
Lysyl Oxidase and the Tumor Microenvironment



{kind=link}
Abstract
:1. Introduction
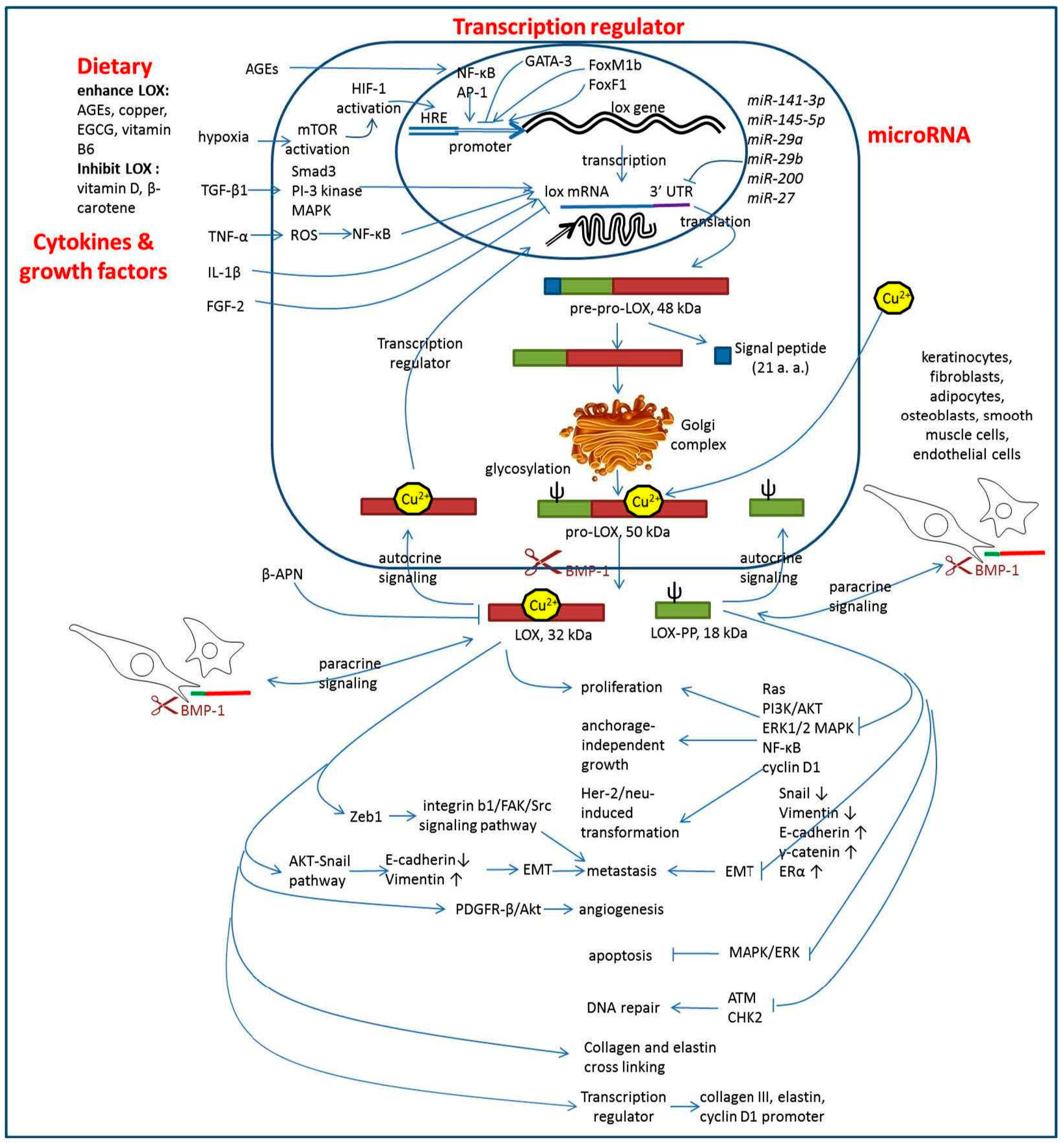
2. LOX Enhances Tumor Progression
3. LOX as a Tumor Suppressor Gene
4. LOX Regulation
5. Downstream Signaling of LOX
6. Dietary Components Regulate LOX Activity
7. LOX as a Target for Anti-Cancer Therapy
8. Conclusions
Acknowledgments
Conflicts of Interest
References
- Harris, E.D. Copper-induced activation of aortic lysyl oxidase in vivo. Proc. Natl. Acad. Sci. USA 1976, 73, 371–374. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.C.; Pinnell, S.R.; Martin, G.R. Cross-linking of collagen and elastin—Properties of lysyl oxidase. Biochemistry 1970, 9, 4486–4492. [Google Scholar] [CrossRef] [PubMed]
- Pischon, N.; Maki, J.M.; Weisshaupt, P.; Heng, N.; Palamakumbura, A.H.; N’Guessan, P.; Ding, A.; Radlanski, R.; Renz, H.; Bronckers, T.A.; et al. Lysyl oxidase (LOX) gene deficiency affects osteoblastic phenotype. Calcif. Tissue Int. 2009, 85, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Giampuzzi, M.; Botti, G.; di Duca, M.; Arata, L.; Ghiggeri, G.; Gusmano, R.; Ravazzolo, R.; Di Donato, A. Lysyl oxidase activates the transcription activity of human collagene III promoter. Possible involvement of Ku antigen. J. Biol. Chem. 2000, 275, 36341–36349. [Google Scholar] [CrossRef] [PubMed]
- Oleggini, R.; Gastaldo, N.; di Donato, A. Regulation of elastin promoter by lysyl oxidase and growth factors: Cross control of lysyl oxidase on TGF-β1 effects. Matrix Biol. 2007, 26, 494–505. [Google Scholar] [CrossRef] [PubMed]
- Giampuzzi, M.; Oleggini, R.; Albanese, C.; Pestell, R.; di Donato, A. β-catenin signaling and regulation of cyclin D1 promoter in NRK-49F cells transformed by down-regulation of the tumor suppressor lysyl oxidase. Biochim. Biophys. Acta 2005, 1745, 370–381. [Google Scholar] [CrossRef] [PubMed]
- Szauter, K.M.; Cao, T.; Boyd, C.D.; Csiszar, K. Lysyl oxidase in development, aging and pathologies of the skin. Pathol. Biol. 2005, 53, 448–456. [Google Scholar] [CrossRef] [PubMed]
- Chanoki, M.; Ishii, M.; Kobayashi, H.; Fushida, H.; Yashiro, N.; Hamada, T.; Ooshima, A. Increased expression of lysyl oxidase in skin with scleroderma. Br. J. Dermatol. 1995, 133, 710–715. [Google Scholar] [CrossRef] [PubMed]
- Trivedy, C.; Warnakulasuriya, K.A.; Hazarey, V.K.; Tavassoli, M.; Sommer, P.; Johnson, N.W. The upregulation of lysyl oxidase in oral submucous fibrosis and squamous cell carcinoma. J. Oral Pathol. Med. 1999, 28, 246–251. [Google Scholar] [CrossRef] [PubMed]
- Mesarwi, O.A.; Shin, M.K.; Drager, L.F.; Bevans-Fonti, S.; Jun, J.C.; Putcha, N.; Torbenson, M.S.; Pedrosa, R.P.; Lorenzi-Filho, G.; Steele, K.E.; et al. Lysyl oxidase as a serum biomarker of liver fibrosis in patients with severe obesity and obstructive sleep apnea. Sleep 2015, 38, 1583–1591. [Google Scholar] [CrossRef] [PubMed]
