222. [Google Scholar] [CrossRef] [PubMed]
- Satoh-Cruz, M.; Crofts, A.J.; Takemoto-Kuno, Y.; Sugino, A.; Washida, H.; Crofts, N.; Okita, T.W.; Ogawa, M.; Satoh, H.; Kumamaru, T. Protein disulfide isomerase like 1-1 participates in the maturation of proglutelin within the endoplasmic reticulum in rice endosperm. Plant Cell Physiol. 2010, 51, 1581–1593. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Wang, Y.; Liu, F.; Zhou, K.; Ding, Y.; Zhou, F.; Wang, Y.; Liu, K.; Gan, L.; Ma, W.; et al. Glutelin Precursor Accumulation3 encodes a regulator of post-Golgi vesicular traffic essential for vacuolar protein sorting in rice endosperm. Plant Cell 2014, 26, 410–425. [Google Scholar] [CrossRef] [PubMed]
- Kusaba, M.; Miyahara, K.; Iida, S.; Fukuoka, H.; Takano, T.; Sassa, H.; Nishimura, M.; Nishio, T. Low glutelin content1: A dominant mutation that suppresses the glutelin multigene family via RNA silencing in rice. Plant Cell 2003, 15, 1455–1467. [Google Scholar] [CrossRef] [PubMed]
- Kawakatsu, T.; Hirose, S.; Yasuda, H.; Takaiwa, F. Reducing rice seed storage protein accumulation leads to changes in nutrient quality and storage organelle formation. Plant Physiol. 2010, 154, 1842–1854. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Lee, J.Y.; Yoon, U.H.; Lim, S.H.; Kim, Y.M. Effects of reduced prolamin on seed storage protein composition and the nutritional quality of rice. Int. J. Mol. Sci. 2013, 14, 17073–17084. [Google Scholar] [CrossRef] [PubMed]
- Iida, S.; Amano, E.; Nishio, T. A rice (Oryza sativa L.) mutant having a low content of glutelin and a high content of prolamine. Theor. Appl. Genet. 1993, 87, 374–378. [Google Scholar] [CrossRef] [PubMed]
- Onda, Y.; Kumamaru, T.; Kawagoe, Y. ER membrane-localized oxidoreductase Ero1 is required for disulfide bond formation in the rice endosperm. Proc. Natl. Acad. Sci. USA 2009, 106, 14156–14161. [Google Scholar] [CrossRef] [PubMed]
- Oono, Y.; Wakasa, Y.; Hirose, S.; Yang, L.; Sakuta, C.; Takaiwa, F. Analysis of ER stress in developing rice endosperm accumulating β-amyloid peptide. Plant Biotechnol. J. 2010, 8, 691–718. [Google Scholar] [CrossRef] [PubMed]
- Wakasa, Y.; Yasuda, H.; Oono, Y.; Kawakatsu, T.; Hirose, S.; Takahashi, H.; Hayashi, S.; Yang, L.; Takaiwa, F. Expression of ER quality control-related genes in response to changes in BiP1 levels in developing rice endosperm. Plant J. 2011, 65, 675–689. [Google Scholar] [CrossRef] [PubMed]
- Onda, Y.; Nagamine, A.; Sakurai, M.; Kumamaru, T.; Ogawa, M.; Kawagoe, Y. Distinct roles of protein disulfide isomerase and P5 sulfhydryl oxidoreductases in multiple pathways for oxidation of structurally diverse storage proteins in rice. Plant Cell 2011, 23, 210–223. [Google Scholar] [CrossRef] [PubMed]
- Onda, Y.; Kawagoe, Y. P5-type sulfhydryl oxidoreductase promotes the sorting of proteins to protein body I in rice endosperm cells. Plant Signal. Behav. 2013, 8, e23075. [Google Scholar] [CrossRef] [PubMed]
- Shewry, P.R.; Tatham, A.S. The prolamin storage proteins of cereal seeds: Structure and evolution. Biochem. J. 1990, 267, 1–12. [Google Scholar] [PubMed]
- Hattori, T.; Terada, T.; Hamasuna, S.T. Sequence and functional analyses of the rice gene homologous to the maize Vp1. Plant Mol. Biol. 1994, 24, 805–810. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, H.B.; White, J.A. Morphometric analysis of rice seed protein bodies (Implication for a significant contribution of prolamine to the total protein content of rice endosperm. Plant Physiol. 1995, 109, 1491–1495. [Google Scholar] [PubMed]
- Katsube-Tanaka, T.; Duldulao, J.B.; Kimura, Y.; Iida, S.; Yamaguchi, T.; Nakano, J.; Utsumi, S. The two subfamilies of rice glutelin differ in both primary and higher-order structures. Biochim. Biophys. Acta 2004, 1699, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Washida, H.; Sugino, A.; Doroshenk, K.A.; Satoh-Cruz, M.; Nagamine, A.; Katsube-Tanaka, T.; Ogawa, M.; Kumamaru, T.; Satoh, H.; Okita, T.W. RNA targeting to a specific ER sub-domain is required for efficient transport and packaging of α-globulins to the protein storage vacuole in developing rice endosperm. Plant J. 2012, 70, 471–479. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.M.; Lee, J.Y.; Lee, T.; Lee, Y.H.; Kim, S.H.; Kang, S.H.; Yoon, U.H.; Ha, S.H.; Lim, S.H. The suppression of the glutelin storage protein gene in transgenic rice seeds results in a higher yield of recombinant protein. Plant Biotechnol. Rep. 2012, 6, 347–353. [Google Scholar] [CrossRef]
- Qu, L.Q.; Tada, Y.; Takaiwa, F. In situ western hybridization: A new, highly sensitive technique to detect foreign and endogenous protein distribution in rice seeds. Plant Cell Rep. 2003, 22, 282–285. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, H.-J.; Jo, Y.-M.; Lee, J.-Y.; Lim, S.-H.; Kim, Y.-M. Lack of Globulin Synthesis during Seed Development Alters Accumulation of Seed Storage Proteins in Rice. Int. J. Mol. Sci. 2015, 16, 14717-14736. https://doi.org/10.3390/ijms160714717
Lee H-J, Jo Y-M, Lee J-Y, Lim S-H, Kim Y-M. Lack of Globulin Synthesis during Seed Development Alters Accumulation of Seed Storage Proteins in Rice. International Journal of Molecular Sciences. 2015; 16(7):14717-14736. https://doi.org/10.3390/ijms160714717
Chicago/Turabian StyleLee, Hye-Jung, Yeong-Min Jo, Jong-Yeol Lee, Sun-Hyung Lim, and Young-Mi Kim. 2015. "Lack of Globulin Synthesis during Seed Development Alters Accumulation of Seed Storage Proteins in Rice" International Journal of Molecular Sciences 16, no. 7: 14717-14736. https://doi.org/10.3390/ijms160714717