
Crystal Structure of a Putative Cytochrome P450 Alkane Hydroxylase (CYP153D17) from Sphingomonas sp. PAMC 26605 and Its Conformational Substrate Binding


Abstract
:1. Introduction
2. Results and Discussion
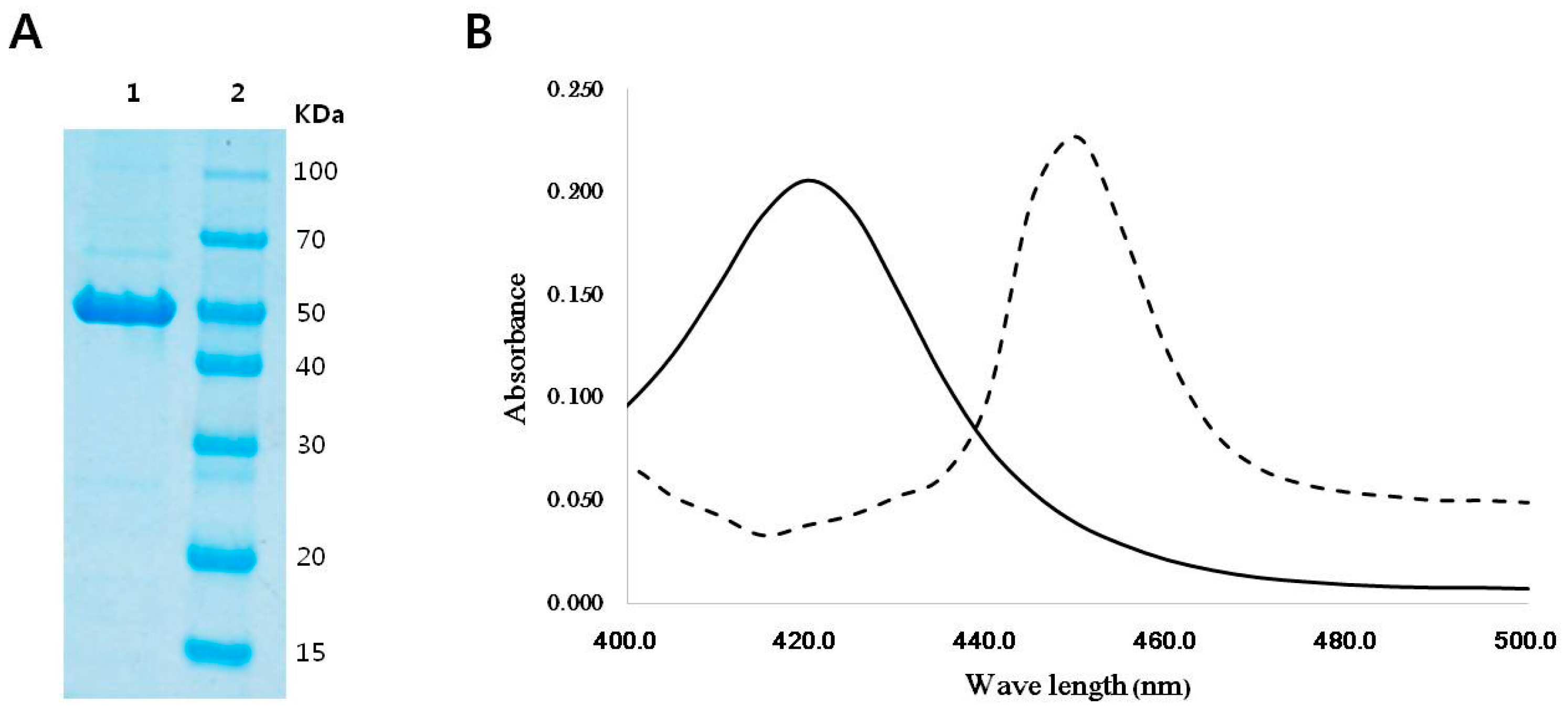
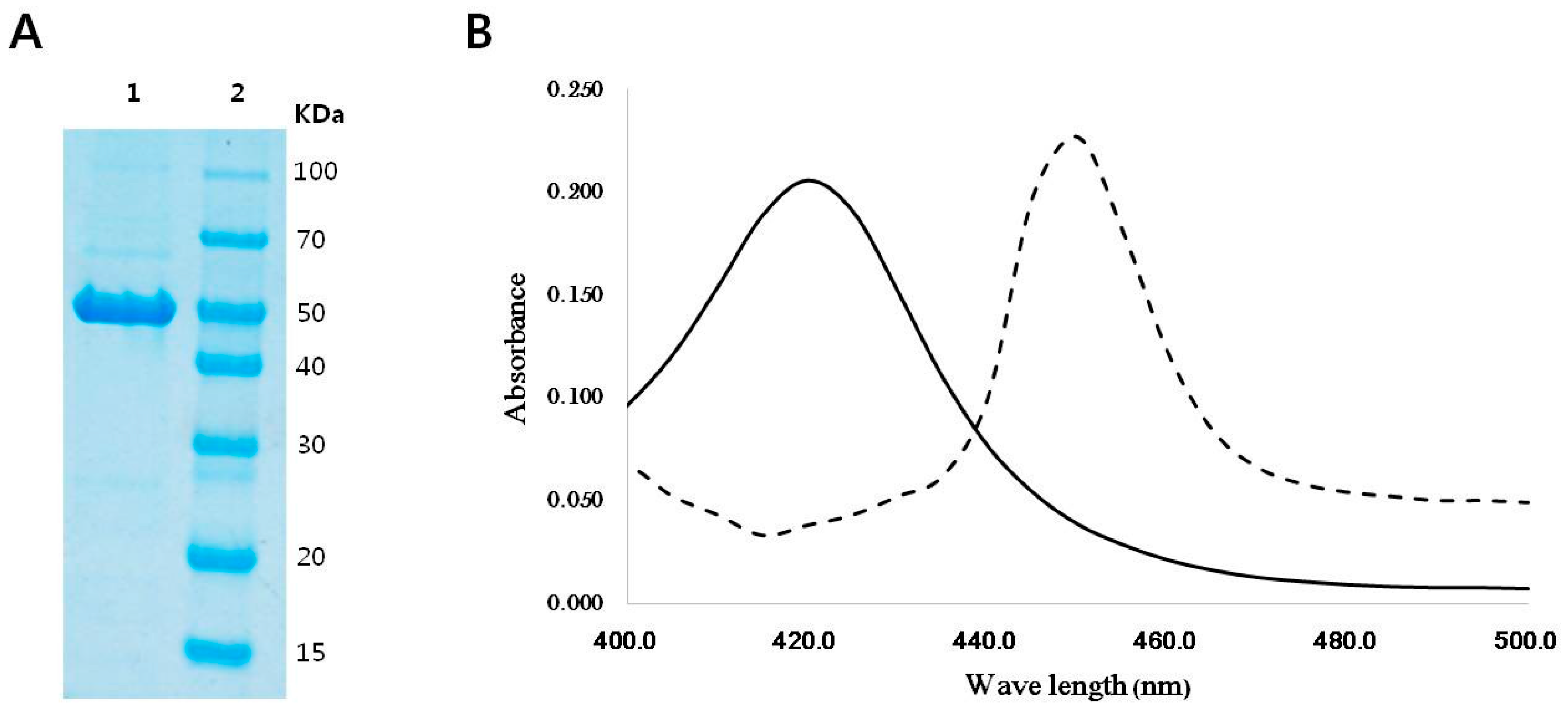
2.1. Sequence Analysis, Purification, and Spectral Analysis of CYP153D17
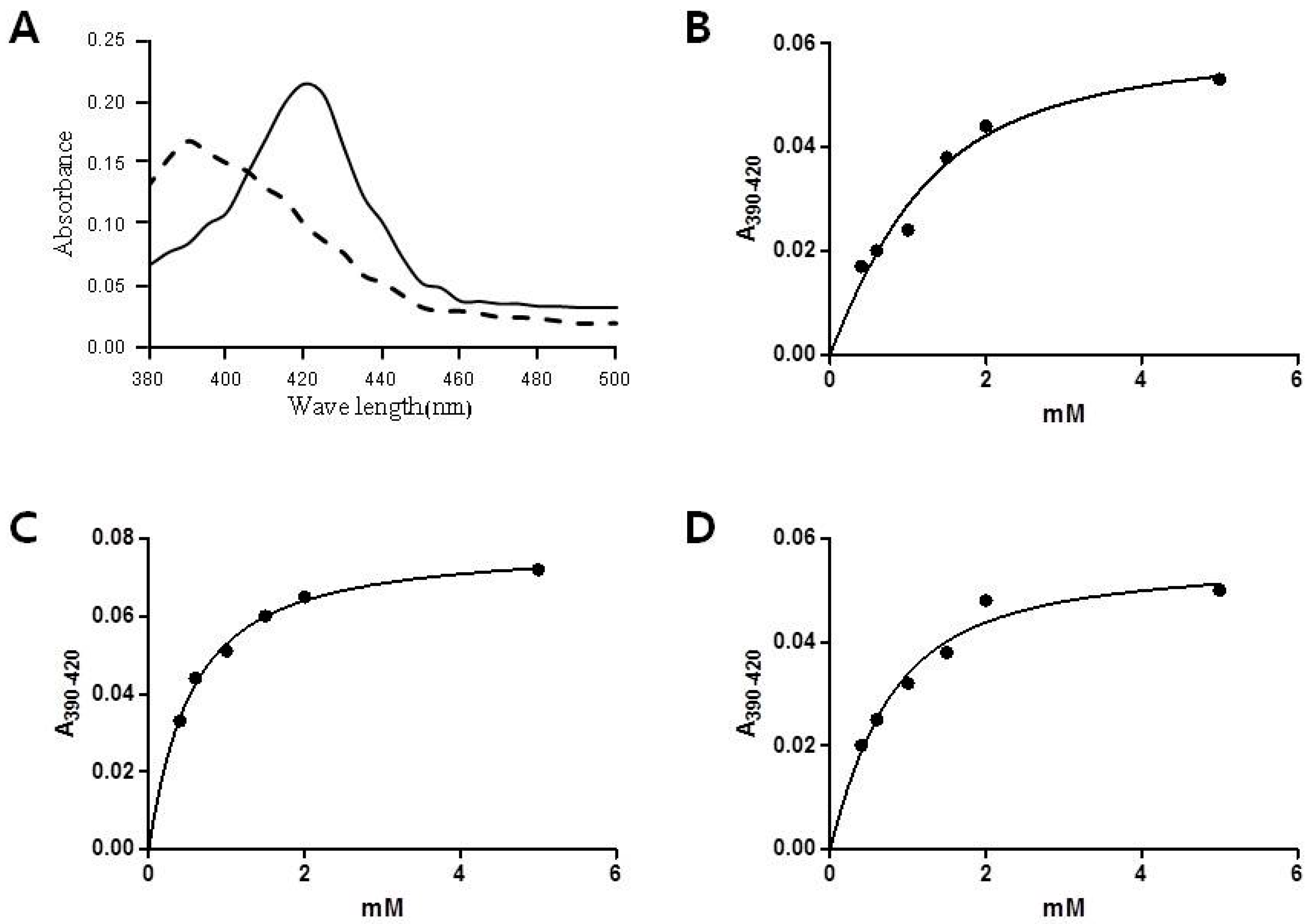
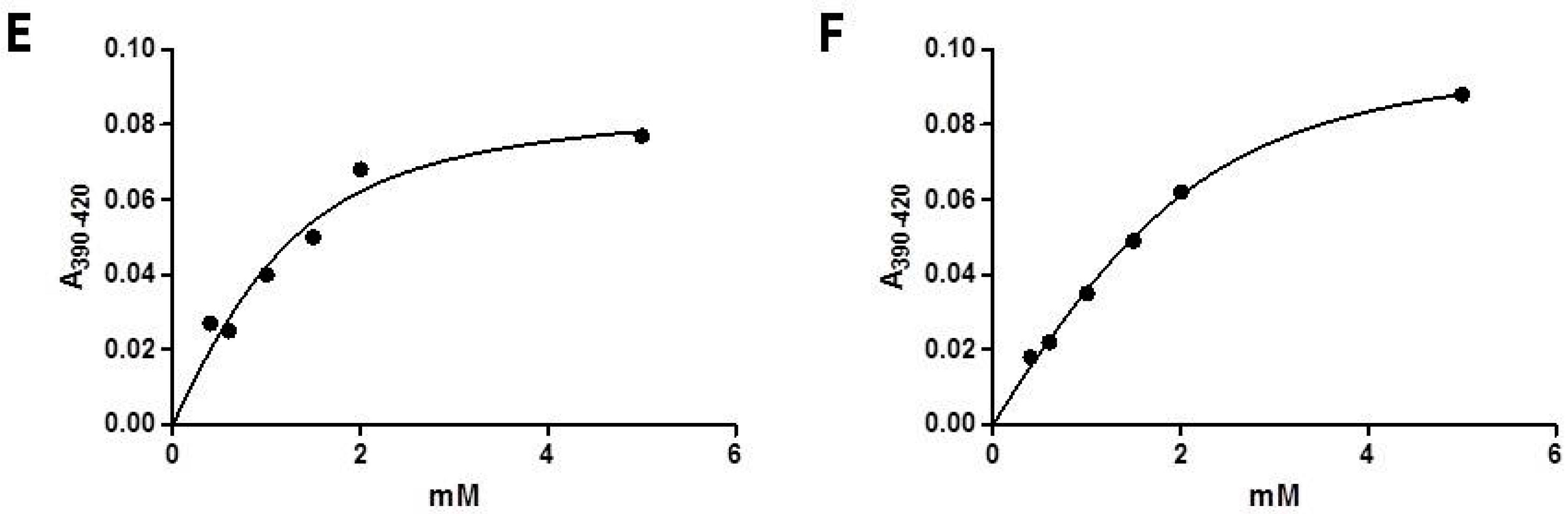
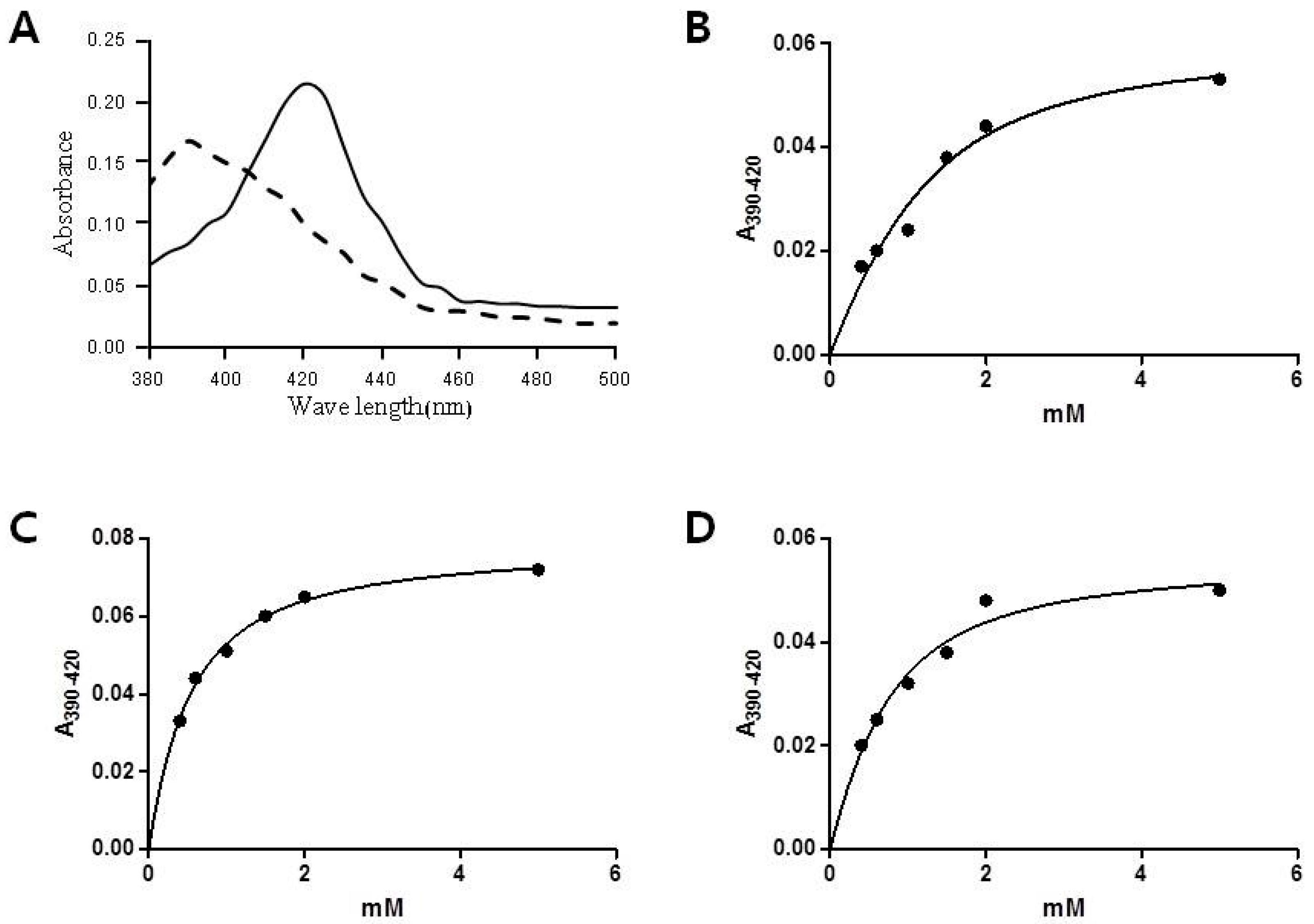
2.2. Substrate Binding Assay of CYP153D17
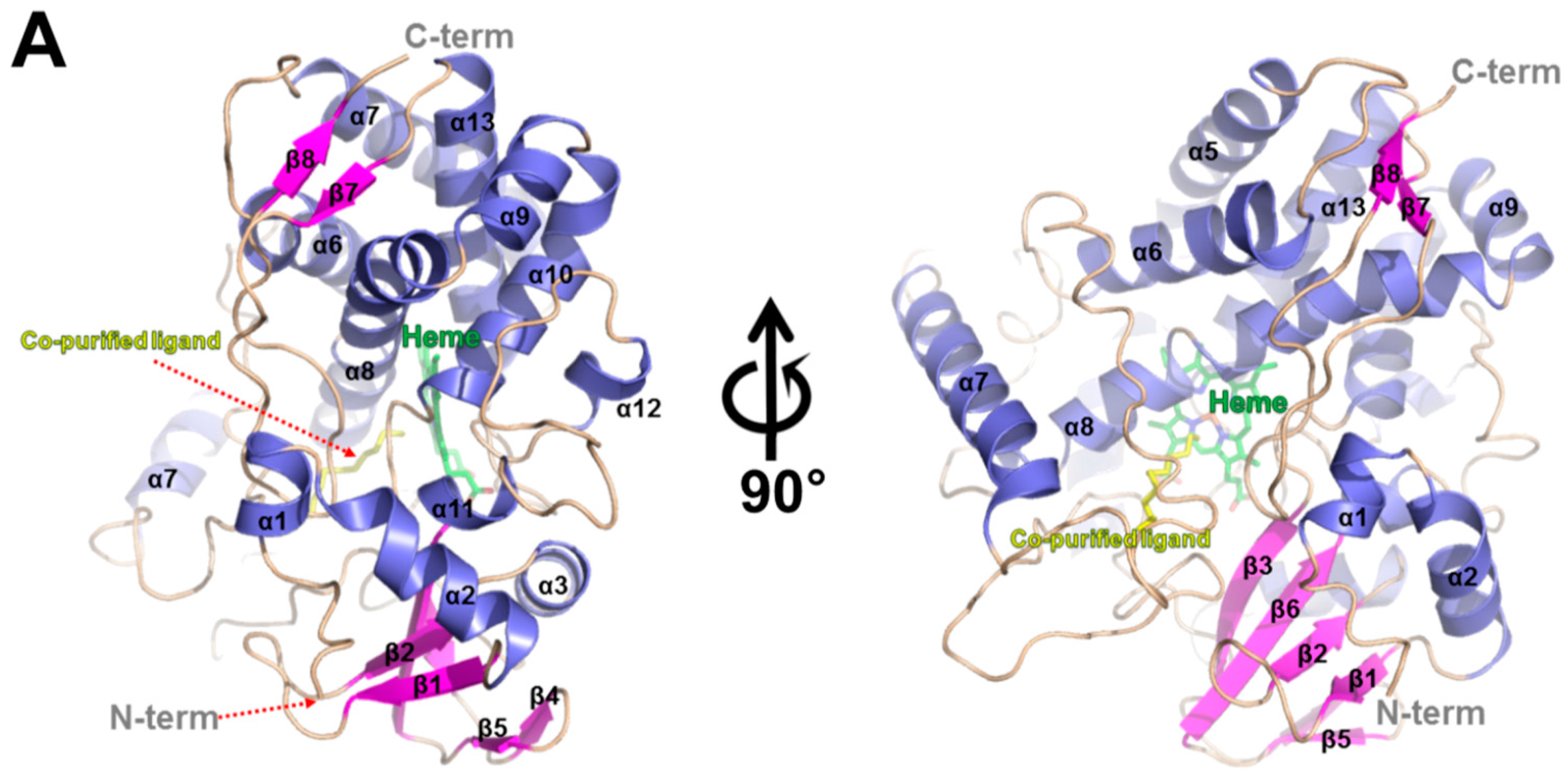
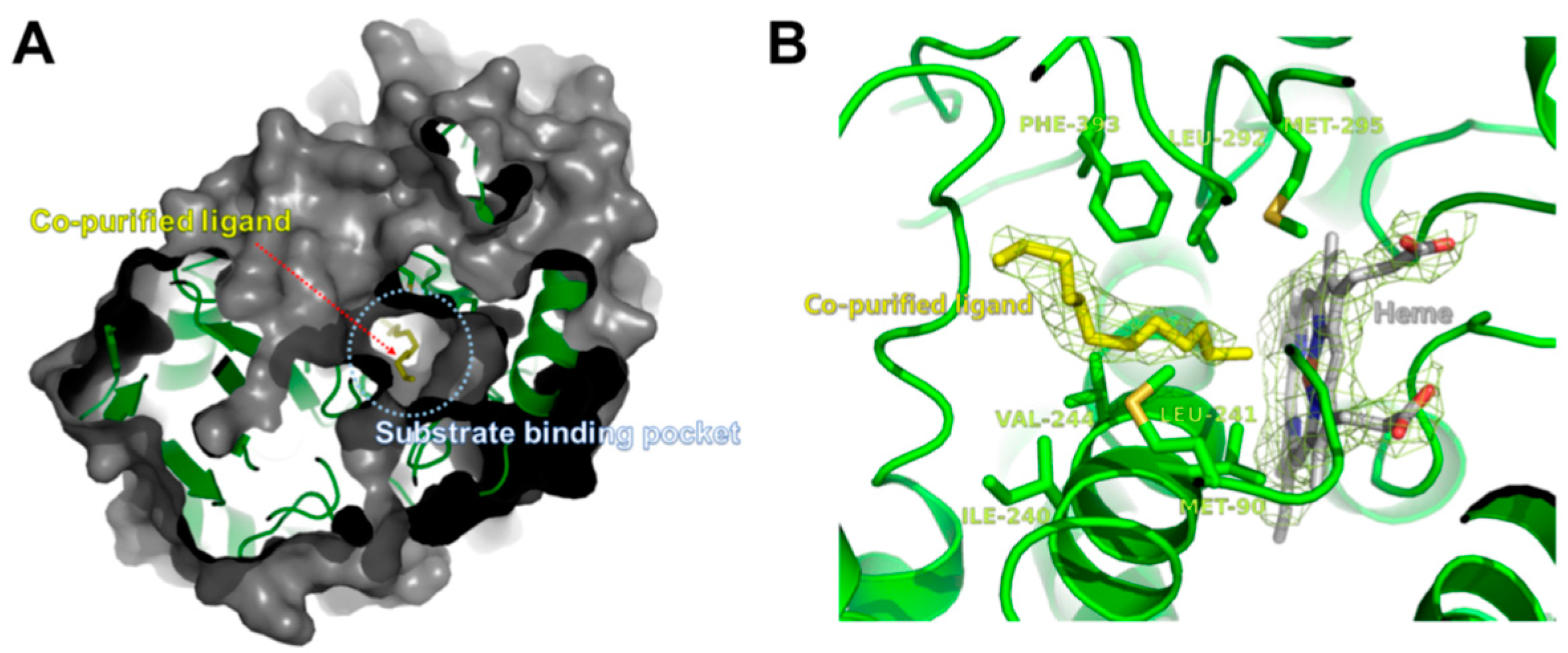
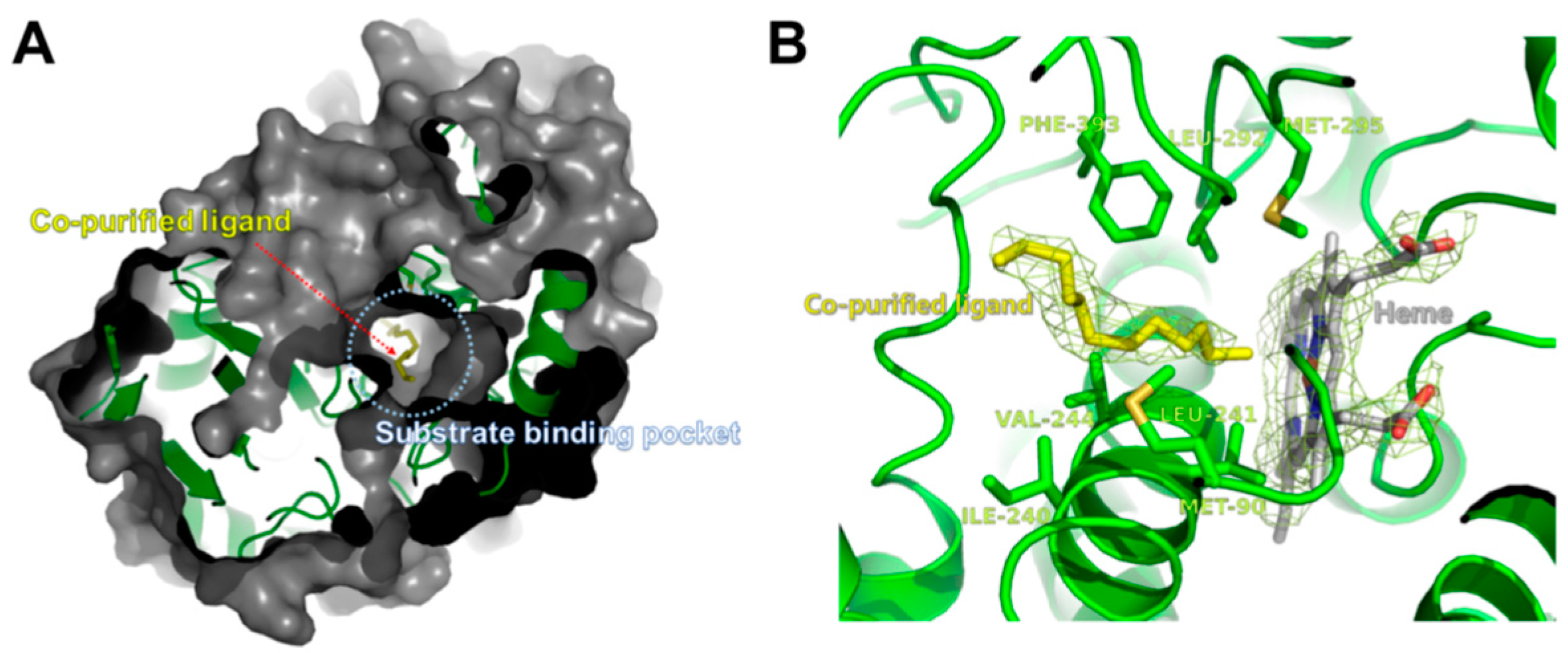
2.3. Overall Structure of CYP153D17
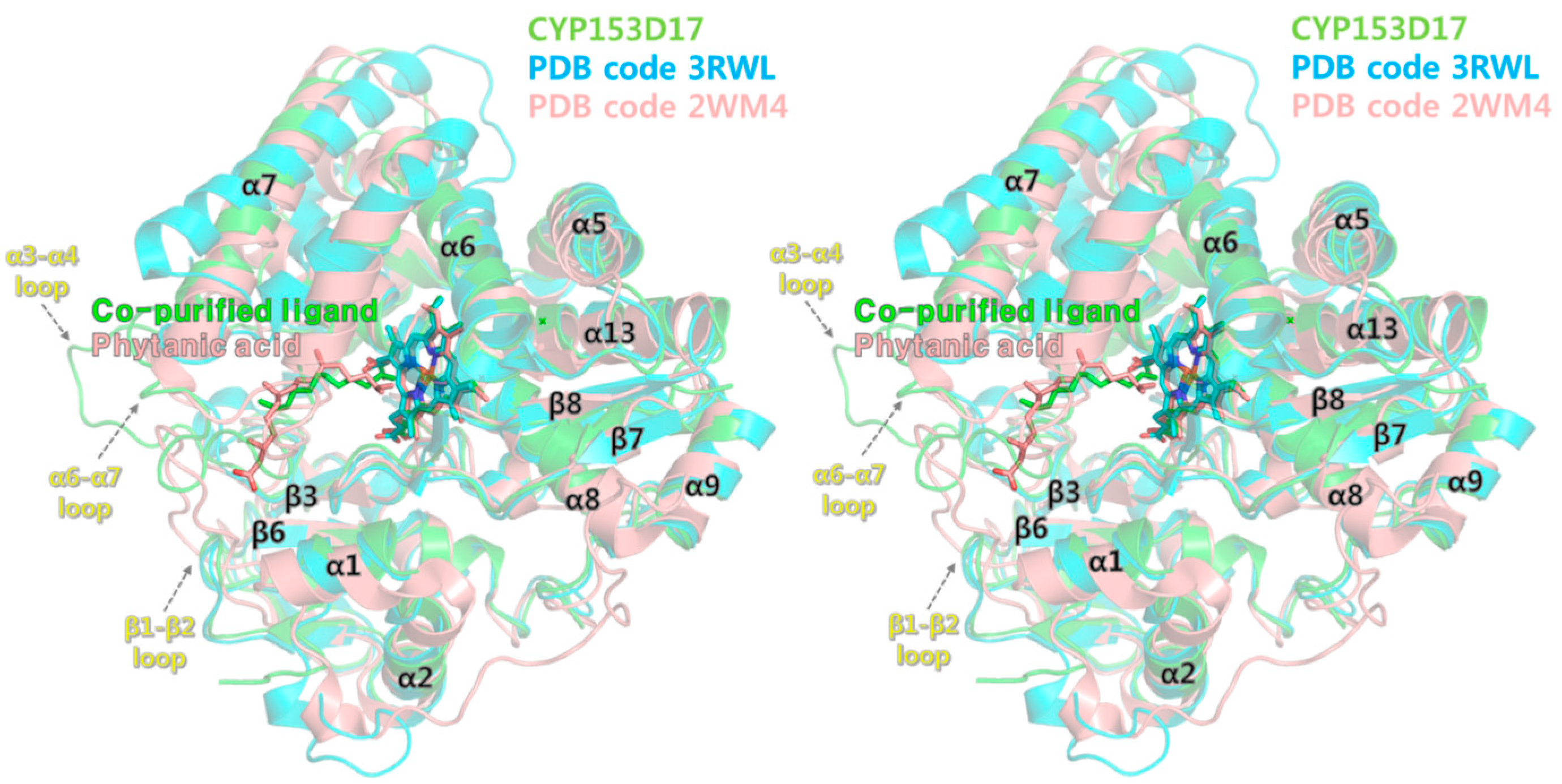
2.4. Structural Comparisons of CYP153D17 with Other CYPs
3. Materials and Methods
3.1. Cloning, Over-Expression, and Purification
3.2. Substrate Binding Assay
3.3. Crystallization and Data Collection
3.4. Structure Determination and Refinement
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Guengerich, F.P. Cytochrome P450 enzymes in the generation of commercial products. Nat. Rev. Drug Discov. 2002, 1, 359–366. [Google Scholar] [CrossRef]
- Ortiz de Montellano, P.R. Cytochrome P450: Structure, Mechanism, and Biochemistry, 3rd ed.; Kluwer Academic/Plenum Publishers: New York, NY, USA, 2005. [Google Scholar]
- Van Beilen, J.B.; Funhoff, E.G.; van Loon, A.; Just, A.; Kaysser, L.; Bouza, M.; Holtackers, R.; Röthlisberger, M.; Li, Z.; Witholt, B. Cytochrome P450 alkane hydroxylases of the CYP153 family are common in alkane-degrading eubacteria lacking integral membrane alkane hydroxylases. Appl. Environ. Microbiol. 