- Kagan, H.M. Lysyl oxidase: Mechanism, regulation and relationship to liver fibrosis. Pathol. Res. Pract. 1994, 190, 910–919. [Google Scholar] [CrossRef]
- Cheng, T.; Liu, Q.; Zhang, R.; Zhang, Y.; Chen, J.; Yu, R.; Ge, G. Lysyl oxidase promotes bleomycin-induced lung fibrosis through modulating inflammation. J. Mol. Cell Biol. 2014, 6, 506–515. [Google Scholar] [CrossRef] [PubMed]
- Goto, Y.; Uchio-Yamada, K.; Anan, S.; Yamamoto, Y.; Ogura, A.; Manabe, N. Transforming growth factor-β1 mediated up-regulation of lysyl oxidase in the kidneys of hereditary nephrotic mouse with chronic renal fibrosis. Virchows Arch. 2005, 447, 859–868. [Google Scholar] [CrossRef] [PubMed]
- Papadantonakis, N.; Matsuura, S.; Ravid, K. Megakaryocyte pathology and bone marrow fibrosis: The lysyl oxidase connection. Blood 2012, 120, 1774–1781. [Google Scholar] [CrossRef] [PubMed]
- Giampuzzi, M.; Oleggini, R.; di Donato, A. Demonstration of in vitro interaction between tumor suppressor lysyl oxidase and histones H1 and H2: Definition of the regions involved. Biochim. Biophys. Acta 2003, 1647, 245–251. [Google Scholar] [CrossRef]
- Perryman, L.; Erler, J.T. Lysyl oxidase in cancer research. Future Oncol. 2014, 10, 1709–1717. [Google Scholar] [CrossRef] [PubMed]
- Kasashima, H.; Yashiro, M.; Kinoshita, H.; Fukuoka, T.; Morisaki, T.; Masuda, G.; Sakurai, K.; Kubo, N.; Ohira, M.; Hirakawa, A. Lysyl oxidase is associated with the epithelial-mesenchymal transition of gastric cancer cells in hypoxia. Gastric Cancer 2016, 19, 431–442. [Google Scholar] [CrossRef] [PubMed]
- Cox, T.R.; Gartland, A.; Erler, J.T. Lysyl oxidase, a targetable secreted molecule involved in cancer metastasis. Cancer Res. 2016, 76, 188–192. [Google Scholar] [CrossRef] [PubMed]
- Shih, Y.H.; Chang, K.W.; Chen, M.Y.; Yu, C.C.; Lin, D.J.; Hsia, S.M.; Huang, H.L.; Shieh, T.M. Lysyl oxidase and enhancement of cell proliferation and angiogenesis in oral squamous cell carcinoma. Head Neck 2013, 35, 250–256. [Google Scholar] [CrossRef] [PubMed]
- Osawa, T.; Ohga, N.; Akiyama, K.; Hida, Y.; Kitayama, K.; Kawamoto, T.; Yamamoto, K.; Maishi, N.; Kondoh, M.; Onodera, Y.; et al. Lysyl oxidase secreted by tumour endothelial cells promotes angiogenesis and metastasis. Br. J. Cancer 2013, 109, 2237–2247. [Google Scholar] [CrossRef] [PubMed]
- Kaneda, A.; Wakazono, K.; Tsukamoto, T.; Watanabe, N.; Yagi, Y.; Tatematsu, M.; Kaminishi, M.; Sugimura, T.; Ushijima, T. Lysyl oxidase is a tumor suppressor gene inactivated by methylation and loss of heterozygosity in human gastric cancers. Cancer Res. 2004, 64, 6410–6415. [Google Scholar] [CrossRef] [PubMed]
- Alsulaiman, M.; Bais, M.V.; Trackman, P.C. Lysyl oxidase propeptide stimulates osteoblast and osteoclast differentiation and enhances PC3 and DU145 prostate cancer cell effects on bone in vivo. J. Cell Commun. Signal. 2016, 10, 17–31. [Google Scholar] [CrossRef] [PubMed]
