
Circadian Rhythm Shapes the Gut Microbiota Affecting Host Radiosensitivity

Abstract
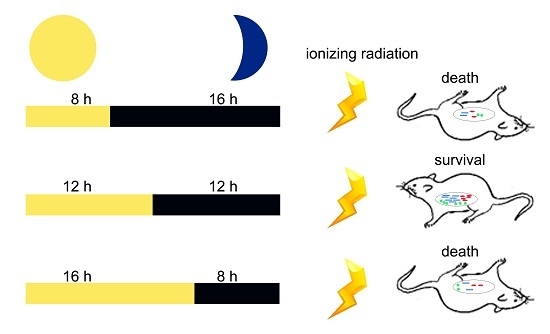
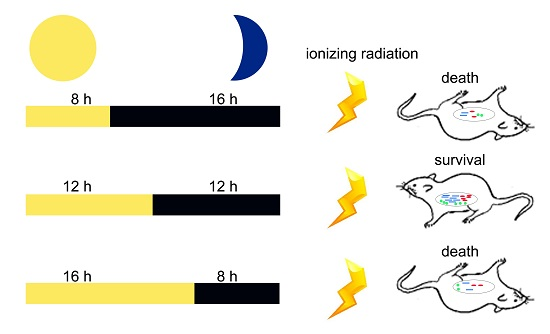
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
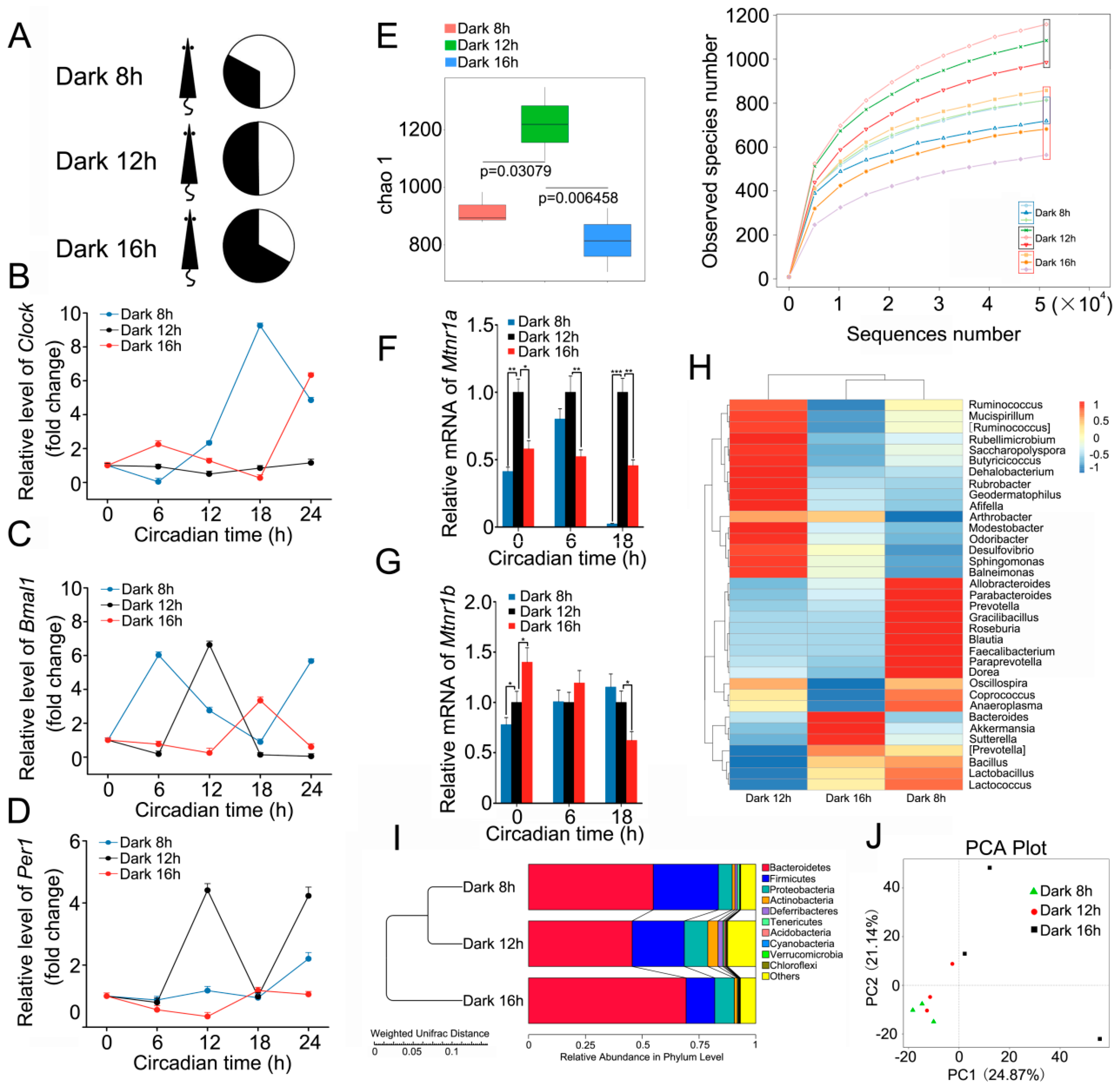
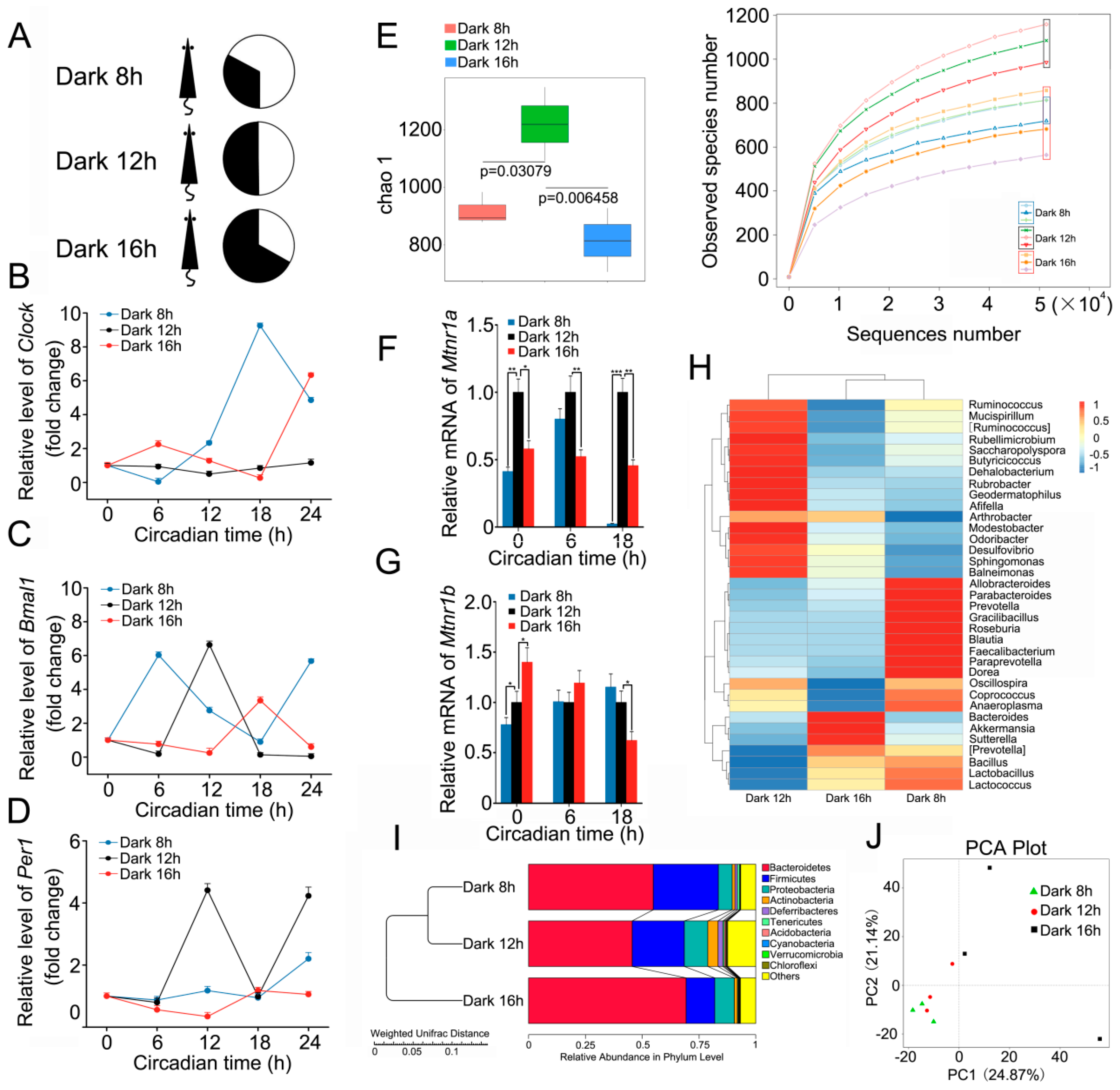
2.1. Circadian Rhythm Disturbance Shapes Intestinal Bacterial Composition in a Mouse Model
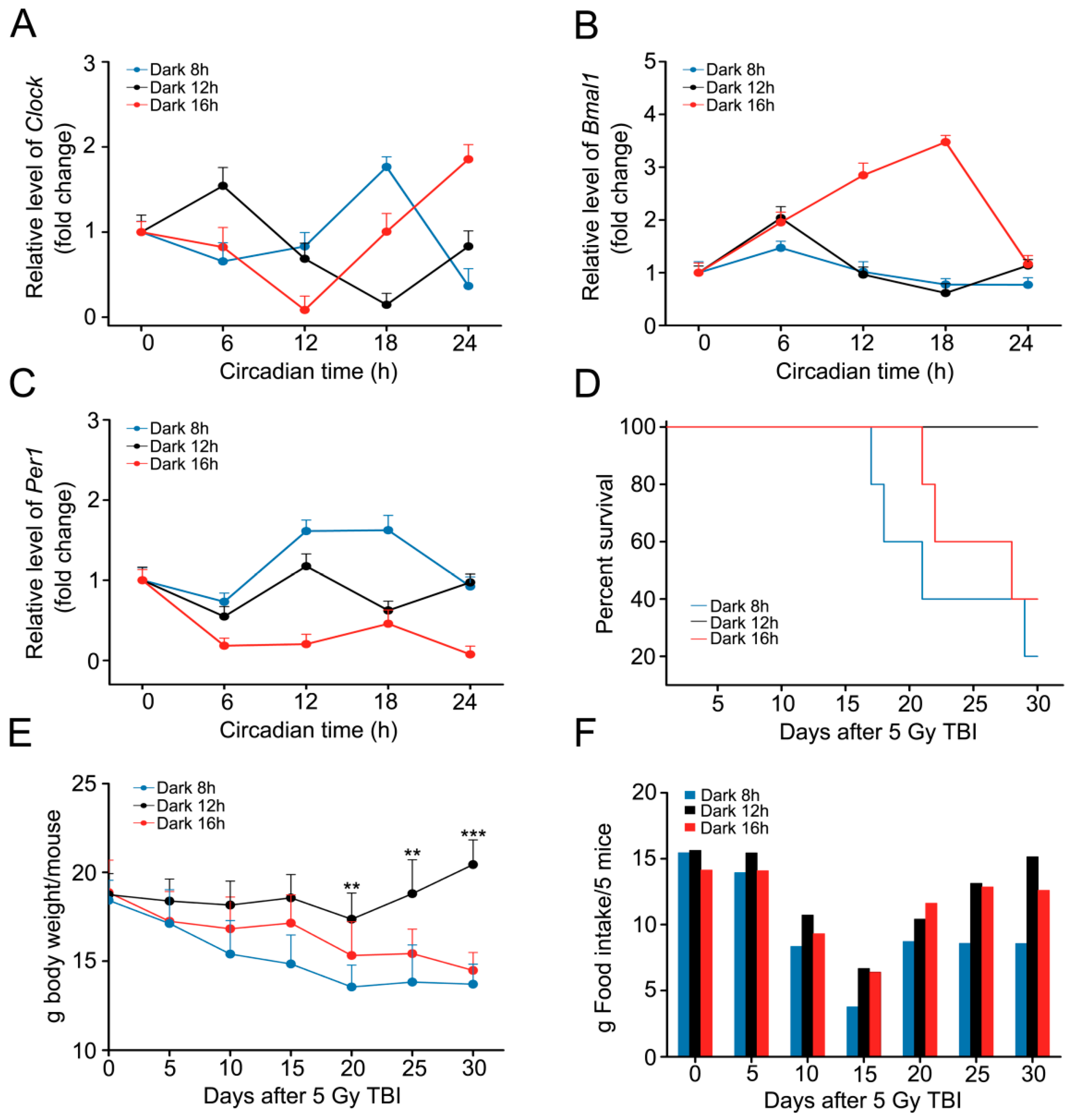
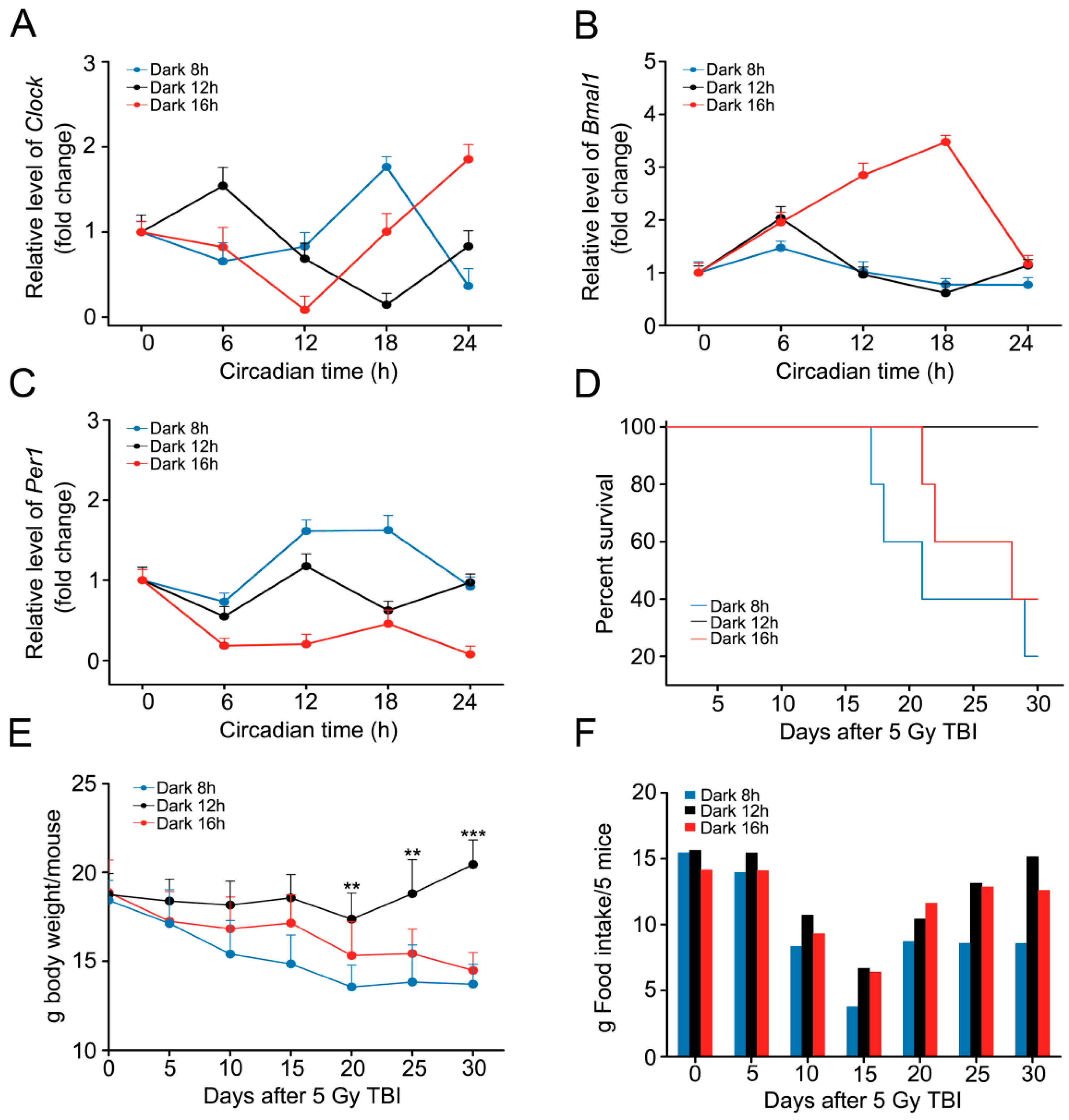
2.2. Circadian Rhythm Disorder Impairs the Radioresistance of Mice
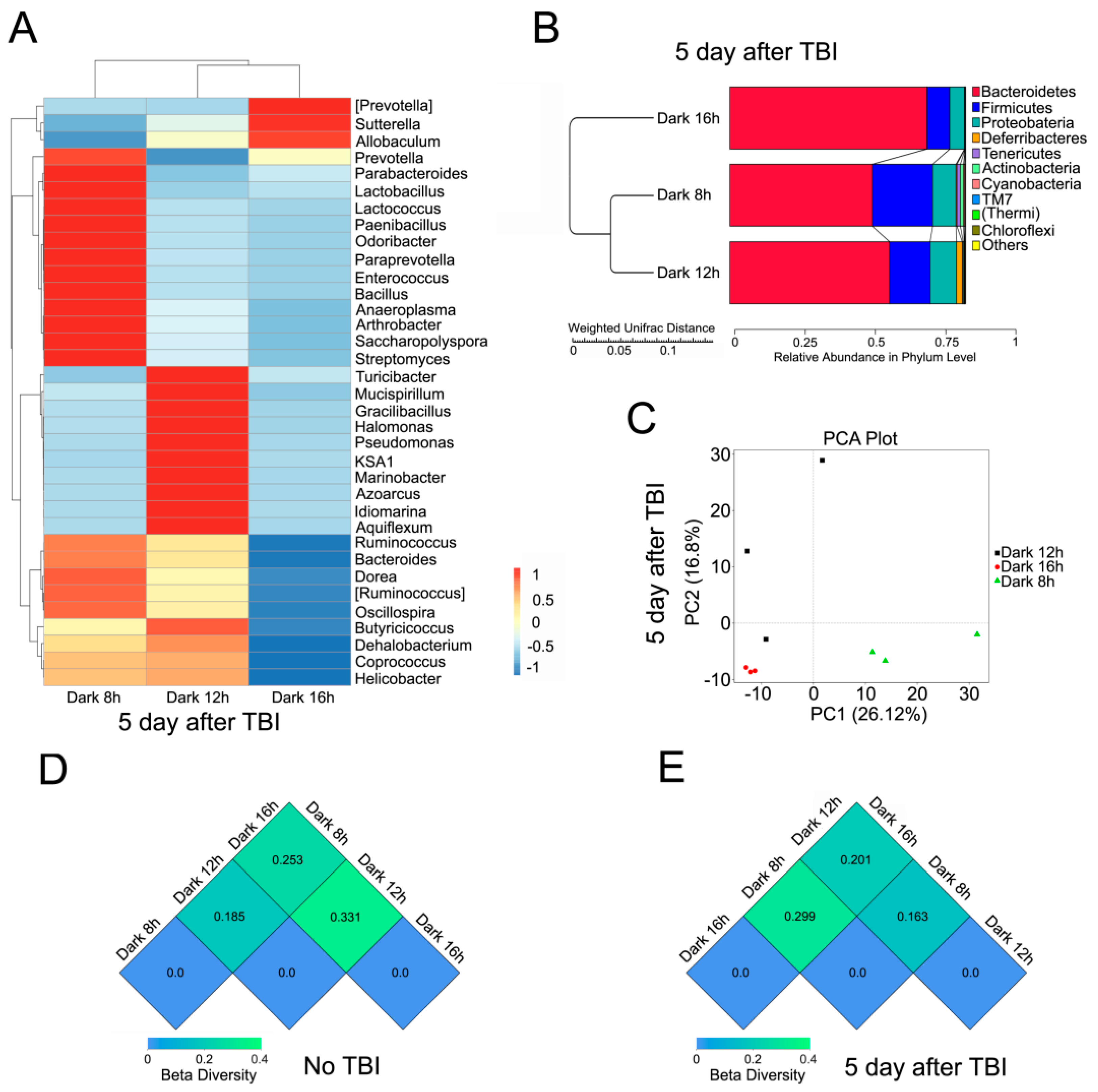
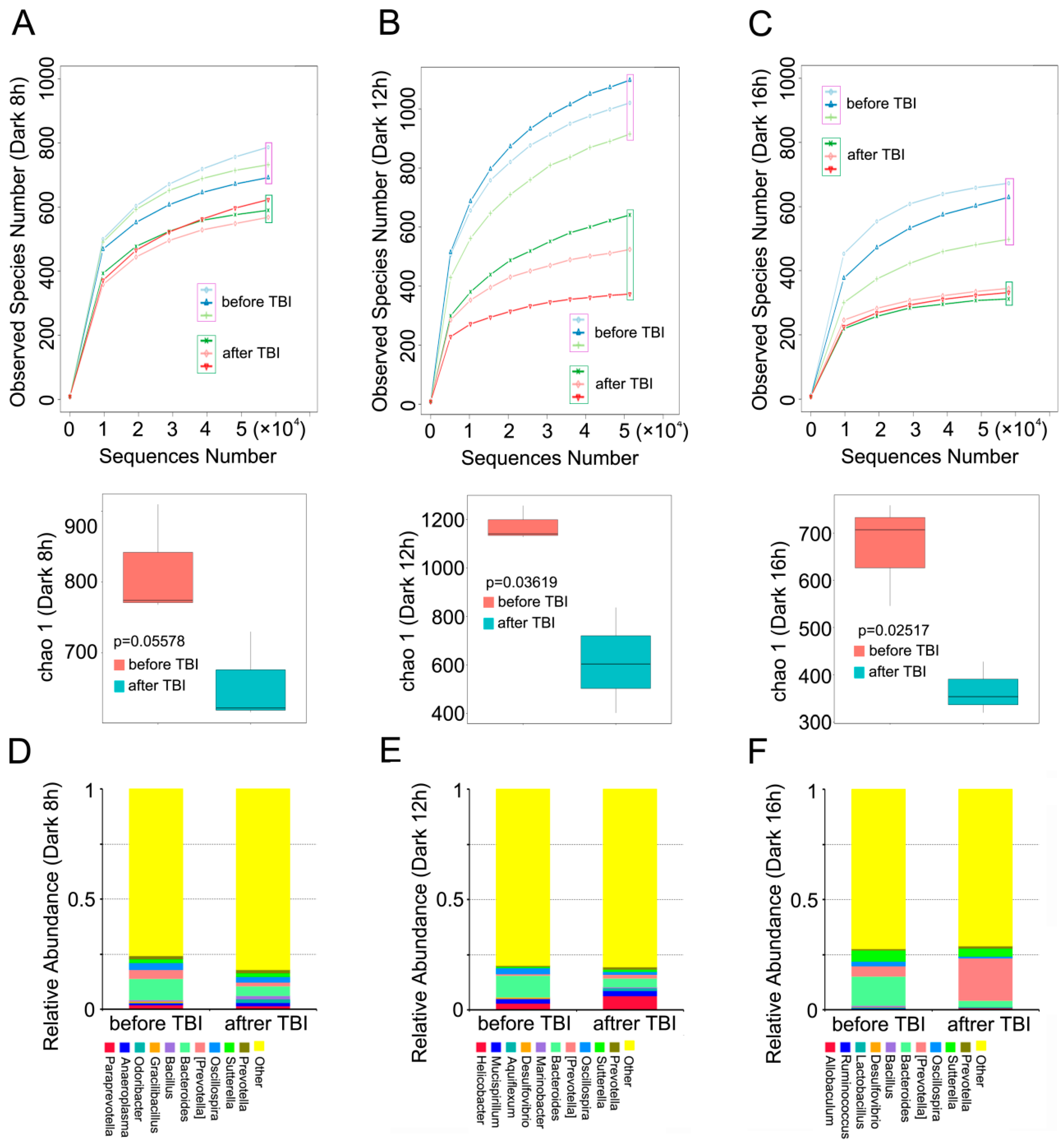
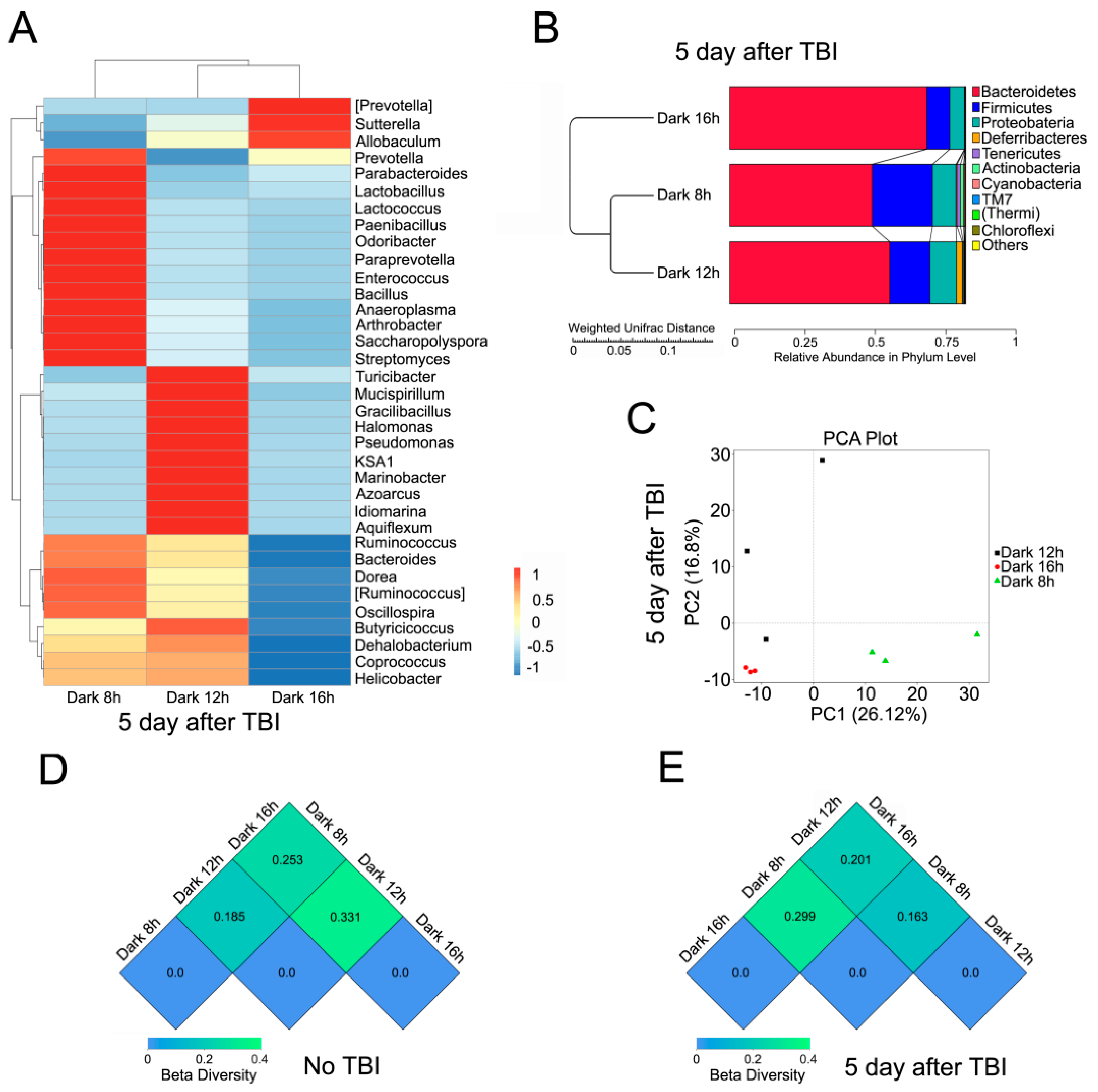
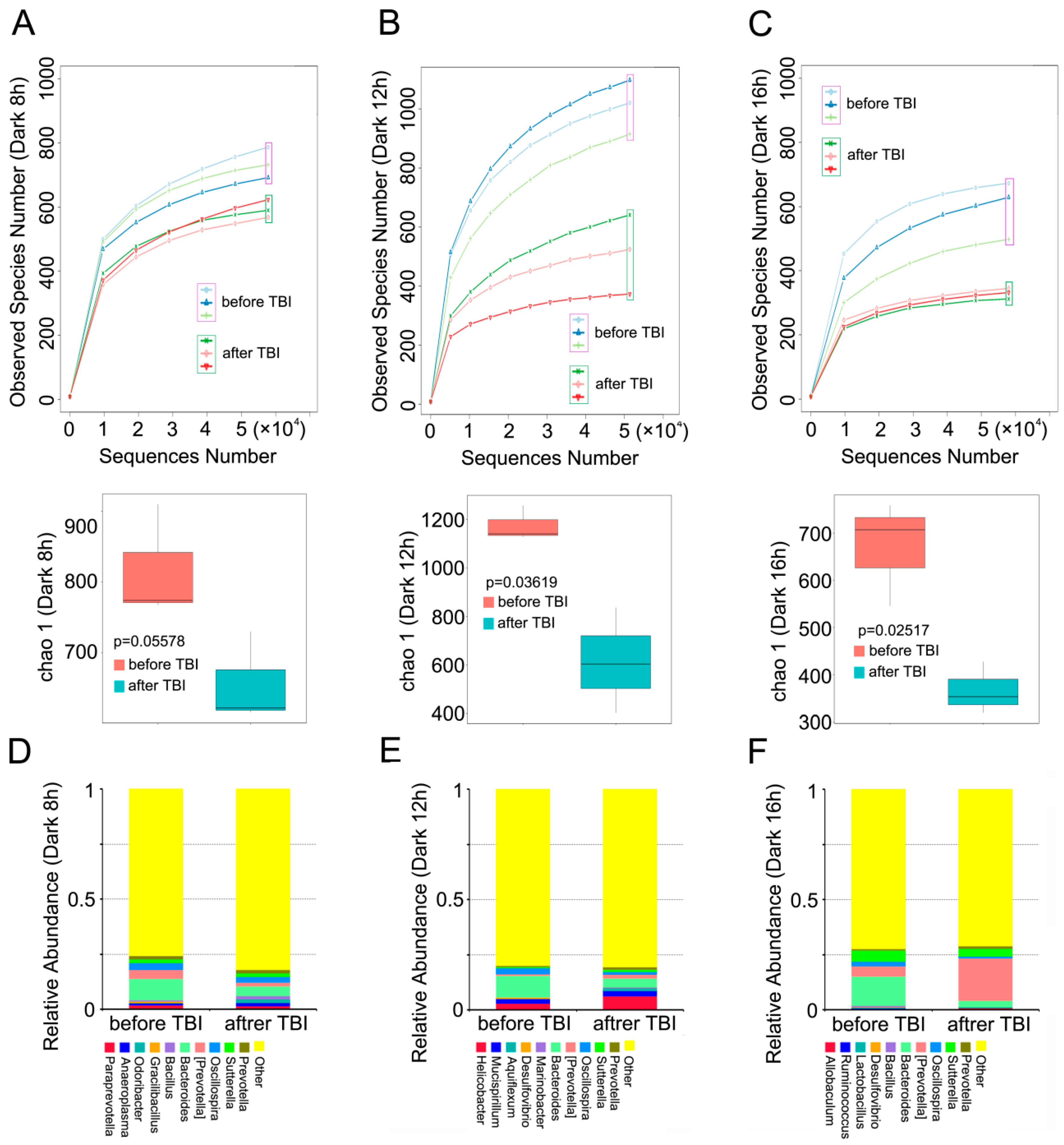
2.3. Circadian Rhythm Affects the Structure of Gut Bacterial Composition after Irradiation
2.4. Circadian Rhythm Governs the Response of Gut Bacterial Composition Shift to Irradiation
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Irradiation Study