2006, 72, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Ji, Y.; Mao, G.; Wang, Y.; Bartlam, M. Structural insights into diversity and n-alkane biodegradation mechanisms of alkane hydroxylases. Front. Microbiol. 2013, 4, 58. [Google Scholar] [CrossRef] [PubMed]
- Nebel, B.A.; Scheps, D.; Honda Malca, S.; Nestl, B.M.; Breuer, M.; Wagner, H.G.; Breitscheidel, B.; Kratz, D.; Hauer, B. Biooxidation of n-butane to 1-butanol by engineered P450 monooxygenase under increased pressure. J. Biotechnol. 2014, 191, 86–92. [Google Scholar] [CrossRef] [PubMed]
- Baer, B.R.; Rettie, A.E. CYP4B1: An enigmatic P450 at the interface between xenobiotic and endobiotic metabolism. Drug Metab. Rev. 2006, 38, 451–476. [Google Scholar] [CrossRef] [PubMed]
- Pinot, F.; Beisson, F. Cytochrome P450 metabolizing fatty acids in plants: Characterization and physiological roles. FEBS J. 2011, 278, 195–205. [Google Scholar] [CrossRef] [PubMed]
- Scheps, D.; Malca, S.H.; Hoffmann, H.; Nestl, B.M.; Hauer, B. Regioselective ω-hydroxylation of medium-chain n-alkanes and primary alcohols by CYP153 enzymes from Mycobacterium marinum and Polaromonas sp. strain JS666. Org. Biomol. Chem. 2011, 9, 6727–6733. [Google Scholar] [CrossRef]
- Hartman, J.H.; Miller, G.P.; Boysen, G. Inhibitory potency of 4-carbon alkanes and alkenes toward CYP2E1 activity. Toxicology 2014, 318, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.C.; Peter, A.; Schwab, W. Expression and characterization of CYP52 genes involved in the biosynthesis of sophorolipid and alkane metabolism from Starmerella bombicola. Appl. Environ. Microbiol. 2014, 80, 766–776. [Google Scholar] [CrossRef] [PubMed]