- Ozdener, G.B.; Bais, M.V.; Trackman, P.C. Determination of cell uptake pathways for tumor inhibitor lysyl oxidase propeptide. Mol. Oncol. 2016, 10, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Bais, M.V.; Ozdener, G.B.; Sonenshein, G.E.; Trackman, P.C. Effects of tumor-suppressor lysyl oxidase propeptide on prostate cancer xenograft growth and its direct interactions with DNA repair pathways. Oncogene 2015, 34, 1928–1937. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Wang, X.; Wang, H.; Yan, W.; Zhang, Q.; Chang, X. Expression of the lysyl oxidase propeptide in hepatocellular carcinoma and its clinical relevance. Oncol. Rep. 2014, 31, 1669–1676. [Google Scholar] [CrossRef] [PubMed]
- Woznick, A.R.; Braddock, A.L.; Dulai, M.; Seymour, M.L.; Callahan, R.E.; Welsh, R.J.; Chmielewski, G.W.; Zelenock, G.B.; Shanley, C.J. Lysyl oxidase expression in bronchogenic carcinoma. Am. J. Surg. 2005, 189, 297–301. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.J.; Li, Y.G.; Huang, L.; Han, M.; Ma, B.J.; Sun, B.J.; Lin, J.J.; Song, T.G. Expression of LOX and MMP-2 in gastric cancer tissue and the effects of LOX and MMP-2 on tumor invasion and metastasis. Zhonghua Zhong Liu Za Zhi 2011, 33, 37–41. [Google Scholar] [PubMed]
- Kirschmann, D.A.; Seftor, E.A.; Fong, S.F.; Nieva, D.R.; Sullivan, C.M.; Edwards, E.M.; Sommer, P.; Csiszar, K.; Hendrix, M.J. A molecular role for lysyl oxidase in breast cancer invasion. Cancer Res. 2002, 62, 4478–4483. [Google Scholar] [PubMed]
- Boufraqech, M.; Nilubol, N.; Zhang, L.; Gara, S.K.; Sadowski, S.M.; Mehta, A.; He, M.; Davis, S.; Dreiling, J.; Copland, J.A.; et al. miR30a inhibits LOX expression and anaplastic thyroid cancer progression. Cancer Res. 2015, 75, 367–377. [Google Scholar] [CrossRef] [PubMed]
- Baker, A.M.; Cox, T.R.; Bird, D.; Lang, G.; Murray, G.I.; Sun, X.F.; Southall, S.M.; Wilson, J.R.; Erler, J.T. The role of lysyl oxidase in SRC-dependent proliferation and metastasis of colorectal cancer. J. Natl. Cancer Inst. 2011, 103, 407–424. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, R.; Uno, M.; Marie, S.K.; Oba-Shinjo, S.M. LOX expression and functional analysis in astrocytomas and impact of IDH1 mutation. PLoS ONE 2015, 10, e0119781. [Google Scholar] [CrossRef] [PubMed]
- Jung, H.Y.; Fattet, L.; Yang, J. Molecular pathways: Linking tumor microenvironment to epithelial-mesenchymal transition in metastasis. Clin. Cancer Res. 2015, 21, 962–968. [Google Scholar] [CrossRef] [PubMed]
- Albinger-Hegyi, A.; Stoeckli, S.J.; Schmid, S.; Storz, M.; Iotzova, G.; Probst-Hensch, N.M.; Rehrauer, H.; Tinguely, M.; Moch, H.; Hegyi, I. Lysyl oxidase expression is an independent marker of prognosis and a predictor of lymph node metastasis in oral and oropharyngeal squamous cell carcinoma (OSCC). Int. J. Cancer 2010, 126, 2653–2662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bondareva, A.; Downey, C.M.; Ayres, F.; Liu, W.; Boyd, S.K.; Hallgrimsson, B.; Jirik, F.R. The lysyl oxidase inhibitor, β-aminopropionitrile, diminishes the metastatic colonization potential of circulating breast cancer cells. PLoS ONE 2009, 4, e5620. [Google Scholar] [CrossRef] [PubMed]
