
Anti-Inflammatory Effects of Chloranthalactone B in LPS-Stimulated RAW264.7 Cells


 ,
,
Abstract
:
1. Introduction
2. Results and Discussion
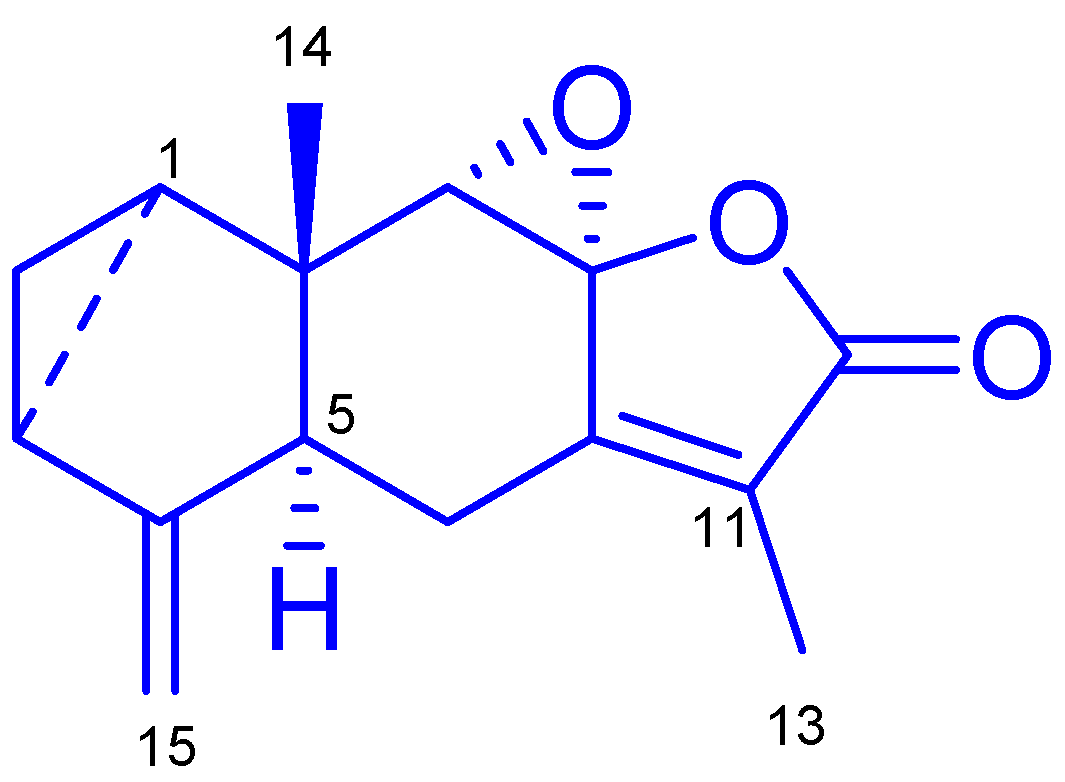
2.1. Elucidation of the Chemical Structure of the Isolated Compound
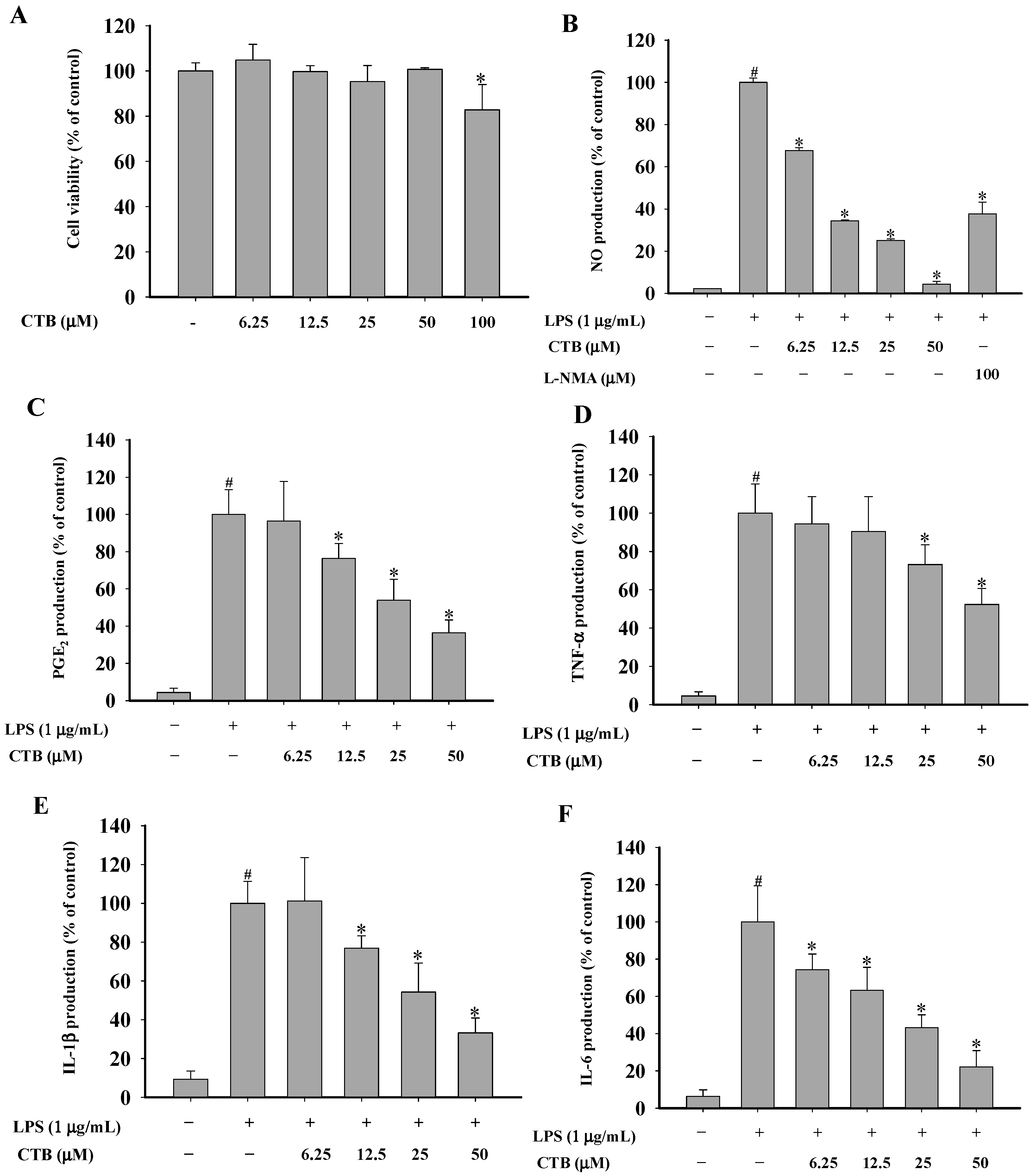
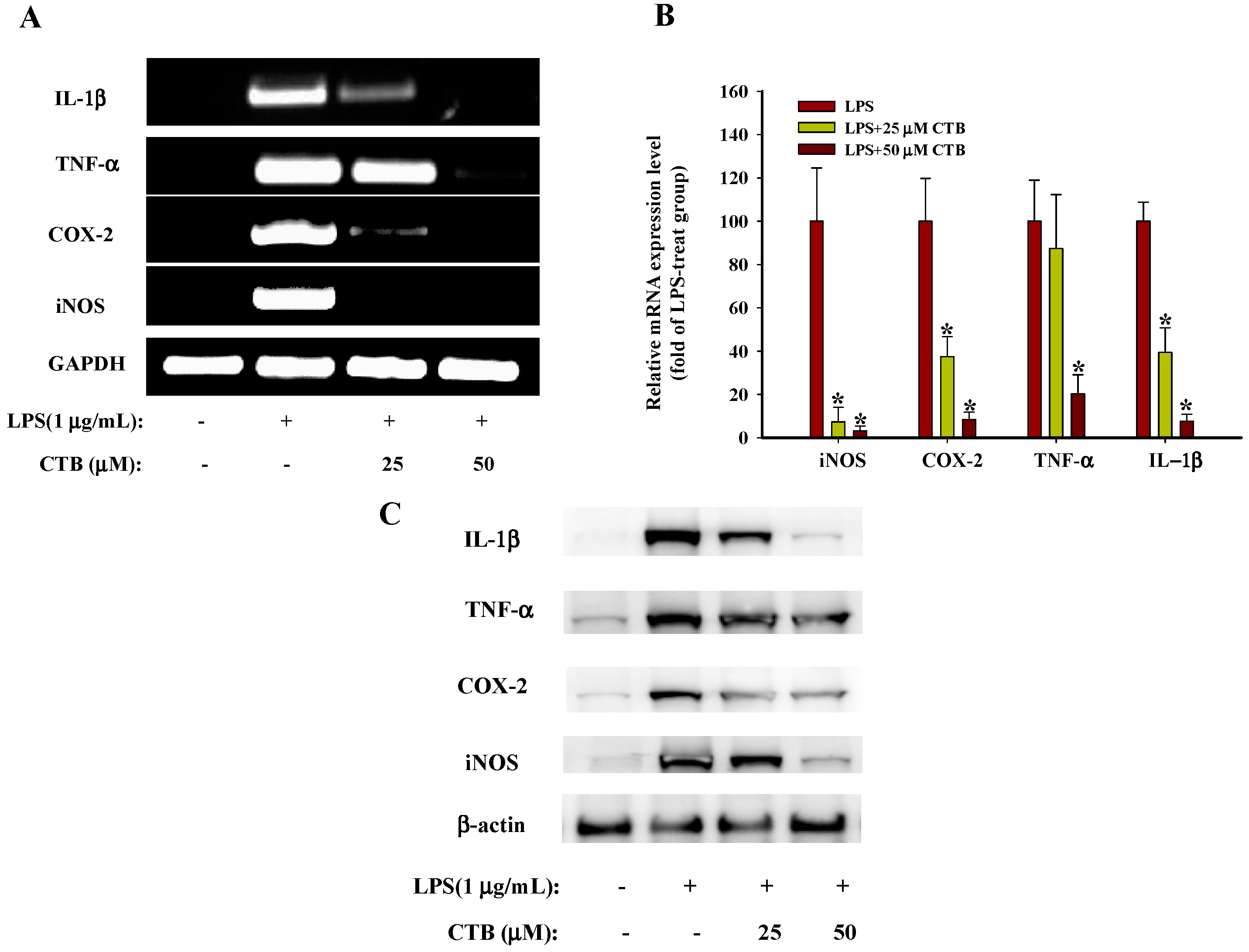
2.2. The Effects of Chloranthalactone B (CTB) on the Production of Inflammatory Mediators in Lipopolysaccharide (LPS)-Activated RAW 264.7 Cells
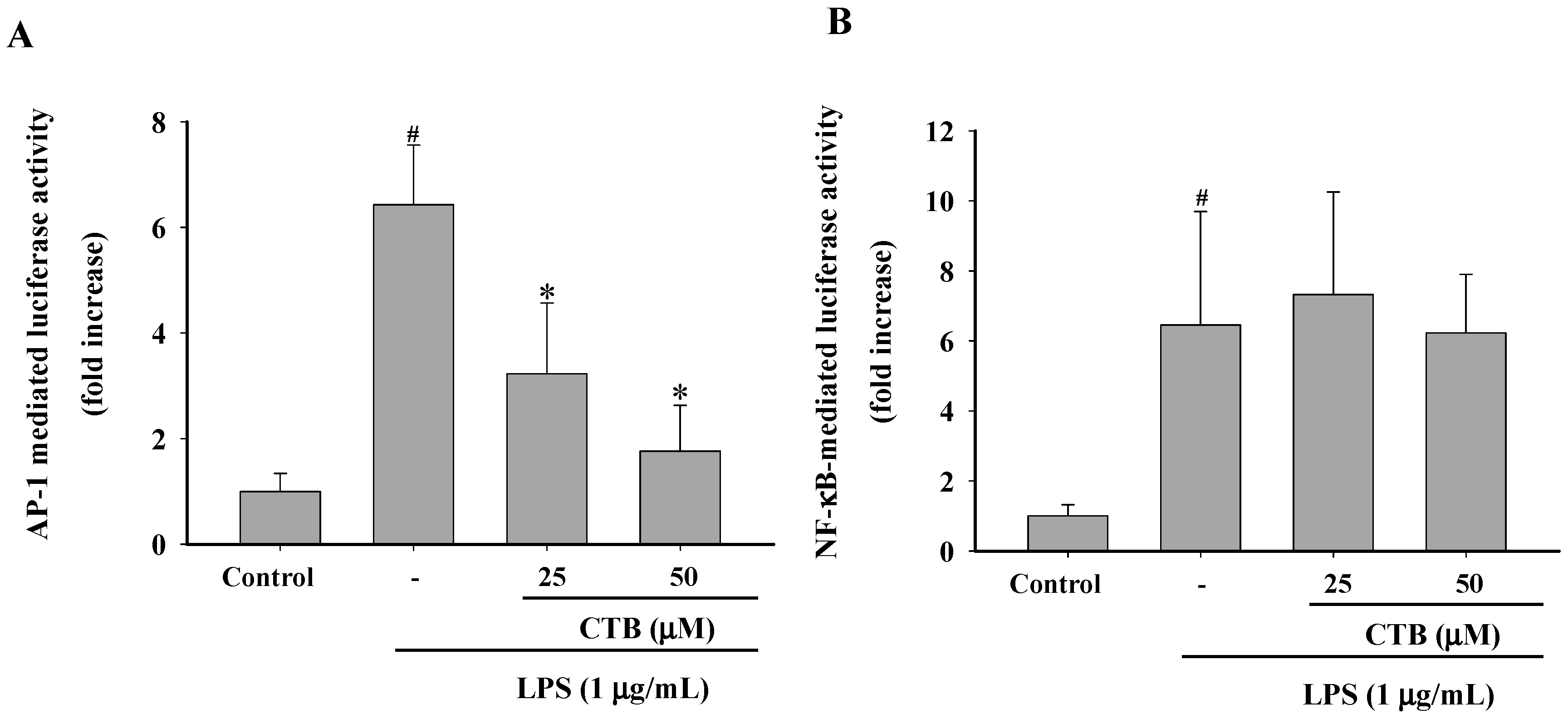
2.3. Effects of CTB on the Expression of Inflammatory Genes and Their Transcriptional Activation
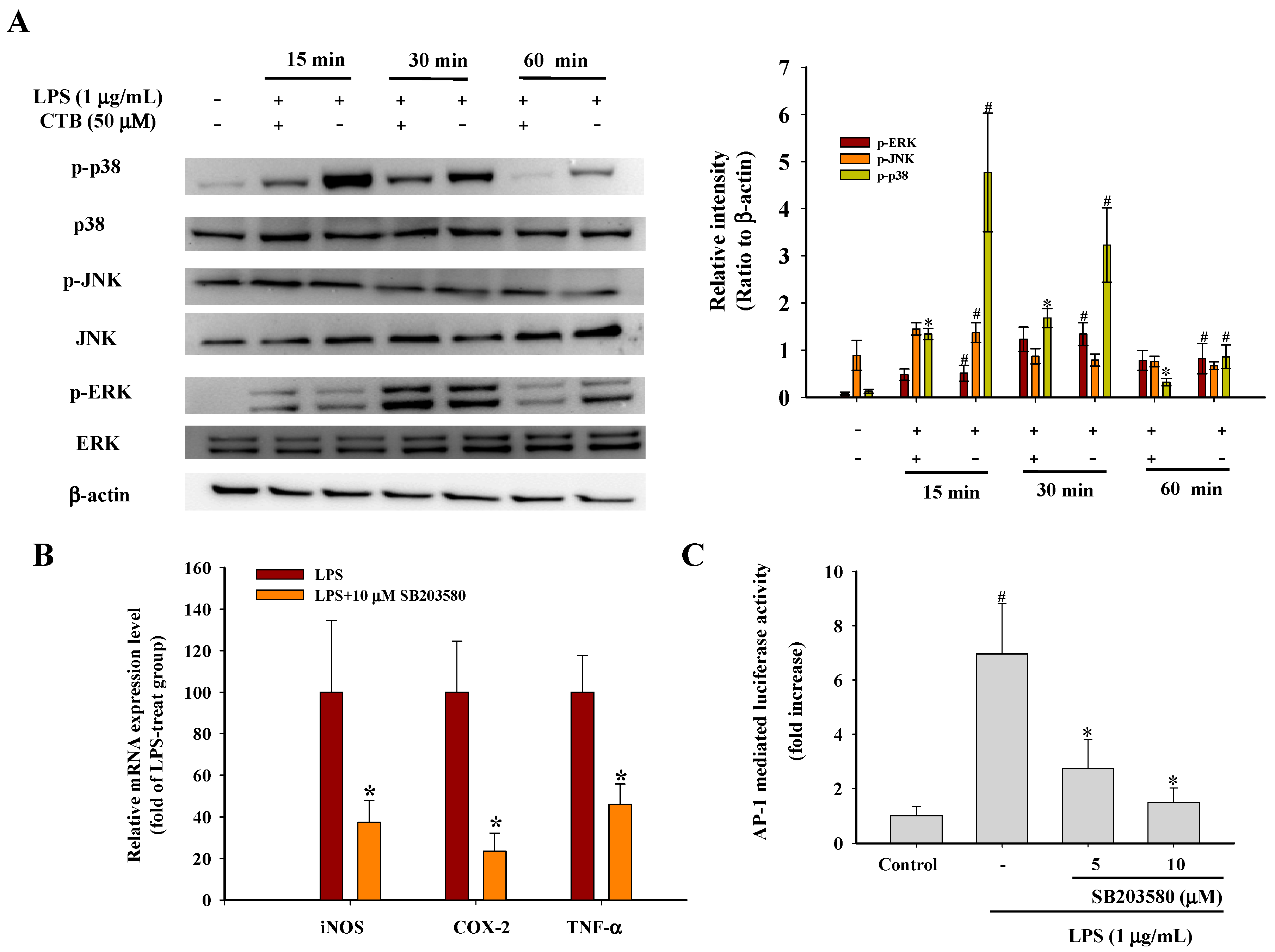
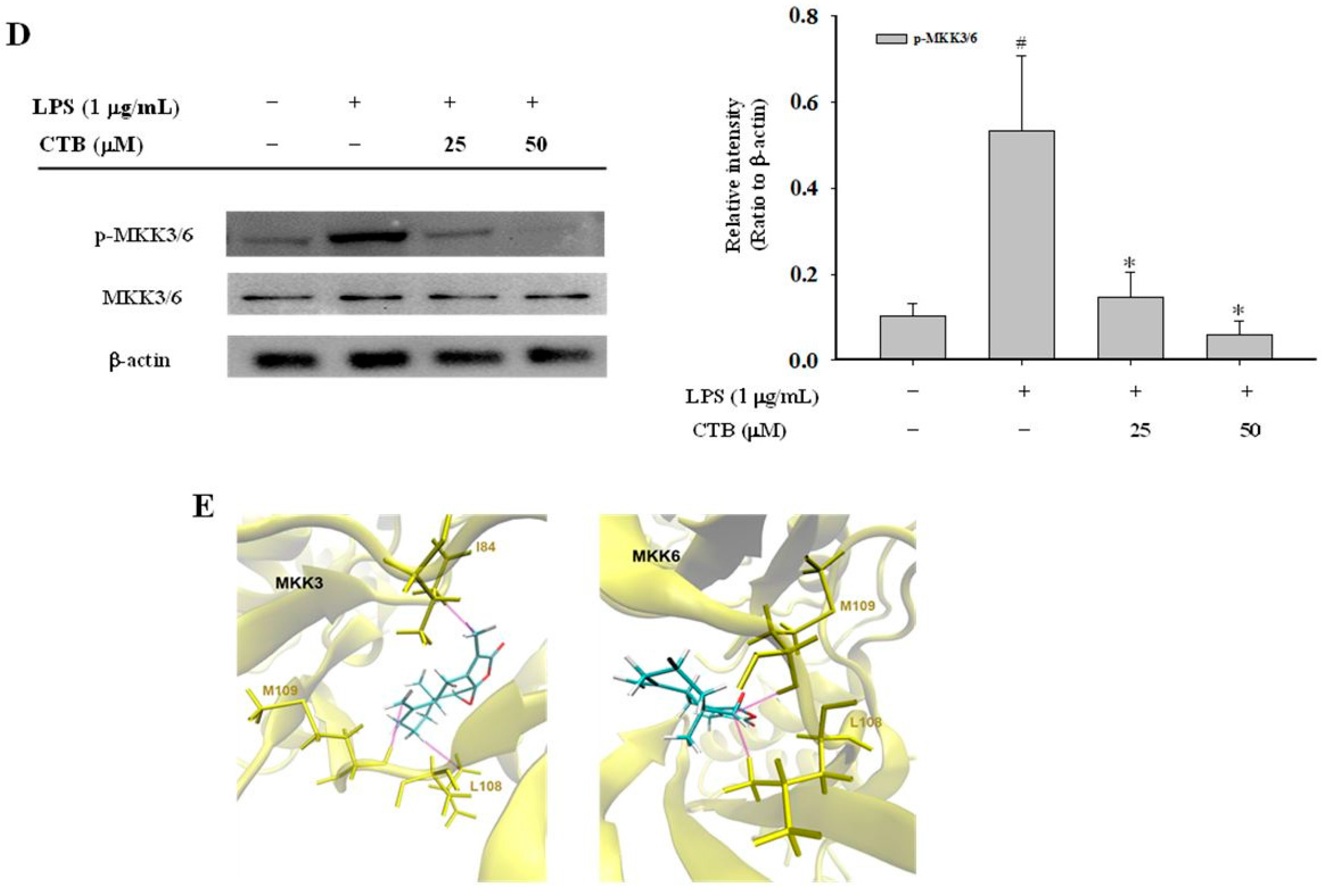
2.4. Effect of CTB on Upstream Signaling of AP-1
3. Materials and Methods
3.1. Plant Material and Chemicals
3.2. General Experimental Procedures
3.3. Extraction, Fractionation, and Isolation
3.4. Cell Line and Cell Culture
3.5. Cell Viability Assay
3.6. Determination of NO, PGE2, TNF-α, IL-6, and IL-1β Production
3.7. RNA Extraction and Reverse-Transcription Polymerase Chain Reaction (RT-PCR)
3.8. Plasmid Transfection and Luciferase Assay
3.9. Western Blot Analysis
3.10. Molecular Docking Study of CTB
3.11. Data Analysis
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| CTB | Chloranthalactone B |
| LPS | Lipopolysaccharide |
| TNF-α | Tumor necrosis factor α |
| IL-1β | Interleukin-1β |
| MAPK | Mitogen-activated protein kinase |
| PRRs | Pattern-recognition receptors |
| TLR4 | Toll-like receptor 4 |
| (NF)-κB | Nuclear factor-κB |
| (IRF)-3 | Interferon regulatory factor-3 |
| (AP)-1 | Activator protein-1 |
| NO | Nitric oxide |
| PGE2 | Prostaglandin E2 |
| MCP-1 | Monocyte chemoattractant protein-1 |
| l-NMA | NG-methyl-l-arginine |
| MTT | 1-(4,5-dimethylthiazol-2-yl)-3,5-diphenylformazan |
References
- Medzhitov, R. Origin and physiological roles of inflammation. Nature 2008, 454, 428–435. [Google Scholar] [CrossRef] [PubMed]
