
Comparative Evaluation of TRAIL, FGF-2 and VEGF-A-Induced Angiogenesis In Vitro and In Vivo


Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
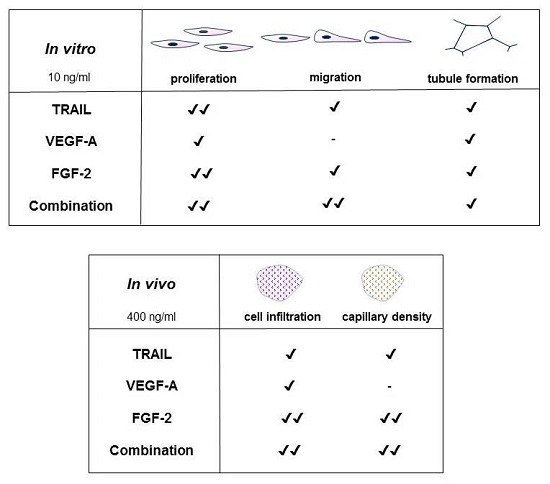
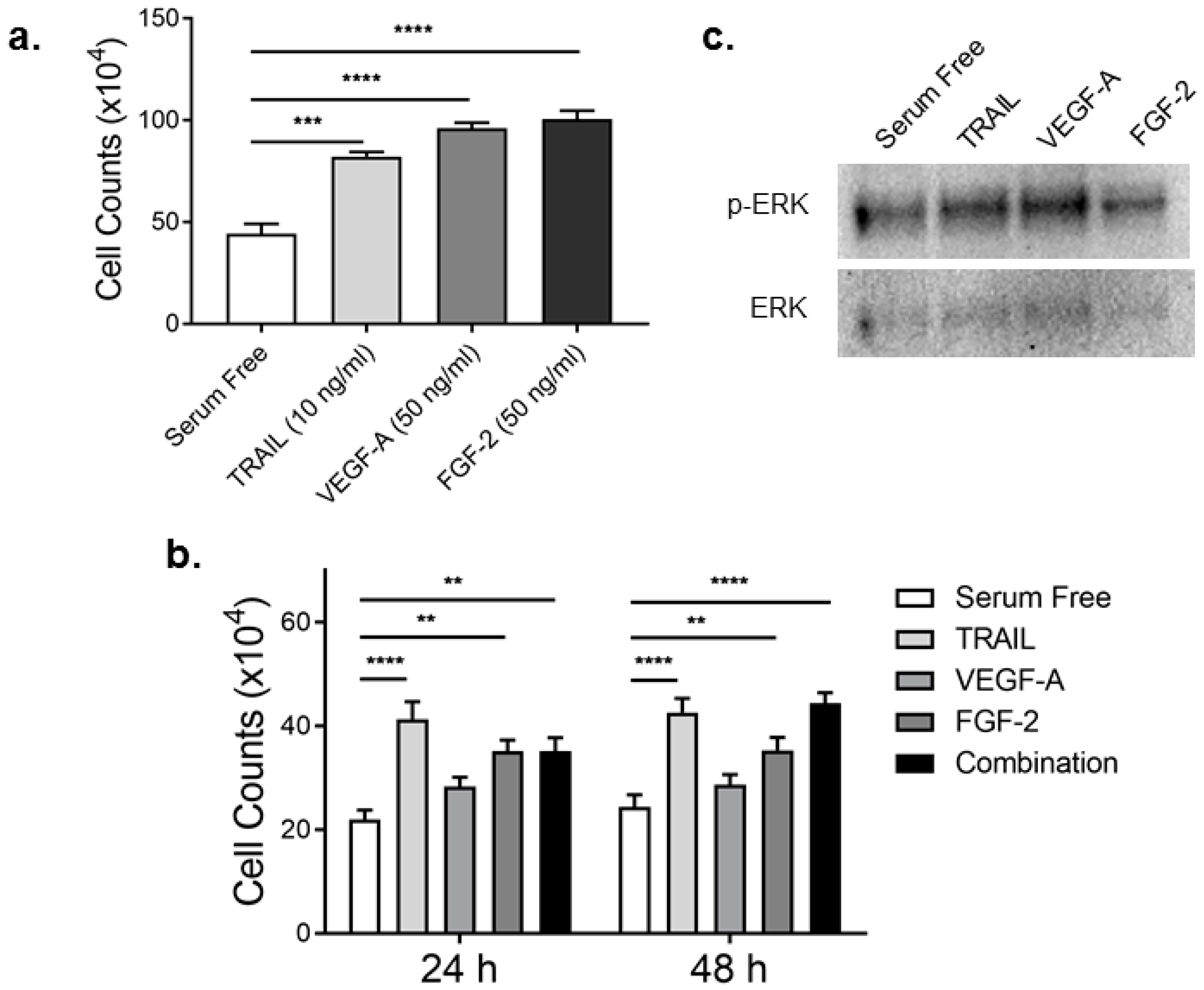
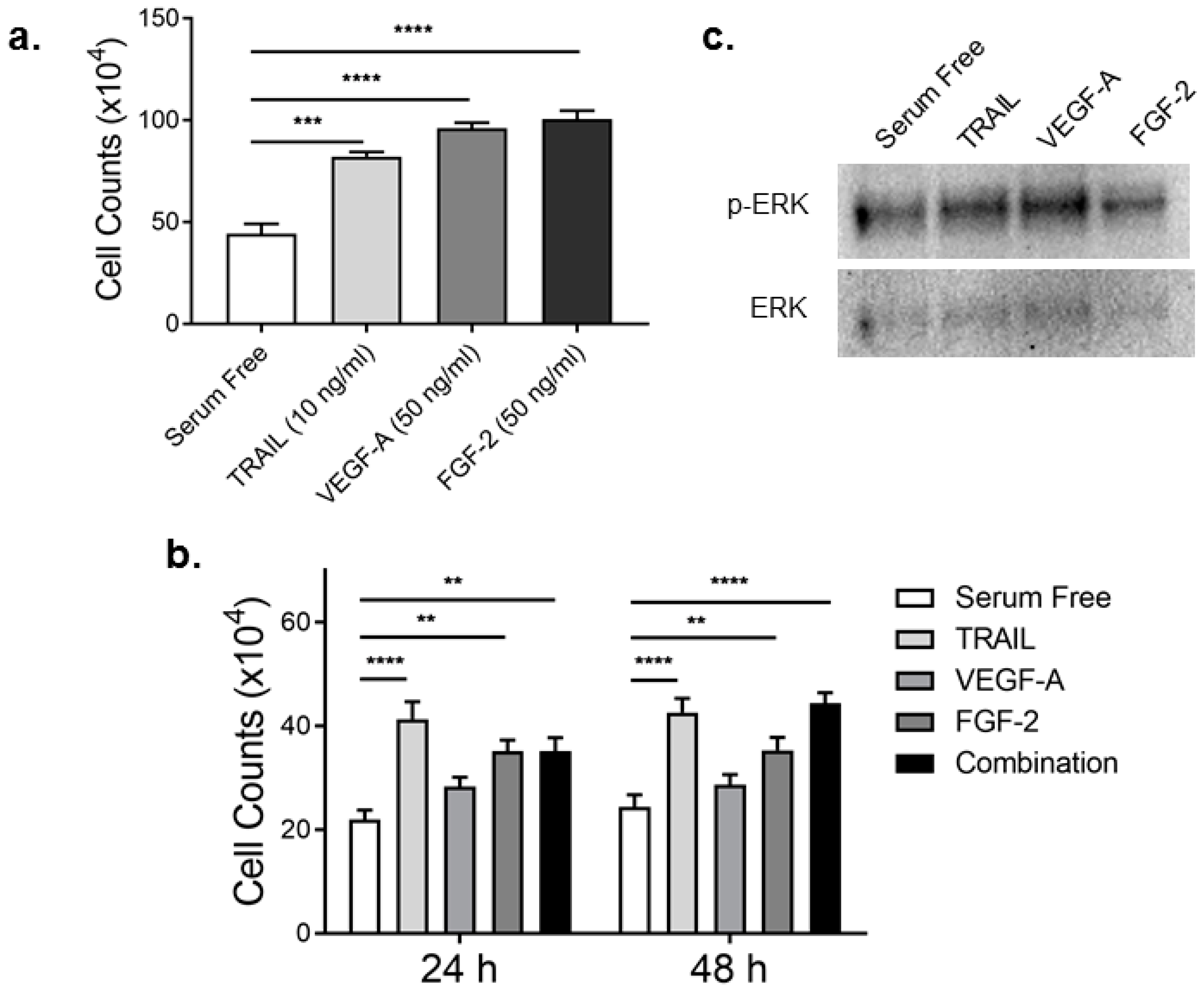
2.1. TRAIL Promotes HMEC-1 Proliferation to the Same Degree as FGF-2
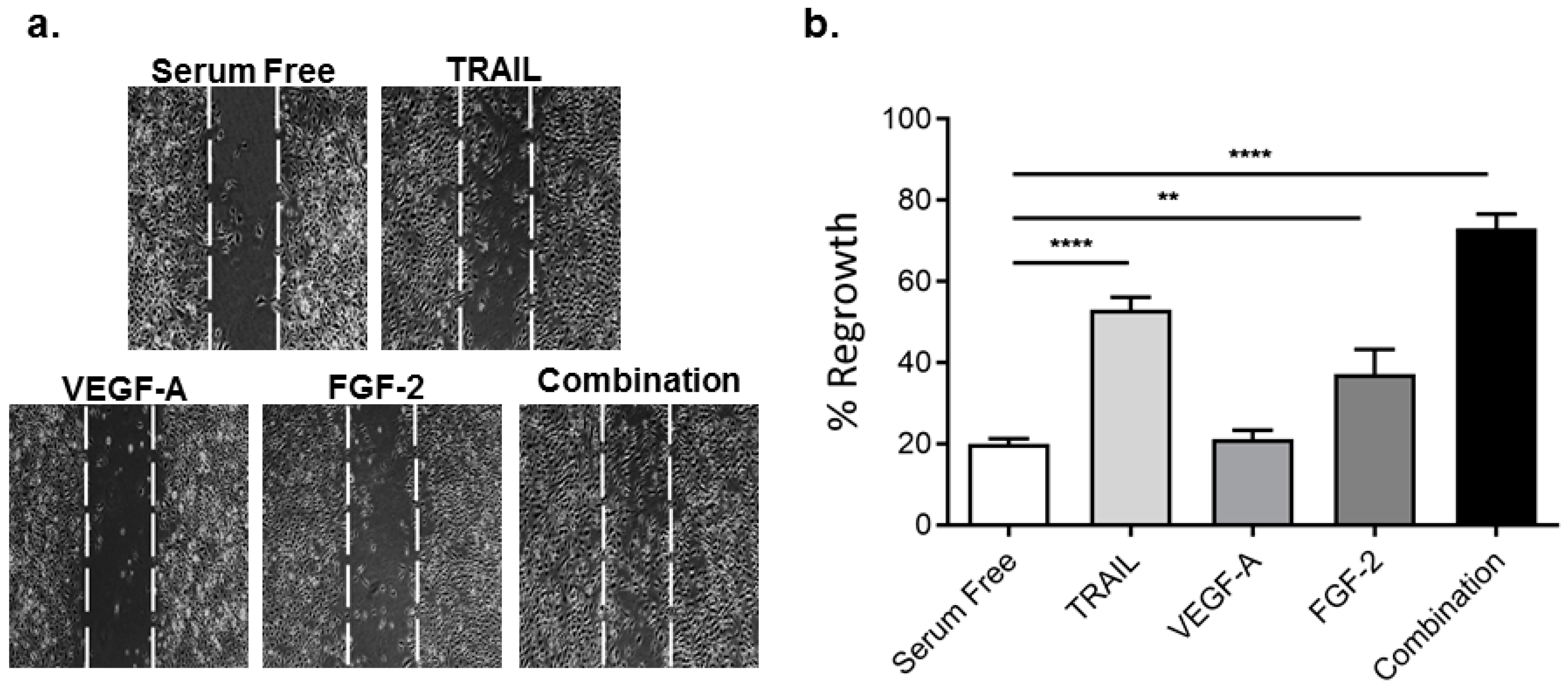
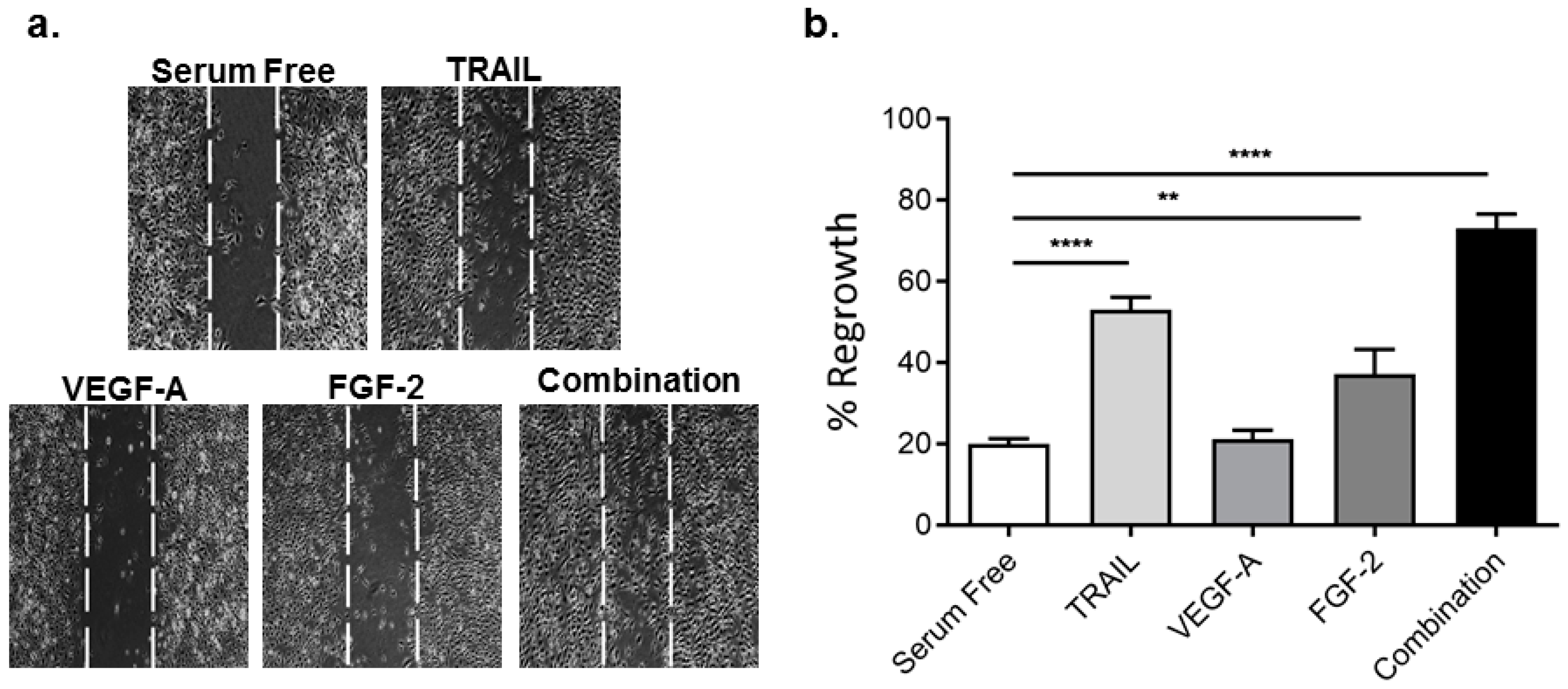
2.2. TRAIL Promotes HMEC-1 Migration More Effectively than VEGF-A and FGF-2, with Combination Treatment Having Additive Effects
2.3. TRAIL Promotes HMEC-1 Tubule Formation More Effectively than VEGF-A, but is Comparable to FGF-2 Treatment
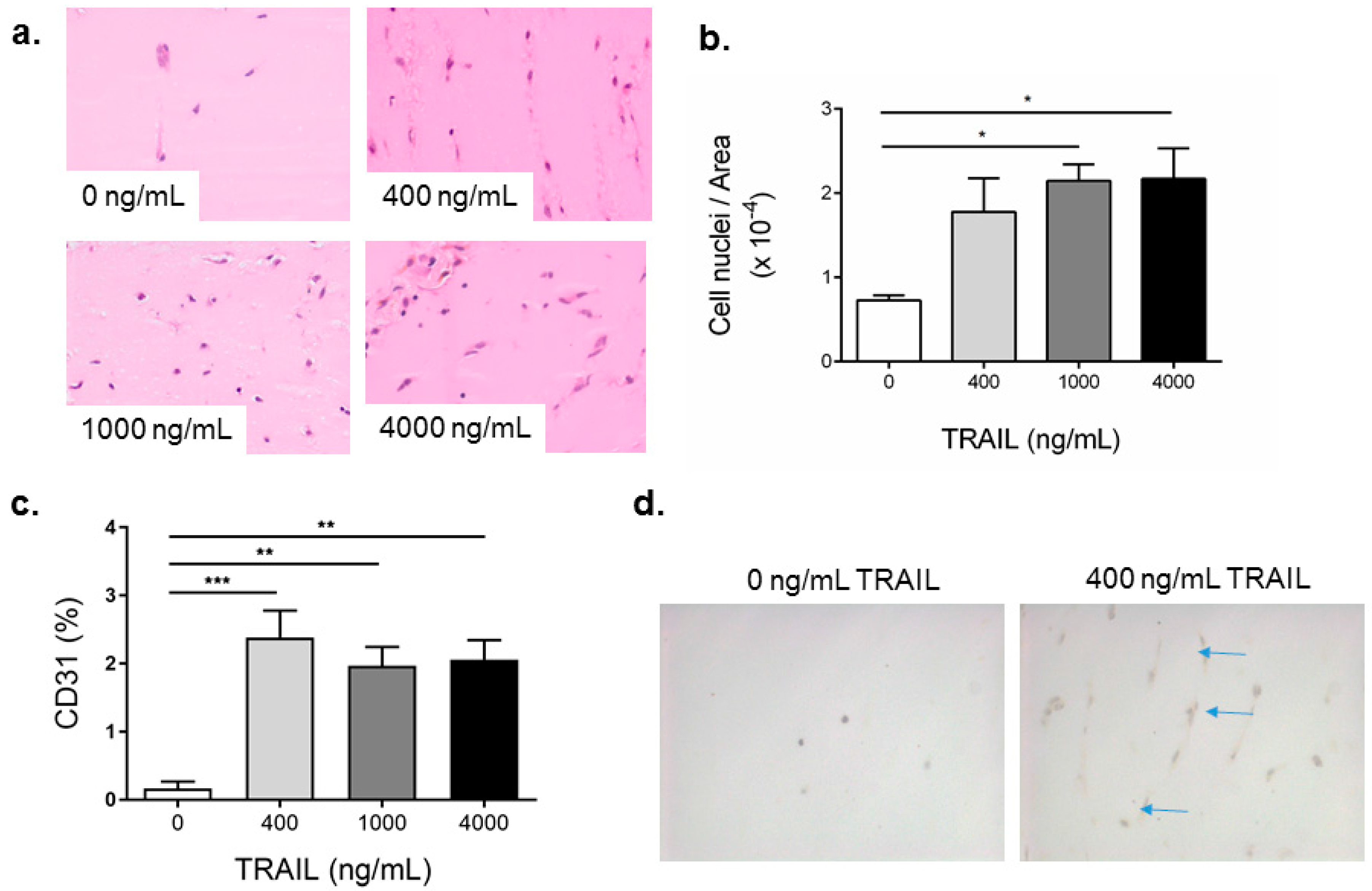
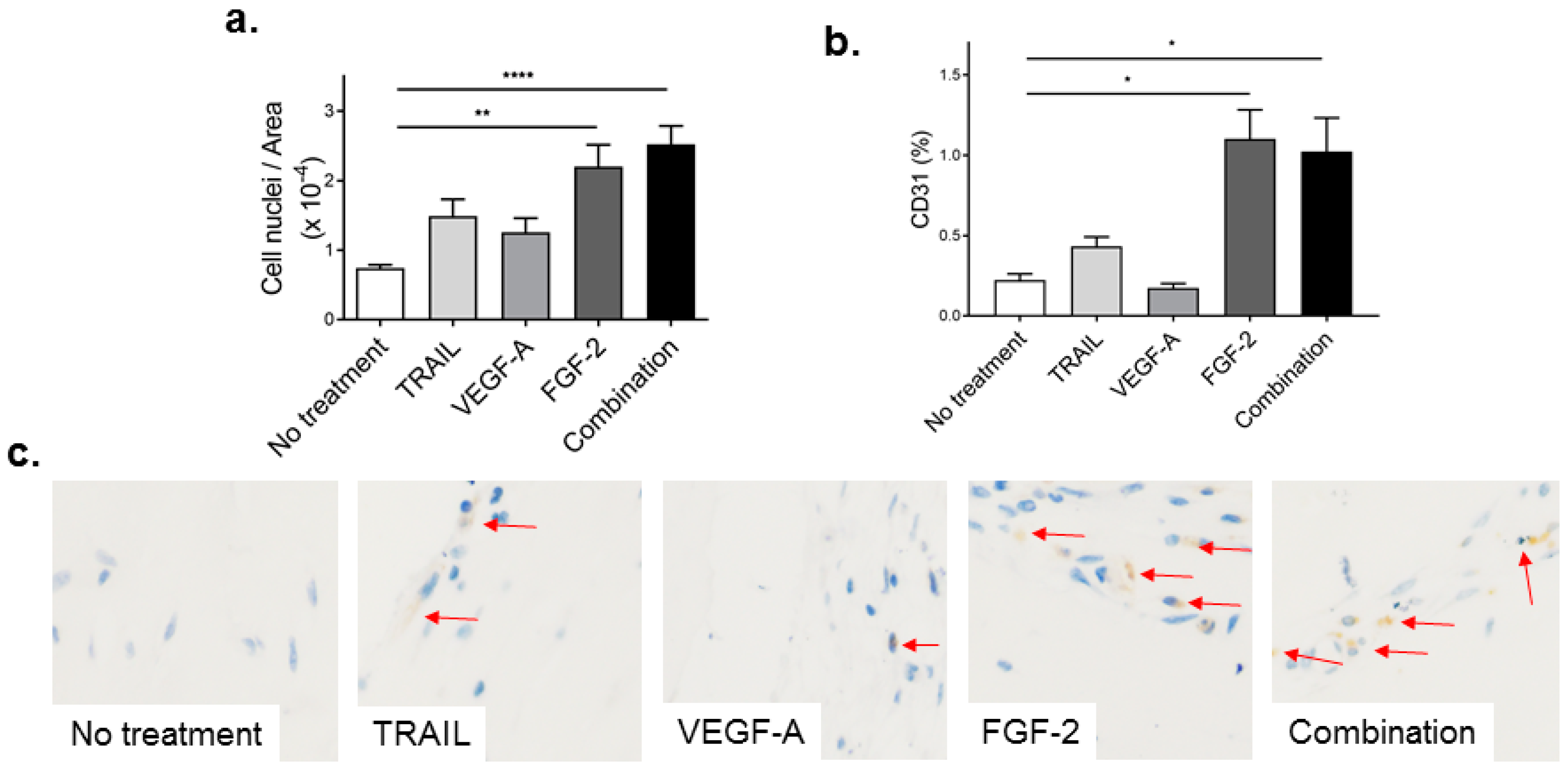
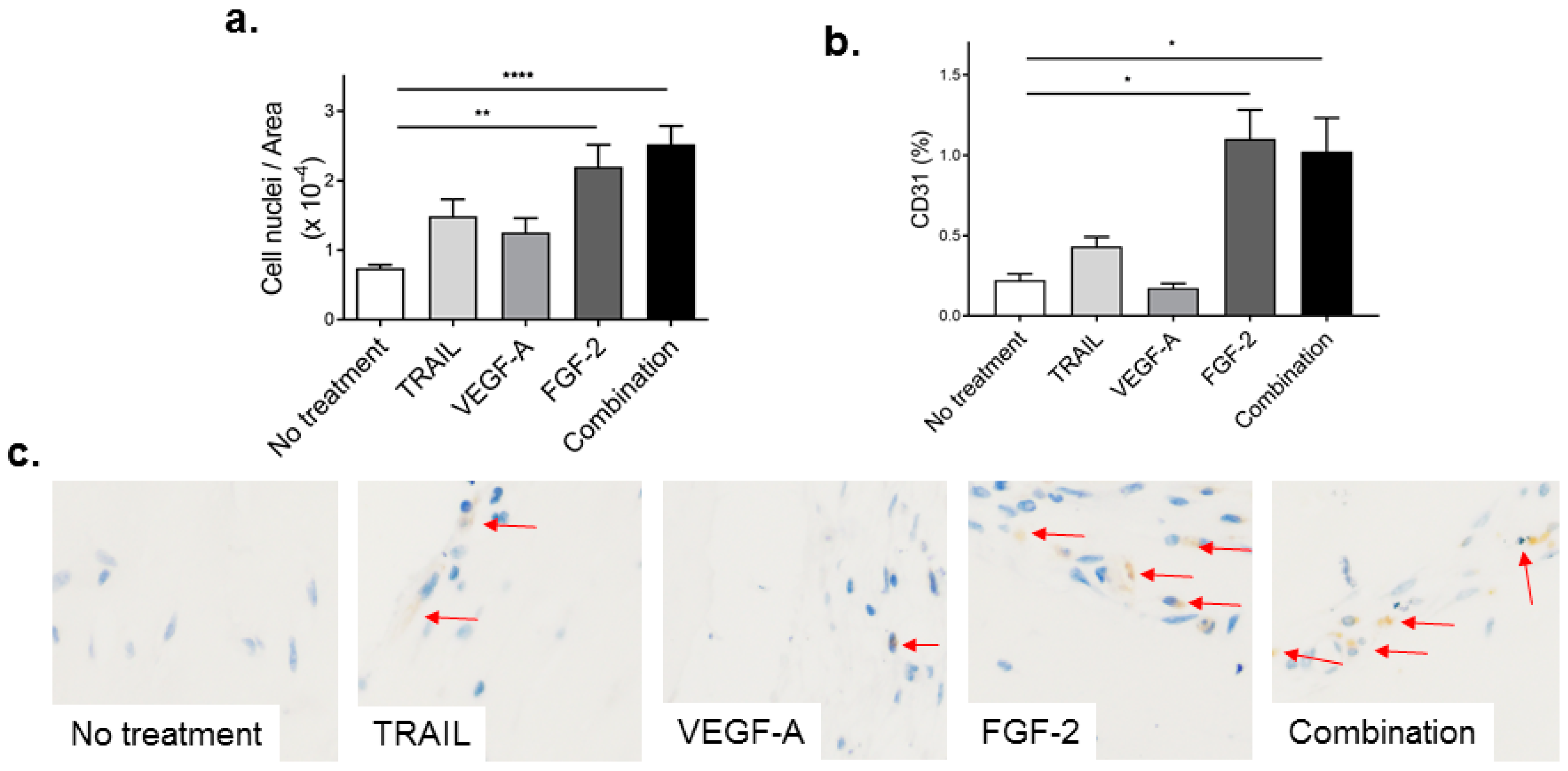
2.4. TRAIL Promotes the Infiltration of Cells and Increases EC Content in Matrigel Plugs
2.5. FGF-2 Increases CD31-Positive Cells in the Matrigel Plug
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Tissue Culture
4.3. Total Cell Counts
4.4. In Vitro Migration Assay
4.5. In Vitro Tubule Assay
4.6. Western Blotting
4.7. Matrigel Plug Assay
4.8. Statistics