- Honda Malca, S.; Scheps, D.; Kuhnel, L.; Venegas-Venegas, E.; Seifert, A.; Nestl, B.M.; Hauer, B. Bacterial CYP153A monooxygenases for the synthesis of omega-hydroxylated fatty acids. Chem. Commun. (Camb) 2012, 48, 5115–5117. [Google Scholar] [CrossRef] [PubMed]
- Bell, S.G.; Wong, L.-L. P450 enzymes from the bacterium Novosphingobium aromaticivorans. Biochem. Biophys. Res. Commun. 2007, 360, 666–672. [Google Scholar] [CrossRef] [PubMed]
- Pham, S.Q.; Pompidor, G.; Liu, J.; Li, X.D.; Li, Z. Evolving P450pyr hydroxylase for highly enantioselective hydroxylation at non-activated carbon atom. Chem. Commun. (Camb) 2012, 48, 4618–4620. [Google Scholar] [CrossRef] [PubMed]
- Munday, S.D.; Maddigan, N.K.; Young, R.J.; Bell, S.G. Characterisation of two self-sufficient CYP102 family monooxygenases from Ktedonobacter racemifer DSM44963 which have new fatty acid alcohol product profiles. Biochim. Biophys. Acta 2016, 1860, 1149–1162. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.M.; Snow, C.D.; Vizcarra, C.L.; Mayo, S.L.; Arnold, F.H. Comparison of random mutagenesis and semi-rational designed libraries for improved cytochrome P450 BM3-catalyzed hydroxylation of small alkanes. Protein Eng. Des. Sel. 2012, 25, 171–178. [Google Scholar] [CrossRef]
- Hoffmann, S.M.; Danesh-Azari, H.R.; Spandolf, C.; Weissenborn, M.J.; Grogan, G.; Hauer, B. Structure-guided redesign of CYP153AM. aq for the improved terminal hydroxylation of fatty acids. ChemCatChem 2016, 8, 3234–3239. [Google Scholar] [CrossRef]