- Tabas, I.; Glass, C.K. Anti-inflammatory therapy in chronic disease: Challenges and opportunities. Science 2013, 339, 166–172. [Google Scholar] [CrossRef] [PubMed]
- Epelman, S.; Liu, P.P.; Mann, D.L. Role of innate and adaptive immune mechanisms in cardiac injury and repair. Nat. Rev. Immunol. 2015, 15, 117–129. [Google Scholar] [CrossRef] [PubMed]
- Qian, C.; Liu, J.; Cao, X. Innate signaling in the inflammatory immune disorders. Cytokine Growth Factor Rev. 2014, 25, 731–738. [Google Scholar] [CrossRef] [PubMed]
- Schindler, S.M.; Little, J.P.; Klegeris, A. Microparticles: A new perspective in central nervous system disorders. BioMed Res. Int. 2014, 2014, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Dyke, T.E.; Winkelhoff, A.J. Infection and inflammatory mechanisms. J. Periodontol. 2013, 40, S1–S7. [Google Scholar] [CrossRef] [PubMed]
- Roy, A.; Srivastava, M.; Saqib, U.; Liu, D.; Faisal, S.M.; Sugathan, S.; Baig, M.S. Potential therapeutic targets for inflammation in toll-like receptor 4 (TLR4)-mediated signaling pathways. Int. Immunopharmacol. 2016, 40, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.C.; Wu, L.; Qiang, Q.; Ji, L.L.; Wang, X.F.; Luo, H.Q.; Shen, T. The dichloromethane fraction from Mahoniabealei (Fort.) Carr. Leaves exerts an anti-inflammatory effect both in vitro and in vivo. J. Ethnopharmacol. 2016, 188, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Kang, Y.X.; Pan, W.; Lei, W.; Feng, B.; Wang, X.J. Enhancement of anti-inflammatory activity of curcumin using phosphatidylserine-containing nanoparticles in cultured macrophages. Int. J. Mol. Sci. 2016, 17, 969. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Li, X.Q.; Wu, H.F.; Long, W.; Jiang, X.; Shen, T.; Hu, W. 5-Methoxyl aesculetin abrogates lipopolysaccharide-induced inflammation by suppressing MAPK and AP-1 pathways in RAW264.7 cells. Int. J. Mol. Sci. 2016, 17, 315. [Google Scholar] [CrossRef] [PubMed]