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Emanueli, C.; Madeddu, P. Angiogenesis gene therapy to rescue ischaemic tissues: Achievements and future directions. Br. J. Pharmacol. 2001, 133, 951–958. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.L.; Easton, A.S. Evidence that tumor necrosis factor-related apoptosis inducing ligand (TRAIL) inhibits angiogenesis by inducing vascular endothelial cell apoptosis. Biochem. Biophys. Res. Commun. 2010, 391, 936–941. [Google Scholar] [CrossRef] [PubMed]
- Secchiero, P.; Gonelli, A.; Carnevale, E.; Corallini, F.; Rizzardi, C.; Zacchigna, S.; Melato, M.; Zauli, G. Evidence for a proangiogenic activity of TNF-related apoptosis-inducing ligand. Neoplasia 2004, 6, 364–373. [Google Scholar] [CrossRef] [PubMed]
- Di Bartolo, B.A.; Cartland, S.P.; Prado-Lourenco, L.; Griffith, T.S.; Gentile, C.; Ravindran, J.; Azahri, N.S.M.; Thai, T.; Yeung, A.W.S.; Thomas, S.R.; et al. Tumor Necrosis Factor-Related Apoptosis-Inducing Ligand (TRAIL) Promotes Angiogenesis and Ischemia-Induced Neovascularization via NADPH Oxidase 4 (NOX4) and Nitric Oxide—Dependent Mechanisms. J. Am. Heart Assoc. 2015, 4. [Google Scholar] [CrossRef] [PubMed]
- Eilken, H.M.; Adams, R.H. Dynamics of endothelial cell behavior in sprouting angiogenesis. Curr. Opin. Cell Biol. 2010, 22, 617–625. [Google Scholar] [CrossRef] [PubMed]