4.3. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
| Clock: forward, AGTTAGGGCTGAAAGACGGC, reverse, TAGAGGAGGCAGAAGGAGTTGG; |
| Bmal1: forward, AGATAAACTCACCGTGCTAAG, reverse, AGATAAACTCACCGTGCTAAG; |
| Per1: forward, AGGGTGAGCCTTGTGCCAT, reverse, AGATGGTGTAGTAGAGCCATAG; |
| Mtnr1a: forward, TGGCTGTTTACCCTTATCCC, reverse, AAACCACCACTGCTATCGTG; |
| Mtnr1b: forward, GTGTCATTGGCTCTGTCTTCA, reverse, TGTGCTGGCTGTCTGGATGAA; |
| GAPDH: forward, TGTTTCCTCGTCCCGTAGA, reverse, CAATCTCCACTTTGCCACTG. |
4.4. Bacterial Diversity Analysis
4.5. Statistical Analyses
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Sahar, S.; Sassone-Corsi, P. Metabolism and cancer: The circadian clock connection. Nat. Rev. Cancer 2009, 9, 886–896. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, J.S.; Hong, H.K.; Ko, C.H.; McDearmon, E.L. The genetics of mammalian circadian order and disorder: Implications for physiology and disease. Nat. Rev. Genet. 2008, 9, 764–775. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Xu, H.; Liu, Y.; Mu, L.; Xiao, J.; Zhao, H. Diurnal expression of the Per2 gene and protein in the lateral habenular nucleus. Int. J. Mol. Sci. 2015, 16, 16740–16749. [Google Scholar] [CrossRef] [PubMed]
- Kondratova, A.A.; Kondratov, R.V. The circadian clock and pathology of the ageing brain. Nat. Rev. Neurosci. 2012, 13, 325–335. [Google Scholar] [CrossRef] [PubMed]
- Barker, H.E.; Paget, J.T.; Khan, A.A.; Harrington, K.J. The tumour microenvironment after radiotherapy: Mechanisms of resistance and recurrence. Nat. Rev. Cancer 2015, 15, 409–425. [Google Scholar] [CrossRef] [PubMed]
- Taniguchi, C.M.; Miao, Y.R.; Diep, A.N.; Wu, C.; Rankin, E.B.; Atwood, T.F.; Xing, L.; Giaccia, A.J. PHD inhibition mitigates and protects against radiation-induced gastrointestinal toxicity via HIF2. Sci. Transl. Med. 2014, 6. [Google Scholar] [CrossRef] [PubMed]
- Belkaid, Y.; Hand, T.W. Role of the microbiota in immunity and inflammation. Cell 2014, 157, 121–141. [Google Scholar] [CrossRef] [PubMed]