- Jeong, D.H.; Kim, K.B.; Kim, M.J.; Kang, B.K.; Ahn, D.H. Skipjack tuna (Katsuwonus pelamis) eyeball oil exerts an anti-inflammatory effect by inhibiting NF-κB and MAPK activation in LPS-induced RAW264.7 cells and croton oil-treated mice. Int. Immunopharmacol. 2016, 40, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Qiao, Q.; Li, J.; Wang, W.; Yao, L.P.; Fu, Y.J. Inhibitory effects of geraniin on LPS-induced inflammation via regulating NF-κB and Nrf2 pathways in RAW 264.7 cells. Chem. Biol. Interact. 2016, 253, 134–142. [Google Scholar] [CrossRef] [PubMed]
- Chun, J.; Tosun, A.; Kim, Y.S. Anti-inflammatory effect of corymbocoumarin from Seseli gummiferum subsp. corymbosum through suppression of NF-κB signaling pathway and induction of HO-1 expression in LPS-stimulated RAW264.7 cells. Int. Immunopharmacol. 2016, 31, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Lampiasi, N.; Montana, G. The molecular events behind ferulic acid mediated modulation of IL-6 expression in LPS-activated RAW264.7 cells. Immunobiology 2016, 221, 486–493. [Google Scholar] [CrossRef] [PubMed]
- State Pharmacopeia Committee of China. Chinese Pharmacopoeia; Chinese Medicine Science and Technology Press: Beijing, China, 2010; pp. 207–208. [Google Scholar]
- Liu, T.Y.; Chen, S.B. Sarcandraglabra combined with lycopene protect rats from lipopolysaccharide induced acute lung injury via reducing inflammatory response. Biomed. Pharmacother. 2016, 84, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Masaaki, U.; Genjiro, K.; Yoshikazu, K. Two news sesquiterpenoids from Chloranthus glaber Makino. Heterocycles 1978, 9, 139–144. [Google Scholar]
- Kao, S.Y.; Su, J.H.; Hwang, T.L.; Sheu, J.H.; Su, Y.D.; Lin, C.S.; Chang, Y.C.; Wang, W.H.; Fang, L.S.; Sung, P.J. Discovery of novel sesquiterpenoids from a gorgonian Menella sp. Tetrahedron 2011, 67, 7311–7315. [Google Scholar] [CrossRef]