- Cross, M.J.; Claesson-Welsh, L. FGF and VEGF function in angiogenesis: Signalling pathways, biological responses and therapeutic inhibition. Trends Pharmacol. Sci. 2001, 22, 201–207. [Google Scholar] [CrossRef]
- Staton, C.A.; Reed, M.W.; Brown, N.J. A critical analysis of current in vitro and in vivo angiogenesis assays. Int. J. Exp. Pathol. 2009, 90, 195–221. [Google Scholar] [CrossRef] [PubMed]
- DeCicco-Skinner, K.L.; Gervaise, H.H.; Cataisson, C.; Tabib, T.; Gwilliam, J.C.; Watson, N.J.; Bullwinkle, E.M.; Falkenburg, L.; O’Neill, R.C.; Morin, A.; et al. Endothelial Cell Tube Formation Assay for the In Vitro Study of Angiogenesis. J. Vis. Exp. 2014, 91. [Google Scholar] [CrossRef] [PubMed]
- Ucuzian, A.A.; Gassman, A.A.; East, A.T.; Greisler, H.P. Molecular mediators of angiogenesis. J. Burn Care Res. 2010, 31, 158–175. [Google Scholar] [CrossRef] [PubMed]
- Stahl, A.; Paschek, L.; Martin, G.; Feltgen, N.; Hansen, L.L.; Agostini, H.T. Combinatory inhibition of VEGF and FGF2 is superior to solitary VEGF inhibition in an in vitro model of RPE-induced angiogenesis. Graefes. Arch. Clin. Exp. Ophthalmol. 2009, 247, 767–773. [Google Scholar] [CrossRef] [PubMed]