- Wilgus, M.L.; Borczuk, A.C.; Stoopler, M.; Ginsburg, M.; Gorenstein, L.; Sonett, J.R.; Powell, C.A. Lysyl oxidase: A lung adenocarcinoma biomarker of invasion and survival. Cancer 2011, 117, 2186–2191. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Zhou, J.; Zhao, Y.; Toselli, P.; Li, W. Hypoxia-response element (HRE)-directed transcriptional regulation of the rat lysyl oxidase gene in response to cobalt and cadmium. Toxicol. Sci. 2013, 132, 379–389. [Google Scholar] [CrossRef] [PubMed]
- Erler, J.T.; Giaccia, A.J. Lysyl oxidase mediates hypoxic control of metastasis. Cancer Res. 2006, 66, 10238–10241. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Li, S.; Li, W.; Chen, J.; Xiao, X.; Wang, Y.; Yan, G.; Chen, L. Inactivation of lysyl oxidase by β-aminopropionitrile inhibits hypoxia-induced invasion and migration of cervical cancer cells. Oncol. Rep. 2013, 29, 541–548. [Google Scholar] [PubMed]
- Semenza, G.L. Cancer-stromal cell interactions mediated by hypoxia-inducible factors promote angiogenesis, lymphangiogenesis, and metastasis. Oncogene 2013, 32, 4057–4063. [Google Scholar] [CrossRef] [PubMed]
- Baker, A.M.; Bird, D.; Welti, J.C.; Gourlaouen, M.; Lang, G.; Murray, G.I.; Reynolds, A.R.; Cox, T.R.; Erler, J.T. Lysyl oxidase plays a critical role in endothelial cell stimulation to drive tumor angiogenesis. Cancer Res. 2013, 73, 583–594. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Tang, H.J.; Wang, Y.Y.; Xiong, M.H.; Zhou, F.; Shao, K.; Li, T.P. Expression of lysyl oxidase gene in upper digestive tract carcinomas and its clinical significance. Chin. J. Cancer 2002, 21, 671–674. [Google Scholar]
- Xu, X.; Wang, B.; Xu, Y. Expression of lysyl oxidase in human osteosarcoma and its clinical significance: A tumor suppressive role of LOX in human osteosarcoma cells. Int. J. Oncol. 2013, 43, 1578–1586. [Google Scholar] [CrossRef] [PubMed]
- Kenyon, K.; Contente, S.; Trackman, P.C.; Tang, J.; Kagan, H.M.; Friedman, R.M. Lysyl oxidase and rrg messenger RNA. Science 1991, 253, 802. [Google Scholar] [CrossRef] [PubMed]
- Su, Z.; Yemul, S.; Estabrook, A.; Friedman, R.; Zimmer, S.; Fisher, P. Transcriptional switching model for the regulation of tumorigenesis and metastasis by the ha-ras oncogene—Transcriptional changes in the Ha-ras tumor-suppressor gene lysyl oxidase. Int. J. Oncol. 1995, 7, 1279–1284. [Google Scholar] [CrossRef] [PubMed]
- Tan, R.S.; Taniguchi, T.; Harada, H. Identification of the lysyl oxidase gene as target of the antioncogenic transcription factor, IRF-1, and its possible role in tumor suppression. Cancer Res. 1996, 56, 2417–2421. [Google Scholar] [PubMed]