- Takeda, Y.; Yamashita, H.; Matsumoto, T.; Terao, H. Chloranthalactone F, a sesquiterpenoid from the leaves of Chloranthus glaber. Phytochemistry 1993, 33, 713–715. [Google Scholar] [CrossRef]
- Zheng, W.J.; Wang, S.F.; Chen, X.G.; Hu, Z.D. Analysis of Sarcandraglabra and its medicinal preparations by capillary electrophoresis. Talanta 2003, 60, 955–960. [Google Scholar] [CrossRef]
- Huang, M.J.; Zeng, G.Y.; Tan, J.B.; Li, Y.L.; Tan, G.S.; Zhou, Y.J. Studies on flavonoid glycosdies of Sarcandra glabra. Zhongguo Zhong Yao Za Zhi 2008, 33, 1700–1702. [Google Scholar] [PubMed]
- Xu, X.D.; Hu, X.R.; Yuan, J.Q.; Yang, J.S. Studies on chemical constituents of Sarcandra glabra. Zhongguo Zhong Yao Za Zhi 2008, 33, 900–902. [Google Scholar] [PubMed]
- Li, Y.; Zhang, D.M.; Li, J.B.; Yu, S.S.; Li, Y.; Luo, Y.M. Hepatoprotective sesquiterpene glycosides from Sarcandraglabra. J. Nat. Prod. 2006, 69, 616–620. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.R.; Wu, H.F.; Zhang, X.P.; Yang, J.S.; Dai, Z.; Lin, R.C.; Xu, X.D. A new sesquiterpene lactone from Sarcandra glabra. Nat. Prod. Res. 2013, 27, 1197–1201. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.Y.; Baik, K.U.; Jung, J.H.; Park, M.H. In vitro anti-inflammatory effects of cynaropicrin, a sesquiterpene lactone, from Saussurealappa. Eur. J. Pharmacol. 2000, 398, 399–407. [Google Scholar] [CrossRef]
- Dong, J.; Shao, W.; Yan, P.; Cai, X.; Fang, L.; Zhao, X.; Cai, Y. Curcumolide, a unique sesquiterpenoid with anti-inflammatory properties from Curcumawenyujin. Bioorg. Med. Chem. Lett. 2015, 25, 198–202. [Google Scholar] [CrossRef] [PubMed]
- Kiefer, R.; Kieseier, B.C.; Stoll, G.; Hartung, H.P. The role of macrophages in immune-mediated damage to the peripheral nervous system. Prog. Neurobiol. 2001, 64, 109–127. [Google Scholar] [CrossRef]
- Mantovani, A.; Allavena, P.; Sica, A.; Balkwill, F. Cancer-related inflammation. Nature 2008, 454, 436–444. [Google Scholar] [CrossRef] [PubMed]