- Isner, J.M.; Pieczek, A.; Schainfeld, R.; Blair, R.; Haley, L.; Asahara, T.; Rosenfield, K.; Razvi, S.; Walsh, K.; Symes, J.F. Clinical evidence of angiogenesis after arterial gene transfer of phVEGF165 in patient with ischaemic limb. Lancet 1996, 348, 370–374. [Google Scholar] [CrossRef]
- Losordo, D.W.; Vale, P.R.; Hendel, R.C.; Milliken, C.E.; Fortuin, F.D.; Cummings, N.; Schatz, R.A.; Asahara, T.; Isner, J.M.; Kuntz, R.E. Phase 1/2 placebo-controlled, double-blind, dose-escalating trial of myocardial vascular endothelial growth factor 2 gene transfer by catheter delivery in patients with chronic myocardial ischemia. Circulation 2002, 105, 2012–2018. [Google Scholar] [CrossRef] [PubMed]
- Vale, P.R.; Losordo, D.W.; Milliken, C.E.; McDonald, M.C.; Gravelin, L.M.; Curry, C.M.; Esakof, D.D.; Maysky, M.; Symes, J.F.; Isner, J.M. Randomized, single-blind, placebo-controlled pilot study of catheter-based myocardial gene transfer for therapeutic angiogenesis using left ventricular electromechanical mapping in patients with chronic myocardial ischemia. Circulation 2001, 103, 2138–2143. [Google Scholar] [CrossRef] [PubMed]
- Laham, R.J.; Sellke, F.W.; Edelman, E.R.; Pearlman, J.D.; Ware, J.A.; Brown, D.L.; Gold, J.P.; Simons, M. Local perivascular delivery of basic fibroblast growth factor in patients undergoing coronary bypass surgery: Results of a phase I randomized, double-blind, placebo-controlled trial. Circulation 1999, 100, 1865–1871. [Google Scholar] [CrossRef] [PubMed]