- Lichtman, J.S.; Sonnenburg, J.L.; Elias, J.E. Monitoring host responses to the gut microbiota. ISME J. 2015, 9, 1908–1915. [Google Scholar] [CrossRef] [PubMed]
- Aron-Wisnewsky, J.; Clement, K. The gut microbiome, diet, and links to cardiometabolic and chronic disorders. Nat. Rev. Nephrol. 2016, 12, 169–181. [Google Scholar] [CrossRef] [PubMed]
- Delzenne, N.M.; Neyrinck, A.M.; Backhed, F.; Cani, P.D. Targeting gut microbiota in obesity: Effects of prebiotics and probiotics. Nat. Rev. Nephrol. 2011, 7, 639–646. [Google Scholar] [CrossRef] [PubMed]
- Sanduzzi Zamparelli, M.; Compare, D.; Coccoli, P.; Rocco, A.; Nardone, O.M.; Marrone, G.; Gasbarrini, A.; Grieco, A.; Nardone, G.; Miele, L. The metabolic role of gut microbiota in the development of nonalcoholic fatty liver disease and cardiovascular disease. Int. J. Mol. Sci. 2016, 17. [Google Scholar] [CrossRef] [PubMed]
- Lozupone, C.A.; Stombaugh, J.I.; Gordon, J.I.; Jansson, J.K.; Knight, R. Diversity, stability and resilience of the human gut microbiota. Nature 2012, 489, 220–230. [Google Scholar] [CrossRef] [PubMed]
- Rosselot, A.E.; Hong, C.I.; Moore, S.R. Rhythm and bugs: Circadian clocks, gut microbiota, and enteric infections. Curr. Opin. Gastroenterol. 2016, 32, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Paulose, J.K.; Wright, J.M.; Patel, A.G.; Cassone, V.M. Human Gut Bacteria Are Sensitive to Melatonin and Express Endogenous Circadian Rhythmicity. PLoS ONE 2016, 11, e0146643. [Google Scholar] [CrossRef] [PubMed]
- LeGates, T.A.; Fernandez, D.C.; Hattar, S. Light as a central modulator of circadian rhythms, sleep and affect. Nat. Rev. Neurosci. 2014, 15, 443–454. [Google Scholar] [CrossRef] [PubMed]
- Bracci, M.; Ciarapica, V.; Copertaro, A.; Barbaresi, M.; Manzella, N.; Tomasetti, M.; Gaetani, S.; Monaco, F.; Amati, M.; Valentino, M.; et al. Peripheral skin temperature and circadian biological clock in shift nurses after a day off. Int. J. Mol. Sci. 2016, 17. [Google Scholar] [CrossRef] [PubMed]