- Elinav, E.; Nowarski, R.; Thaiss, C.A.; Hu, B.; Jin, C.; Flavell, R.A. Inflammation-induced cancer: Crosstalk between tumours, immune cells and microorganisms. Nat. Rev. Cancer 2013, 13, 759–771. [Google Scholar] [CrossRef] [PubMed]
- Choi, E.Y.; Kim, H.J.; Han, J.S. Anti-inflammatory effects of calcium citrate in RAW264.7 cells via suppression of NF-κB activation. Environ. Toxicol. Pharmacol. 2015, 39, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, A.; Yaghoobi, M.M.; Najar, A.G.; Kalantari-Khandani, B.; Sharifi, H.; Saravani, M. HSP90 inhibition suppresses PGE2production via modulating COX-2 and 15-PGDH expression in HT-29 colorectal cancer cells. Inflammation 2016, 39, 1116–1123. [Google Scholar] [PubMed]
- Wynn, T.A.; Chawla, A.; Pollard, J.W. Macrophage biology in development, homeostasis and disease. Nature 2013, 496, 445–455. [Google Scholar] [CrossRef] [PubMed]
- Seo, S.; Lee, K.G.; Shin, J.S.; Chung, E.K.; Lee, J.Y.; Kim, H.J.; Lee, K.T. 6′-O-Caffeoyldihydrosyringin isolated from Aster glehni suppresses lipopolysaccharide-induced iNOS, COX-2, TNF-α, IL-1β and IL-6 expression via NF-κB and AP-1 inactivation in RAW264.7 macrophages. Bioorg. Med. Chem. Lett. 2016, 26, 4592–4598. [Google Scholar] [CrossRef] [PubMed]
- Liou, C.J.; Len, W.B.; Wu, S.J.; Lin, C.F.; Wu, X.L.; Huang, W.C. Casticin inhibits COX-2 and iNOS expression via suppression of NF-κB and MAPK signaling in lipopolysaccharide-stimulated mouse macrophages. J. Ethnopharmacol. 2014, 158, 310–316. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Wang, X.; Wu, L.; Shen, T.; Ji, L.; Zhao, X.; Wang, G. Apigenin-7-O-β-d-glucuronide inhibits LPS-induced inflammation through the inactivation of AP-1 and MAPK signaling pathways in RAW264.7 macrophages and protects mice against endotoxin shock. Food Funct. 2016, 7, 1002–1013. [Google Scholar] [CrossRef] [PubMed]