- Contente, S.; Kenyon, K.; Sriraman, P.; Subramanyan, S.; Friedman, R.M. Epigenetic inhibition of lysyl oxidase transcription after transformation by ras oncogene. Mol. Cell. Biochem. 1999, 194, 79–91. [Google Scholar] [CrossRef] [PubMed]
- Palamakumbura, A.H.; Jeay, S.; Guo, Y.; Pischon, N.; Sommer, P.; Sonenshein, G.E.; Trackman, P.C. The propeptide domain of lysyl oxidase induces phenotypic reversion of ras-transformed cells. J. Biol. Chem. 2004, 279, 40593–40600. [Google Scholar] [CrossRef] [PubMed]
- Min, C.; Kirsch, K.H.; Zhao, Y.; Jeay, S.; Palamakumbura, A.H.; Trackman, P.C.; Sonenshine, G.E. The tumor suppressor activity of the lysyl oxidase propeptide reverses the invasive phenotype of Her-2/neu-driven breast cancer. Cancer Res. 2007, 67, 1105–1112. [Google Scholar] [CrossRef] [PubMed]
- Vadasz, Z.; Kessler, O.; Akiri, G.; Gengrinovitch, S.; Kagan, H.M.; Baruch, Y.; Izack, O.B.; Nuefeld, G. Abnormal deposition of collagen around hepatocytes in Wilson’s disease is associated with hepatocyte specific expression of lysyl oxidase and lysyl oxidase like protein-2. J. Hepatol. 2005, 43, 499–507. [Google Scholar] [CrossRef] [PubMed]
- Barker, H.E.; Cox, T.R.; Erler, J.T. The rationale for targeting the LOX family in cancer. Nat. Rev. Cancer 2012, 12, 540–552. [Google Scholar] [CrossRef] [PubMed]
- Atsawasuwan, P.; Mochida, Y.; Katafuchi, M.; Kaku, M.; Fong, K.S.; Csiszar, K.; Yamauchi, M. Lysyl oxidase binds transforming growth factor-β and regulates its signaling via amine oxidase activity. J. Biol. Chem. 2008, 283, 34229–34240. [Google Scholar] [CrossRef] [PubMed]
- Taylor, M.A.; Amin, J.D.; Kirschmann, D.A.; Schiemann, W.P. Lysyl oxidase contributes to mechanotransduction-mediated regulation of transforming growth factor-β signaling in breast cancer cells. Neoplasia 2011, 13, 406–418. [Google Scholar] [CrossRef] [PubMed]
- Voloshenyuk, T.G.; Landesman, E.S.; Khoutorova, E.; Hart, A.D.; Gardner, J.D. Induction of cardiac fibroblast lysyl oxidase by TGF-β1 requires PI3K/Akt, Smad3, and MAPK signaling. Cytokine 2011, 55, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Voloshenyuk, T.G.; Hart, A.D.; Khoutorova, E.; Gardner, J.D. TNF-α increases cardiac fibroblast lysyl oxidase expression through TGF-β and PI3Kinase signaling pathways. Biochem. Biophys. Res. Commun. 2011, 413, 370–375. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, C.; Alcudia, J.F.; Martinez-Gonzalez, J.; Raposo, B.; Navarro, M.A.; Badimon, L. Lysyl oxidase (LOX) down-regulation by TNFα: A new mechanism underlying TNFα-induced endothelial dysfunction. Atherosclerosis 2008, 196, 558–564. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Wang, C.; Yin, L.; Xu, C.; Zhang, Y.; Sung, K.L. Interleukin-1 β influences on lysyl oxidases and matrix metalloproteinases profile of injured anterior cruciate ligament and medial collateral ligament fibroblasts. Int. Orthop. 2013, 37, 495–505. [Google Scholar] [CrossRef] [PubMed]