- Hamano, K.; Li, T.S.; Kobayashi, T.; Kobayashi, S.; Matsuzaki, M.; Esato, K. Angiogenesis induced by the implantation of self-bone marrow cells: A new material for therapeutic angiogenesis. Cell Transplant. 2000, 9, 439–443. [Google Scholar] [PubMed]
- Tateishi-Yuyama, E.; Matsubara, H.; Murohara, T.; Ikeda, U.; Shintani, S.; Masaki, H.; Amano, K.; Kishimoto, Y.; Yoshimoto, K.; Akashi, H.; et al. Therapeutic angiogenesis for patients with limb ischaemia by autologous transplantation of bone-marrow cells: A pilot study and a randomised controlled trial. Lancet 2002, 360, 427–435. [Google Scholar] [CrossRef]
- Hamano, K.; Nishida, M.; Hirata, K.; Mikamo, A.; Li, T.S.; Harada, M.; Miura, T.; Matsuzaki, M.; Esato, K. Local implantation of autologous bone marrow cells for therapeutic angiogenesis in patients with ischemic heart disease: Clinical trial and preliminary results. Jpn. Circ. J. 2001, 65, 845–847. [Google Scholar] [CrossRef] [PubMed]
- Samant, R.S.; Shevde, L.A. Recent advances in anti-angiogenic therapy of cancer. Oncotarget 2011, 2, 122–134. [Google Scholar] [CrossRef] [PubMed]
- Stewart, M.W. Aflibercept (VEGF Trap-eye): The newest anti-VEGF drug. Br. J. Ophthalmol. 2012, 96, 1157–1158. [Google Scholar] [CrossRef] [PubMed]