- Kwon, D.J.; Ju, S.M.; Youn, G.S.; Choi, S.Y.; Park, J. Suppression of iNOS and COX-2 expression by flavokawain A via blockade of NF-κB and AP-1 activation in RAW264.7 macrophages. Food Chem. Toxicol. 2013, 58, 479–486. [Google Scholar] [CrossRef] [PubMed]
- Xiao, W.; Yu, A.; Liu, D.; Shen, J.; Xu, Z. Ligustilide treatment promotes functional recovery in a rat model of spinal cord injury via preventing ROS production. Int. J. Clin. Exp. Pathol. 2015, 8, 12005–12013. [Google Scholar] [PubMed]
- Lu, Y.; Suh, S.J.; Kwak, C.H.; Kwon, K.M.; Seo, C.S.; Li, Y.; Jin, Y.; Li, X.; Hwang, S.L.; Kwon, O.; et al. Saucerneol F, a new lignan, inhibits iNOS expression via MAPKs, NF-κB and AP-1 inactivation in LPS-induced RAW264.7 cells. Int. Immunopharmacol. 2012, 12, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Chun, J.; Choi, R.J.; Khan, S.; Lee, D.S.; Kim, Y.C.; Nam, Y.J.; Lee, D.U.; Kim, Y.S. Alantolactone suppresses inducible nitric oxide synthase and cyclooxygenase-2 expression by down-regulating NF-κB, MAPK and AP-1 via the MyD88 signaling pathway in LPS-activated RAW264.7 cells. Int. Immunopharmacol. 2012, 14, 375–383. [Google Scholar] [CrossRef] [PubMed]
- Yeom, M.; Kim, J.H.; Min, J.H.; Hwang, M.K.; Jung, H.S.; Sohn, Y. Xanthii fructus inhibits inflammatory responses in LPS-stimulated RAW264.7 macrophages through suppressing NF-κB and JNK/p38 MAPK. J. Ethnopharmacol. 2015, 24, 394–401. [Google Scholar] [CrossRef] [PubMed]
- Kyriakis, J.M.; Avruch, J. Mammalian MAPK signal transduction pathways activated by stress and inflammation: A 10-year update. Physiol. Rev. 2012, 92, 689–737. [Google Scholar] [CrossRef] [PubMed]
- Xing, J.; Li, R.; Li, N.; Zhang, J.; Li, Y.; Gong, P.; Liu, H.; Zhang, Y. Anti-inflammatory effect of procyanidin B1 on LPS-treated THP1 cells via interaction with the TLR4–MD-2 heterodimer and p38 MAPK and NF-κB signaling. Mol. Cell. Biochem. 