- O'Neill, J.S.; Reddy, A.B. Circadian clocks in human red blood cells. Nature 2011, 469, 498–503. [Google Scholar] [CrossRef] [PubMed]
- Gamble, K.L.; Berry, R.; Frank, S.J.; Young, M.E. Circadian clock control of endocrine factors. Nat. Rev. Endocrinol. 2014, 10, 466–475. [Google Scholar] [CrossRef] [PubMed]
- Scheiermann, C.; Kunisaki, Y.; Frenette, P.S. Circadian control of the immune system. Nat. Rev. Immunol. 2013, 13, 190–198. [Google Scholar] [CrossRef] [PubMed]
- Videnovic, A.; Lazar, A.S.; Barker, R.A.; Overeem, S. “The clocks that time us”—Circadian rhythms in neurodegenerative disorders. Nat. Rev. Neurol. 2014, 10, 683–693. [Google Scholar] [CrossRef] [PubMed]
- Cui, M.; Zheng, M.; Sun, B.; Wang, Y.; Ye, L.; Zhang, X. A long noncoding RNA perturbs the circadian rhythm of hepatoma cells to facilitate hepatocarcinogenesis. Neoplasia 2015, 17, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Takeda, N.; Maemura, K. Circadian clock and the onset of cardiovascular events. Hypertens. Res. 2016, 39, 383–390. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, I.; Yoshida, Y.; Minamino, T. A role for circadian clock in metabolic disease. Hypertens. Res. 2016, 39, 483–491. [Google Scholar] [CrossRef] [PubMed]
- Verdu, E.F.; Galipeau, H.J.; Jabri, B. Novel players in coeliac disease pathogenesis: Role of the gut microbiota. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Chan, C.W.; Wong, R.S.; Law, P.T.; Wong, C.L.; Tsui, S.K.; Tang, W.P.; Sit, J.W. Environmental factors associated with altered gut microbiota in children with eczema: A systematic review. Int. J. Mol. Sci. 2016, 17. [Google Scholar] [CrossRef] [PubMed]
- DuPont, A.W.; DuPont, H.L. The intestinal microbiota and chronic disorders of the gut. Nat. Rev. Gastroenterol. Hepatol. 2011, 8, 523–531. [Google Scholar] [CrossRef] [PubMed]
- Vogtmann, E.; Goedert, J.J. Epidemiologic studies of the human microbiome and cancer. Br. J. Cancer 2016, 114, 237–242. [Google Scholar] [CrossRef] [PubMed]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cui, M.; Xiao, H.; Luo, D.; Zhang, X.; Zhao, S.; Zheng, Q.; Li, Y.; Zhao, Y.; Dong, J.; Li, H.; et al. Circadian Rhythm Shapes the Gut Microbiota Affecting Host Radiosensitivity. Int. J. Mol. Sci. 2016, 17, 1786. https://doi.org/10.3390/ijms17111786
Cui M, Xiao H, Luo D, Zhang X, Zhao S, Zheng Q, Li Y, Zhao Y, Dong J, Li H, et al. Circadian Rhythm Shapes the Gut Microbiota Affecting Host Radiosensitivity. International Journal of Molecular Sciences. 2016; 17(11):1786. https://doi.org/10.3390/ijms17111786
Chicago/Turabian StyleCui, Ming, Huiwen Xiao, Dan Luo, Xin Zhang, Shuyi Zhao, Qisheng Zheng, Yuan Li, Yu Zhao, Jiali Dong, Hang Li, and et al. 2016. "Circadian Rhythm Shapes the Gut Microbiota Affecting Host Radiosensitivity" International Journal of Molecular Sciences 17, no. 11: 1786. https://doi.org/10.3390/ijms17111786