- Cenizo, V.; Andre, V.; Reymermier, C.; Sommer, P.; Damour, O.; Perrier, E. LOXL as a target to increase the elastin content in adult skin: A dill extract induces the LOXL gene expression. Exp. Dermatol. 2006, 15, 574–581. [Google Scholar] [CrossRef] [PubMed]
- Roy, R.; Polgar, P.; Wang, Y.; Goldstein, R.H.; Taylor, L.; Kagan, H.M. Regulation of lysyl oxidase and cyclooxygenase expression in human lung fibroblasts: Interactions among TGF-β, IL-1 β, and prostaglandin E. J. Cell. Biochem. 1996, 62, 411–417. [Google Scholar] [CrossRef]
- Palamakumbura, A.H.; Sommer, P.; Trackman, P.C. Autocrine growth factor regulation of lysyl oxidase expression in transformed fibroblasts. J. Biol. Chem. 2003, 278, 30781–30787. [Google Scholar] [CrossRef] [PubMed]
- Chu, I.M.; Michalowski, A.M.; Hoenerhoff, M.; Szauter, K.M.; Luger, D.; Sato, M.; Flanders, K.; Oshima, A.; Csiszar, K.; Green, J.E. GATA3 inhibits lysyl oxidase-mediated metastases of human basal triple-negative breast cancer cells. Oncogene 2012, 31, 2017–2027. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Gusarova, G.; Wang, Z.; Carr, J.R.; Li, J.; Kim, K.H.; Qiu, J.; Park, Y.D.; Williamson, P.R.; Hay, N.; et al. Deregulation of FoxM1b leads to tumour metastasis. EMBO Mol. Med. 2011, 3, 21–34. [Google Scholar] [CrossRef] [PubMed]
- Erler, J.T.; Bennewith, K.L.; Nicolau, M.; Dornhofer, N.; Kong, C.; Le, Q.T.; Chi, J.T.; Jeffrey, S.S.; Giaccia, A.J. Lysyl oxidase is essential for hypoxia-induced metastasis. Nature 2006, 440, 1222–1226. [Google Scholar] [CrossRef] [PubMed]
- Mayorca-Guiliani, A.; Erler, J.T. The potential for targeting extracellular LOX proteins in human malignancy. OncoTargets Ther. 2013, 6, 1729–1735. [Google Scholar]
- Sahlgren, C.; Gustafsson, M.V.; Jin, S.; Poellinger, L.; Lendahl, U. Notch signaling mediates hypoxia-induced tumor cell migration and invasion. Proc. Natl. Acad. Sci. USA 2008, 105, 6392–6397. [Google Scholar] [CrossRef] [PubMed]
- Sumual, S.; Saad, S.; Tang, O.; Yong, R.; McGinn, S.; Chen, X.M.; Pollock, C.A. Differential regulation of Snail by hypoxia and hyperglycemia in human proximal tubule cells. Int. J. Biochem. Cell Biol. 2010, 42, 1689–1697. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Xiao, Q.; Ma, H.; Li, L.; Liu, J.; Feng, Y.; Fang, Z.; Wu, J.; Han, X.; Zhang, J.; et al. LKB1 inhibits lung cancer progression through lysyl oxidase and extracellular matrix remodeling. Proc. Natl. Acad. Sci. USA 2010, 107, 18892–18897. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Li, F.; Fang, Z.; Gao, Y.; Li, F.; Fang, R.; Yao, S.; Sun, Y.; Li, L.; Zhang, W.; et al. Transdifferentiation of lung adenocarcinoma in mice with Lkb1 deficiency to squamous cell carcinoma. Nat. Commun. 