2015, 407, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.H.; Sharrocks, A.D.; Whitmarsh, A.J. MAP kinase signalling cascades and transcriptional regulation. Gene 2013, 513, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Intayoung, P.; Limtrakul, P.; Yodkeeree, S. Anti-inflammatory activities of crebanine by inhibition of NF-κB and AP-1 activation through suppressing MAPKs and Aktsignaling in LPS-induced RAW264.7 macrophages. Biol. Pharm. Bull. 2016, 39, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Kang, E.H.; Gebru, E.; Kim, M.H.; Cheng, H.; Park, S.C. EstA protein, a novel virulence factor of Streptococcus pneumoniae, induces nitric oxide and pro-inflammatory cytokine production in RAW264.7 macrophages through NF-κB/MAPK. Microb. Pathog. 2009, 47, 196–201. [Google Scholar] [CrossRef] [PubMed]
- Endale, M.; Park, S.C.; Kim, S.; Kim, S.H.; Yang, Y.; Cho, J.Y.; Rhee, M.H. Quercetin disrupts tyrosine-phosphorylated phosphatidylinositol 3-kinase and myeloid differentiation factor-88 association, and inhibits MAPK/AP-1 and IKK/NF-κB-induced inflammatory mediators production in RAW264.7 cells. Immunobiology 2013, 218, 1452–1467. [Google Scholar] [CrossRef] [PubMed]
- Youn, C.K.; Park, S.J.; Lee, M.Y.; Cha, M.J.; Kim, O.H.; You, H.J.; Chang, I.Y.; Yoon, S.P.; Jeon, Y.J. Silibinin inhibits LPS-induced macrophage activation by blocking p38 MAPK in RAW264.7 cells. Biomol. Ther. 2013, 21, 258–263. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Wang, G.; Li, P.; Wang, Y.; Si, C.L.; He, J.; Wang, X.F. Neuroprotective effects of macranthoin G from Eucommia ulmoides against hydrogen peroxide-induced apoptosis in PC12 cells via inhibiting NF-κB activation. Chem. Biol. Interact. 2014, 224, 108–116. [Google Scholar] [CrossRef] [PubMed]
- Si, C.L.; Shen, T.; Jiang, Y.Y.; Wu, L.; Yu, G.J.; Ren, X.D.; Xu, G.H.; Hu, W.C. Antioxidant properties and neuroprotective effects of isocampneoside II on hydrogen peroxide-induced oxidative injury in PC12 cells. Food Chem. Toxicol. 