- Li, Z.; Feiten, H.J.; Chang, D.; Duetz, W.A.; van Beilen, J.B.; Witholt, B. Preparation of (R)- and (S)-N-protected 3-hydroxypyrrolidines by hydroxylation with Sphingomonas sp. HXN-200, a highly active, regio- and stereoselective, and easy to handle biocatalyst. J. Org. Chem. 2001, 66, 8424–8430. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Li, Z. Evolving P450pyr monooxygenase for regio- and stereoselective hydroxylations. Chimia (Aarau) 2015, 69, 136–141. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.C.; Ahn, D.H.; Lee, J.K.; Kim, S.J.; Hong, S.G.; Kim, E.H.; Park, H. Genome sequence of Sphingomonas sp. strain PAMC 26605, isolated from Arctic lichen (Ochrolechia sp.). J. Bacteriol. 2012, 194, 1607. [Google Scholar] [CrossRef]
- Bhattarai, S.; Liou, K.; Oh, T.-J. Hydroxylation of long chain fatty acids by CYP147F1, a new cytochrome P450 subfamily protein from Streptomyces peucetius. Arch. Biochem. Biophys. 2013, 539, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.W.; Lee, J.-H.; Rimal, H.; Park, H.; Lee, J.H.; Oh, T.-J. Crystal structure of cytochrome P450 (CYP105P2) from Streptomyces peucetius and its conformational changes in response to substrate binding. Int. J. Mol. Sci. 2016, 17, 813. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [PubMed]
- GeneDoc. Available online: http://www.psc.edu/index.php/user-resources/software/genedoc (accessed on 18 March 2015).
- Holm, L.; Sander, C. Dali: A network tool for protein structure comparison. Trends Biochem. Sci. 1995, 20, 478–480. [Google Scholar] [CrossRef]
