
Pioglitazone, a Peroxisome Proliferator-Activated Receptor γ Agonist, Suppresses Rat Prostate Carcinogenesis

 ,
, 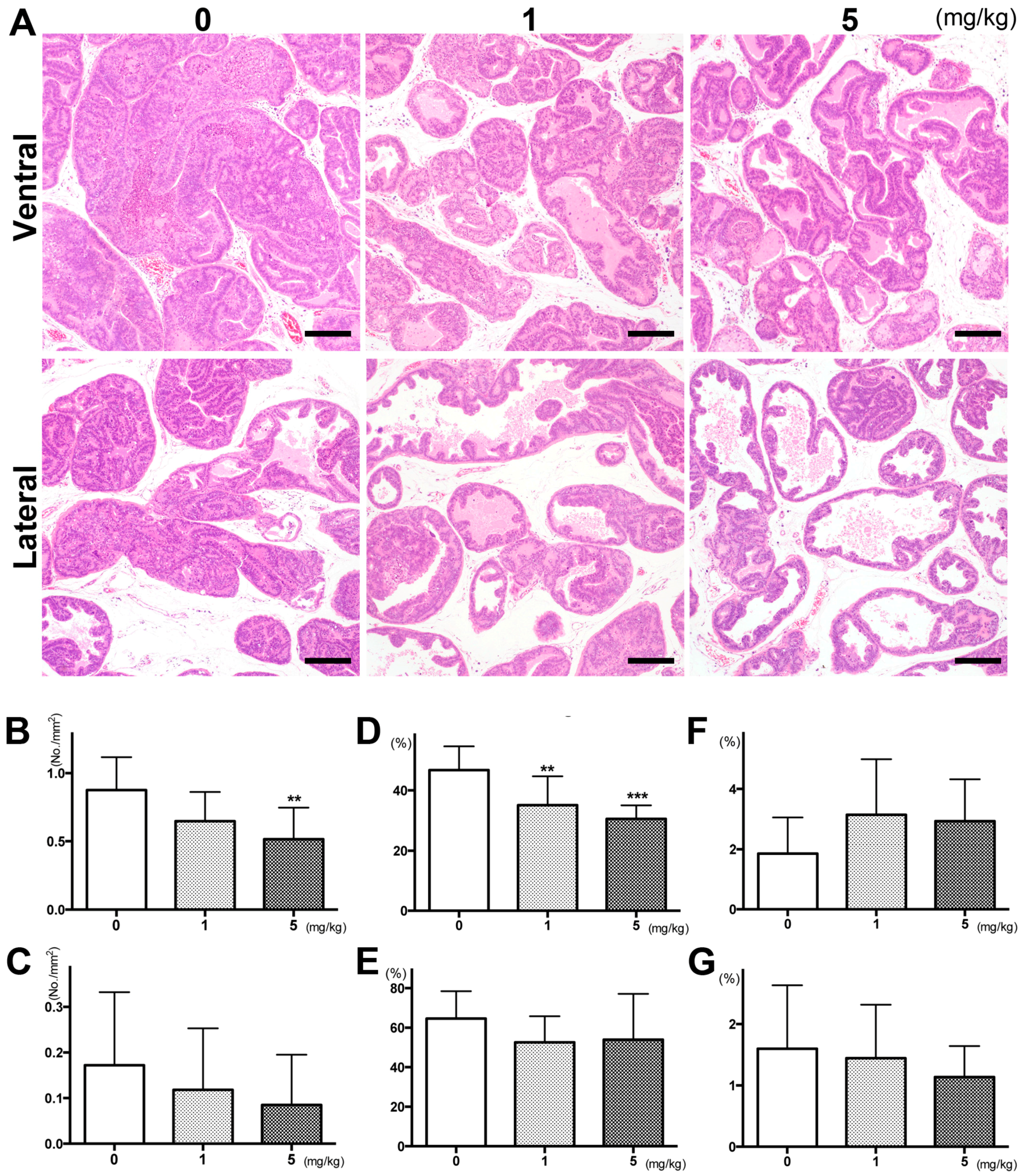{kind=link}
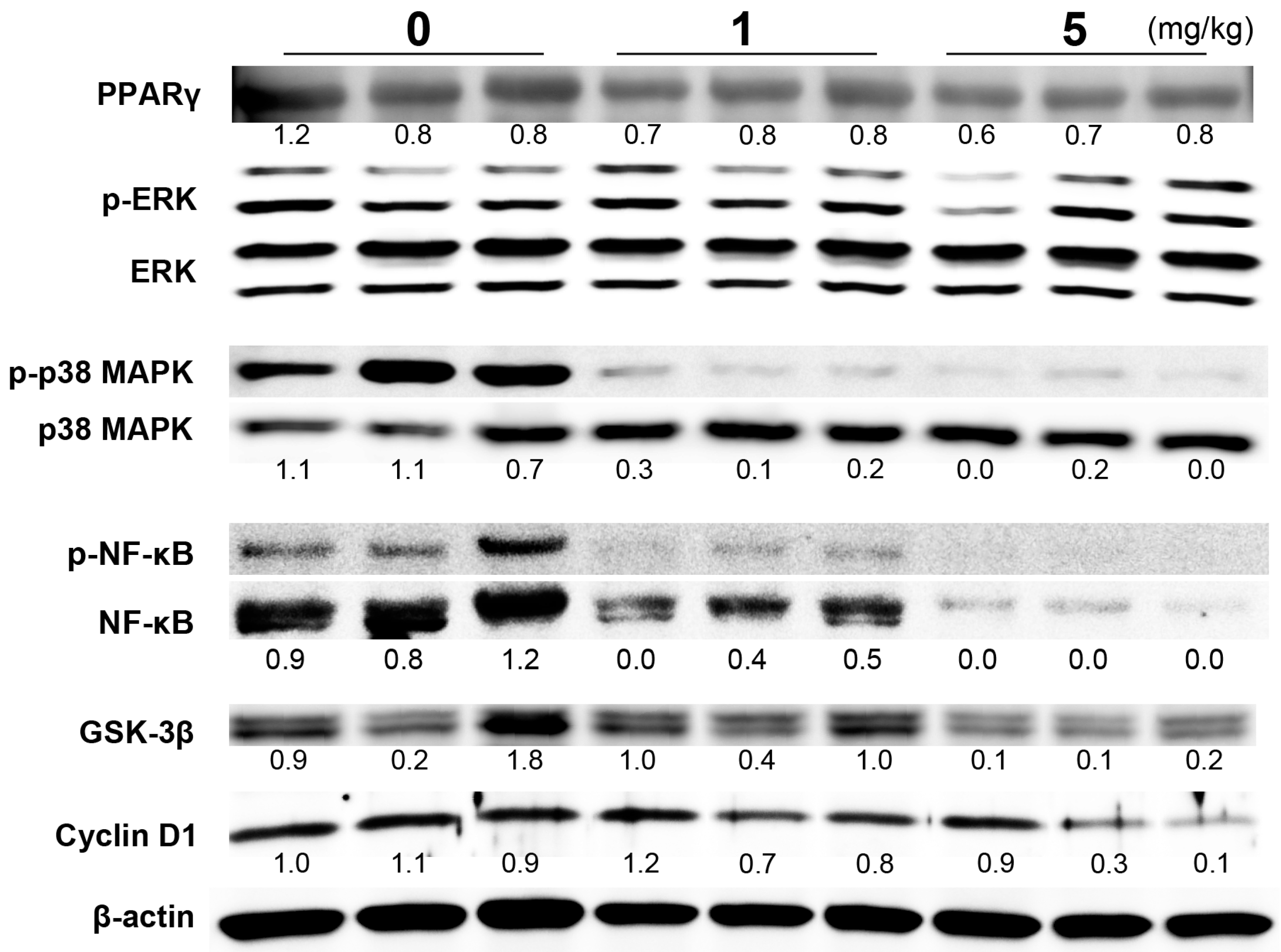{kind=link}
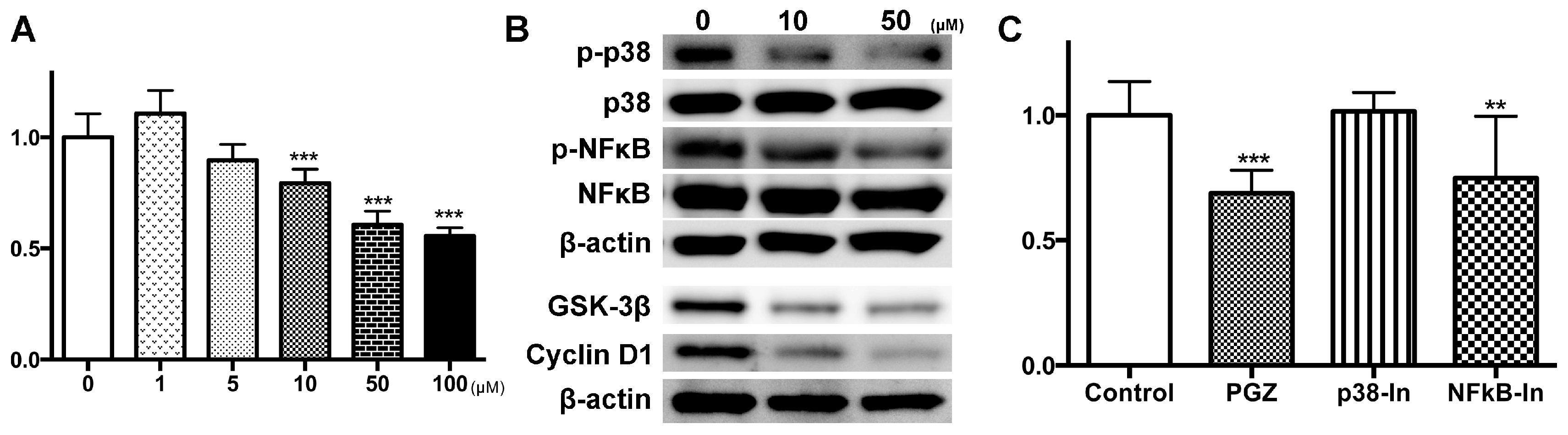{kind=link}
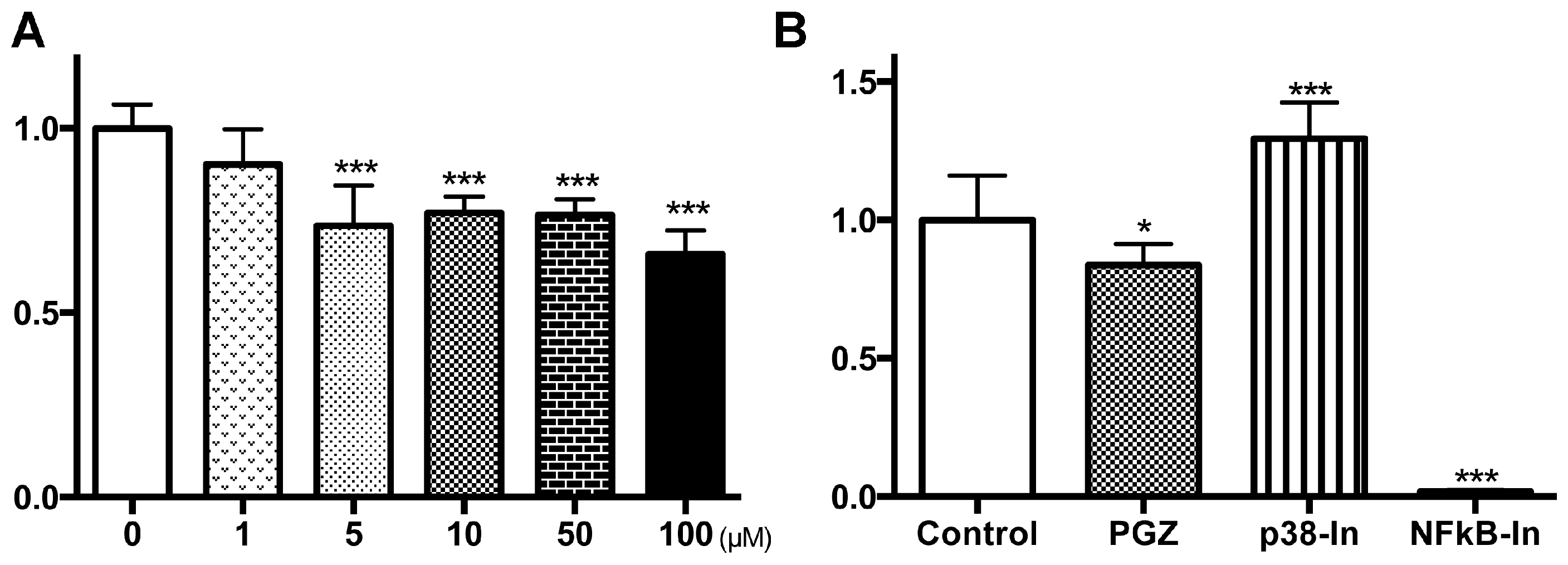{kind=link}
Abstract
Share and Cite
Suzuki, S.; Mori, Y.; Nagano, A.; Naiki-Ito, A.; Kato, H.; Nagayasu, Y.; Kobayashi, M.; Kuno, T.; Takahashi, S. Pioglitazone, a Peroxisome Proliferator-Activated Receptor γ Agonist, Suppresses Rat Prostate