2013, 59, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Shen, T.; Li, X.; Hu, W.C.; Zhang, L.; Xu, X.; Wu, H.F.; Ji, L.L. Hepatoprotective effect of phenylethanoid glycosides from Incarvillea compacta against CCl4-induced cytotoxicity in HepG2 cells. J. Korean Soc. Appl. Biol. Chem. 2015, 58, 617–625. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | GenBank Accession Number | Primer Sequence (5′–3′) |
|---|---|---|
| GAPDH | NM_001289726 | F: CACTCACGGCAAATTCAACGGCA |
| R: GACTCCACGACATACTCAGCAC | ||
| iNOS | NM_001313921 | F: CCCTTCCGAAGTTTCTGGCAGCAG |
| R: GGCTGTCAGAGCCTCGTGGCTTTGG | ||
| COX-2 | NM_011198 | F: CACTACATCCTGACCCACTT |
| R: ATGCTCCTGCTTGAGTATGT | ||
| TNF-α | NM_013693.3 | F: TGCCTATGTCTCAGCCTCTTC |
| R: GAGGCCATTTGGGAACTTCT | ||
| IL-1β | NM_008361 | F: TGAAGCAGCTATGGCAACTG |
| R: AGGTCAAAGGTTTGGAAGGA |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Shen, J.; Jiang, Y.; Shen, T.; You, L.; Sun, X.; Xu, X.; Hu, W.; Wu, H.; Wang, G. Anti-Inflammatory Effects of Chloranthalactone B in LPS-Stimulated RAW264.7 Cells. Int. J. Mol. Sci. 2016, 17, 1938. https://doi.org/10.3390/ijms17111938
Li X, Shen J, Jiang Y, Shen T, You L, Sun X, Xu X, Hu W, Wu H, Wang G. Anti-Inflammatory Effects of Chloranthalactone B in LPS-Stimulated RAW264.7 Cells. International Journal of Molecular Sciences. 2016; 17(11):1938. https://doi.org/10.3390/ijms17111938
Chicago/Turabian StyleLi, Xueqin, Jun Shen, Yunyao Jiang, Ting Shen, Long You, Xiaobo Sun, Xudong Xu, Weicheng Hu, Haifeng Wu, and Gongcheng Wang. 2016. "Anti-Inflammatory Effects of Chloranthalactone B in LPS-Stimulated RAW264.7 Cells" International Journal of Molecular Sciences 17, no. 11: 1938. https://doi.org/10.3390/ijms17111938
APA StyleLi, X., Shen, J., Jiang, Y., Shen, T., You, L., Sun, X., Xu, X., Hu, W., Wu, H., & Wang, G. (2016). Anti-Inflammatory Effects of Chloranthalactone B in LPS-Stimulated RAW264.7 Cells. International Journal of Molecular Sciences, 17(11), 1938. https://doi.org/10.3390/ijms17111938