2014, 5, 3261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, B.; Gogol, M.; Gaudenz, K.; Gerton, J.L. Improved transcription and translation with l-leucine stimulation of mTORC1 in Roberts syndrome. BMC Genom. 2016, 17, 25. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Lee, K.K.; Zhang, L.; Gerton, J.L. Stimulation of mTORC1 with l-leucine rescues defects associated with Roberts syndrome. PLoS Genet. 2013, 9, e1003857. [Google Scholar] [CrossRef] [PubMed]
- Peng, D.H.; Ungewiss, C.; Tong, P.; Byers, L.A.; Wang, J.; Canales, J.R.; Villalobos, P.A.; Uraoka, N.; Mino, B.; Behrens, C.; et al. ZEB1 induces LOXL2-mediated collagen stabilization and deposition in the extracellular matrix to drive lung cancer invasion and metastasis. Oncogene 2016. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, G.; Kannius-Janson, M. Forkhead Box F1 promotes breast cancer cell migration by upregulating lysyl oxidase and suppressing Smad2/3 signaling. BMC Cancer 2016, 16, 142. [Google Scholar] [CrossRef] [PubMed]
- Liep, J.; Kilic, E.; Meyer, H.A.; Busch, J.; Jung, K.; Rabien, A. Cooperative Effect of miR-141–3p and miR-145–5p in the regulation of targets in clear cell renal cell carcinoma. PLoS ONE 2016, 11, e0157801. [Google Scholar] [CrossRef] [PubMed]
- Hebrant, A.; Floor, S.; Saiselet, M.; Antoniou, A.; Desbuleux, A.; Snyers, B.; La, C.; de Saint Aubainet, N.; Leteuttre, E.; Andry, G.; et al. miRNA expression in anaplastic thyroid carcinomas. PLoS ONE 2014, 9, e103871. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Ghazwani, M.; Li, J.; Sun, M.; Stolz, D.B.; He, F.; Fan, J.; Xie, W.; Li, S. MiR-29b inhibits collagen maturation in hepatic stellate cells through down-regulating the expression of HSP47 and lysyl oxidase. Biochem. Biophys. Res. Commun. 2014, 446, 940–944. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Gomes, S.; Chen, P.; Frankenberger, C.A.; Sankarasharma, D.; Chung, C.H.; Chada, K.K.; Rosner, S.R. RKIP and HMGA2 regulate breast tumor survival and metastasis through lysyl oxidase and syndecan-2. Oncogene 2014, 33, 3528–3537. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Hou, Y.; Yang, G.; Wang, X.; Tang, S.; Du, Y.E.; Yang, L.; Yu, T.; Zhang, H.; Zhou, M.; et al. Stromal miR-200s contribute to breast cancer cell invasion through CAF activation and ECM remodeling. Cell Death Differ. 2016, 23, 132–145. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.Z.; Xu, X.; Ning, L.F.; Jiang, W.Y.; Xing, C.; Tang, Q.Q.; Huang, H.Y. miR-27 impairs the adipogenic lineage commitment via targeting lysyl oxidase. Obesity 2015, 23, 2445–2453. [Google Scholar] [CrossRef] [PubMed]
- Agra, N.; Cidre, F.; Garcia-Garcia, L.; de la Parra, J.; Alonso, J. Lysyl oxidase is downregulated by the EWS/FLI1 oncoprotein and its propeptide domain displays tumor supressor activities in Ewing sarcoma cells. PLoS ONE 2013, 8, e66281. [Google Scholar] [CrossRef] [PubMed]