- Johnston, J.B.; Kells, P.M.; Podust, L.M.; Ortiz de Montellano, P.R. Biochemical and structural characterization of CYP124: A methyl-branched lipid ω-hydroxylase from Mycobacterium tuberculosis. Proc. Natl. Acad. Sci. USA 2009, 106, 20687–20692. [Google Scholar] [CrossRef] [PubMed]
- Driscoll, M.D.; McLean, K.J.; Levy, C.; Mast, N.; Pikuleva, I.A.; Lafite, P.; Rigby, S.E.; Leys, D.; Munro, A.W. Structural and biochemical characterization of Mycobacterium tuberculosis CYP142: Evidence for multiple cholesterol 27-hydroxylase activities in a human pathogen. J. Biol. Chem. 2010, 285, 38270–38282. [Google Scholar] [CrossRef] [PubMed]
- Hasemann, C.A.; Ravichandran, K.G.; Peterson, J.A.; Deisenhofer, J. Crystal structure and refinement of cytochrome P450terp at 2.3 Å resolution. J. Mol. Biol. 1994, 236, 1169–1185. [Google Scholar] [CrossRef]
- Ouellet, H.; Guan, S.; Johnston, J.B.; Chow, E.D.; Kells, P.M.; Burlingame, A.L.; Cox, J.S.; Podust, L.M.; de Montellano, P.R. Mycobacterium tuberculosis CYP125A1, a steroid C27 monooxygenase that detoxifies intracellularly generated cholest-4-en-3-one. Mol. Microbiol. 2010, 77, 730–742. [Google Scholar] [CrossRef] [PubMed]
- Bell, S.G.; Yang, W.; Yorke, J.A.; Zhou, W.; Wang, H.; Harmer, J.; Copley, R.; Zhang, A.; Zhou, R.; Bartlam, M.; et al. Structure and function of CYP108D1 from Novosphingobium aromaticivorans DSM12444: An aromatic hydrocarbon-binding P450 enzyme. Acta Crystallogr. D Biol. Crystallogr. 2012, 68, 277–291. [Google Scholar] [CrossRef] [PubMed]
- Miles, J.; Munro, A.; Rospendowski, B.; Smith, W.; McKnight, J.; Thomson, A. Domains of the catalytically self-sufficient cytochrome P-450 BM-3. Genetic construction, overexpression, purification and spectroscopic characterization. Biochem. J. 1992, 288 Pt 2, 503–509. [Google Scholar] [CrossRef] [PubMed]