- Cantarella, G.; Risuglia, N.; Dell’Eva, R.; Lempereur, L.; Albini, A.; Pennisi, G.; Scoto, G.M.; Noonan, D.N.; Bernardini, R. TRAIL inhibits angiogenesis stimulated by VEGF expression in human glioblastoma cells. Br. J. Cancer 2006, 94, 1428–1435. [Google Scholar] [CrossRef] [PubMed]
- Nanobashvili, J.; Jozkowicz, A.; Neumayer, C.H.; Fügl, A.; Sporn, E.; Polterauer, P.; Huk, I. Comparison of Angiogenic Potential of Human Microvascular Endothelial Cells and Human Umbilical Vein Endothelial Cells. Eur. Surg. 2003, 35, 214–219. [Google Scholar] [CrossRef]
- Smith, E.J.; Staton, C.A. Tubule formation assays. In Angiogenesis Assays: A Critical Appraisal of Current Techniques; Staton, C.A., Lewis, C., Bicknell, R., Eds.; John Wiley & Sons: Chichester, UK, 2007; pp. 65–87. [Google Scholar]
- Kano, M.R.; Morishita, Y.; Iwata, C.; Iwasaka, S.; Watabe, T.; Ouchi, Y.; Miyazono, K.; Miyazawa, K. VEGF-A and FGF-2 synergistically promote neoangiogenesis through enhancement of endogenous PDGF-B–PDGFRβ signaling. J. Cell Sci. 2005, 118, 3759–3768. [Google Scholar] [CrossRef] [PubMed]