Carcinogenesis. Int. J. Mol. Sci. 2016, 17, 2071. https://doi.org/10.3390/ijms17122071
Suzuki S, Mori Y, Nagano A, Naiki-Ito A, Kato H, Nagayasu Y, Kobayashi M, Kuno T, Takahashi S. Pioglitazone, a Peroxisome Proliferator-Activated Receptor γ Agonist, Suppresses Rat Prostate Carcinogenesis. International Journal of Molecular Sciences. 2016; 17(12):2071. https://doi.org/10.3390/ijms17122071
Chicago/Turabian StyleSuzuki, Shugo, Yukiko Mori, Aya Nagano, Aya Naiki-Ito, Hiroyuki Kato, Yuko Nagayasu, Mizuho Kobayashi, Toshiya Kuno, and Satoru Takahashi. 2016. "Pioglitazone, a Peroxisome Proliferator-Activated Receptor γ Agonist, Suppresses Rat Prostate Carcinogenesis" International Journal of Molecular Sciences 17, no. 12: 2071. https://doi.org/10.3390/ijms17122071
APA StyleSuzuki, S., Mori, Y., Nagano, A., Naiki-Ito, A., Kato, H., Nagayasu, Y., Kobayashi, M., Kuno, T., & Takahashi, S. (2016). Pioglitazone, a Peroxisome Proliferator-Activated Receptor γ Agonist, Suppresses Rat Prostate Carcinogenesis. International Journal of Molecular Sciences, 17(12), 2071. https://doi.org/10.3390/ijms17122071