- Funhoff, E.G.; Bauer, U.; Garcia-Rubio, I.; Witholt, B.; van Beilen, J.B. CYP153A6, a soluble P450 oxygenase catalyzing terminal-alkane hydroxylation. J. Bacteriol. 2006, 188, 5220–5227. [Google Scholar] [CrossRef] [PubMed]
- Otwinowski, Z.; Minor, W. Processing of X-ray diffraction data. Methods Enzymol. 1997, 276, 307–326. [Google Scholar] [PubMed]
- Vagin, A.; Teplyakov, A. MOLREP: An automated program for molecular replacement. J. Appl. Crystallogr. 1997, 30, 1022–1025. [Google Scholar] [CrossRef]
- Winn, M.D.; Ballard, C.C.; Cowtan, K.D.; Dodson, E.J.; Emsley, P.; Evans, P.R.; Keegan, R.M.; Krissinel, E.B.; Leslie, A.G.; McCoy, A.; et al. Overview of the CCP4 suite and current developments. Acta Crystallogr. D Biol. Crystallogr. 2011, 67, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Emsley, P.; Cowtan, K. Coot: Model-building tools for molecular graphics. Acta Crystallogr. D Biol. Crystallogr. 2004, 60, 2126–2132. [Google Scholar] [CrossRef] [PubMed]
- Murshudov, G.N.; Vagin, A.A.; Dodson, E.J. Refinement of macromolecular structures by the maximum-likelihood method. Acta Crystallogr. D Biol. Crystallogr. 1997, 53, 240–255. [Google Scholar] [CrossRef] [PubMed]
- Chen, V.B.; Arendall, W.B.; Headd, J.J.; Keedy, D.A.; Immormino, R.M.; Kapral, G.J.; Murray, L.W.; Richardson, J.S.; Richardson, D.C. MolProbity: All-atom structure validation for macromolecular crystallography. Acta Crystallogr. D Biol. Crystallogr. 