- Ferber, S.; Tiram, G.; Satchi-Fainaro, R. Monitoring Functionality and Morphology of Vasculature Recruited by Factors Secreted by Fast-growing Tumor-generating Cells. J. Vis. Exp. 2014, 93. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.-W.; Lien, J.-C.; Kuo, S.-C.; Huang, T.-F. Antiangiogenic mechanisms of PJ-8, a novel inhibitor of vascular endothelial growth factor receptor signaling. Carcinogenesis 2012, 33, 1022–1030. [Google Scholar] [CrossRef] [PubMed]
- Hase, T.; Kawashiri, S.; Tanaka, A.; Nozaki, S.; Noguchi, N.; Kato, K.; Nakaya, H.; Nakagawa, K.; Yamamoto, E. Fibroblast growth factor-2 accelerates invasion of oral squamous cell carcinoma. Int. J. Oral Sci. 2006, 3, 1–9. [Google Scholar] [CrossRef]
- Polnaszek, N.; Kwabi-Addo, B.; Peterson, L.E.; Ozen, M.; Greenberg, N.M.; Ortega, S.; Basilico, C.; Ittmann, M. Fibroblast growth factor 2 promotes tumor progression in an autochthonous mouse model of prostate cancer. Cancer Res. 2003, 63, 5754–5760. [Google Scholar] [PubMed]
- Horstmann, M.; Merseburger, A.S.; von der Heyde, E.; Serth, J.; Wegener, G.; Mengel, M.; Feil, G.; Hennenlotter, J.; Nagele, U.; Anastasiadis, A.; et al. Correlation of bFGF expression in renal cell cancer with clinical and histopathological features by tissue microarray analysis and measurement of serum levels. J. Cancer Res. Clin. Oncol. 2005, 131, 715–722. [Google Scholar] [CrossRef] [PubMed]