2010, 66, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Vaguine, A.A.; Richelle, J.; Wodak, S. SFCHECK: A unified set of procedures for evaluating the quality of macromolecular structure-factor data and their agreement with the atomic model. Acta Crystallogr. D Biol. Crystallogr. 1999, 55, 191–205. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substrate | Spin Shift (%) | Apparent Kd Value (μM) |
|---|---|---|
| Decane | ≥80 | 0.59 ± 0.69 |
| Dodecane | ≥90 | 0.42 ± 0.14 |
| Capric acid | ≥90 | 0.46 ± 0.41 |
| Lauric acid | ≥90 | 0.49 ± 0.70 |
| Myristic acid | ≥80 | 0.54 ± 0.33 |
| Data Set | CYP153D17 Complexed with Dodecane |
|---|---|
| Data collection | |
| X-ray source | PLS-7A |
| Space group | P3121 |
| Unit-cell parameters (Å) | a = 110.837, b = 110.837, c = 113.18 |
| Wavelength (Å) | 0.97933 |
| Resolution Range (Å) | 50.00–3.10 (3.15–3.10) |
| No. of observed reflections | 62,438 |
| No. of unique reflections | 14,358 |
| Completeness (%) | 94.3 (97.2) |
| Redundancy | 4.4 (4.5) |
| Rmerge a | 0.093 (0.501) |
| I/σ | 26.8 (4.9) |
| Refinement | |
| Resolution range | 50.00–3.10 (3.18–3.10) |
| Reflections: working/free | 13,128/689 (981/43) |
| Rcryst b | 0.261 (0.430) |
| Rfree c | 0.340 (0.463) |
| Ramachandran plot: favored/allowed/disallowed (%) d | 75.1/98.0/22.9 |
| R.m.s.d. bonds (Å) | 0.0126 |
| R.m.s.d. angles (°) | 1.7974 |
| PDB accession code | 5H1Z |
| Protein | PDB Code | DALI Score | Ligand | Sequence % ID with CYP153D17 (Aligned Residue no./Total Residue no.) | Reference |
|---|---|---|---|---|---|
| P450pyr | 3RWL | 47.6 | NL b | 47% (361/404) | [13] |
| CYP124 (Methyl-branched lipid ω-hydroxylase) | 2WM4 | 44.7 | Phytanic acid | 30% (365/426) | [25] |