- Azahri, N.S.; di Bartolo, B.A.; Khachigian, L.M.; Kavurma, M.M. Sp1, acetylated histone-3 and p300 regulate TRAIL transcription: Mechanisms for PDGF-BB-mediated VSMC proliferation and migration. J. Cell. Biochem. 2012, 13, 2597–2606. [Google Scholar] [CrossRef] [PubMed]
- Kavurma, M.M.; Khachigian, L.M. ERK, JNK and p38 MAP kinase regulate proliferation and migration of phenotypically-distinct SMC subtypes. J. Cell. Biochem. 2003, 89, 289–300. [Google Scholar] [CrossRef] [PubMed]


© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cartland, S.P.; Genner, S.W.; Zahoor, A.; Kavurma, M.M. Comparative Evaluation of TRAIL, FGF-2 and VEGF-A-Induced Angiogenesis In Vitro and In Vivo. Int. J. Mol. Sci. 2016, 17, 2025. https://doi.org/10.3390/ijms17122025
Cartland SP, Genner SW, Zahoor A, Kavurma MM. Comparative Evaluation of TRAIL, FGF-2 and VEGF-A-Induced Angiogenesis In Vitro and In Vivo. International Journal of Molecular Sciences. 2016; 17(12):2025. https://doi.org/10.3390/ijms17122025
Chicago/Turabian StyleCartland, Siân P., Scott W. Genner, Amna Zahoor, and Mary M. Kavurma. 2016. "Comparative Evaluation of TRAIL, FGF-2 and VEGF-A-Induced Angiogenesis In Vitro and In Vivo" International Journal of Molecular Sciences 17, no. 12: 2025. https://doi.org/10.3390/ijms17122025