| CYP142 (Cholesterol oxidase) | 2XKR | 44.6 | Tetraethylene glycol | 28% (359/395) | [26] |
| P450terp | 1CPT | 44.5 | NL | 27% (366/412) | [27] |
| CYP125 | 2X5W | 44.0 | Cholest-4-en-3-one | 27% (366/409) | [28] |
| CYP108D1 | 3TKT | 43.6 | NL b | 29% (367/411) | [29] |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, C.W.; Yu, S.-C.; Lee, J.-H.; Park, S.-H.; Park, H.; Oh, T.-J.; Lee, J.H. Crystal Structure of a Putative Cytochrome P450 Alkane Hydroxylase (CYP153D17) from Sphingomonas sp. PAMC 26605 and Its Conformational Substrate Binding. Int. J. Mol. Sci. 2016, 17, 2067. https://doi.org/10.3390/ijms17122067
Lee CW, Yu S-C, Lee J-H, Park S-H, Park H, Oh T-J, Lee JH. Crystal Structure of a Putative Cytochrome P450 Alkane Hydroxylase (CYP153D17) from Sphingomonas sp. PAMC 26605 and Its Conformational Substrate Binding. International Journal of Molecular Sciences. 2016; 17(12):2067. https://doi.org/10.3390/ijms17122067
Chicago/Turabian StyleLee, Chang Woo, Sang-Cheol Yu, Joo-Ho Lee, Sun-Ha Park, Hyun Park, Tae-Jin Oh, and Jun Hyuck Lee. 2016. "Crystal Structure of a Putative Cytochrome P450 Alkane Hydroxylase (CYP153D17) from Sphingomonas sp. PAMC 26605 and Its Conformational Substrate Binding" International Journal of Molecular Sciences 17, no. 12: 2067. https://doi.org/10.3390/ijms17122067
APA StyleLee, C. W., Yu, S.-C., Lee, J.-H., Park, S.-H., Park, H., Oh, T.-J., & Lee, J. H. (2016). Crystal Structure of a Putative Cytochrome P450 Alkane Hydroxylase (CYP153D17) from Sphingomonas sp. PAMC 26605 and Its Conformational Substrate Binding. International Journal of Molecular Sciences, 17(12), 2067. https://doi.org/10.3390/ijms17122067