- Peng, L.; Ran, Y.L.; Hu, H.; Yu, L.; Liu, Q.; Zhou, Z.; Sun, Y.M.; Sun, L.C.; Pan, J.; Sun, L.X.; et al. Secreted LOXL2 is a novel therapeutic target that promotes gastric cancer metastasis via the Src/FAK pathway. Carcinogenesis 2009, 30, 1660–1669. [Google Scholar] [CrossRef] [PubMed]
- Baker, A.M.; Bird, D.; Lang, G.; Cox, T.R.; Erler, J.T. Lysyl oxidase enzymatic function increases stiffness to drive colorectal cancer progression through FAK. Oncogene 2013, 32, 1863–1868. [Google Scholar] [CrossRef] [PubMed]
- Li, R.K.; Zhao, W.Y.; Fang, F.; Zhuang, C.; Zhang, X.X.; Yang, X.M.; Jiang, S.H.; Kong, F.Z.; Tu, L.; Zhang, W.M.; et al. Lysyl oxidase-like 4 (LOXL4) promotes proliferation and metastasis of gastric cancer via FAK/Src pathway. J. Cancer Res. Clin. Oncol. 2015, 141, 269–281. [Google Scholar] [CrossRef] [PubMed]
- Pez, F.; Dayan, F.; Durivault, J.; Kaniewski, B.; Aimond, G.; le Provost, G.S.; Duex, B.; Clezardin, P.; Sommer, P.; Pouyssegur, J.; et al. The HIF-1-inducible lysyl oxidase activates HIF-1 via the Akt pathway in a positive regulation loop and synergizes with HIF-1 in promoting tumor cell growth. Cancer Res. 2011, 71, 1647–1657. [Google Scholar] [CrossRef] [PubMed]
- Setozaki, S.; Minakata, K.; Masumoto, H.; Hirao, S.; Yamazaki, K.; Kuwahara, K.; Ikeda, T.; Sakata, R. Prevention of abdominal aortic aneurysm progression by oral administration of green tea polyphenol in a rat model. J. Vasc. Surg. 2016. [Google Scholar] [CrossRef] [PubMed]
- Rucker, R.B.; Rucker, B.R.; Mitchell, A.E.; Cui, C.T.; Clegg, M.; Kosonen, T.; Uriu-Adams, J.Y.; Tchaparian, E.H.; Fishman, M.; Keen, C.L. Activation of chick tendon lysyl oxidase in response to dietary copper. J. Nutr. 1999, 129, 2143–2146. [Google Scholar] [PubMed]
- Pelham, C.J.; Drews, E.M.; Agrawal, D.K. Vitamin D controls resistance artery function through regulation of perivascular adipose tissue hypoxia and inflammation. J. Mol. Cell. Cardiol. 2016, 98, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Myers, B.A.; Dubick, M.A.; Gerriets, J.E.; Reiser, K.M.; Last, J.A.; Rucker, R.B. Lung collagen and elastin after ozone exposure in vitamin B-6-deficient rats. Toxicol. Lett. 1986, 30, 55–61. [Google Scholar] [CrossRef]
- Guruvayoorappan, C.; Kuttan, G. β-carotene inhibits tumor-specific angiogenesis by altering the cytokine profile and inhibits the nuclear translocation of transcription factors in B16F-10 melanoma cells. Integr. Cancer Ther. 2007, 6, 258–270. [Google Scholar] [CrossRef] [PubMed]
- Deluyker, D.; Ferferieva, V.; Noben, J.P.; Swennen, Q.; Bronckaers, A.; Lambrichts, I.; Rigo, J.M.; Bito, V. Cross-linking versus RAGE: How do high molecular weight advanced glycation products induce cardiac dysfunction? Int. J. Cardiol. 2016, 210, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Aoki, C.; Uto, K.; Honda, K.; Kato, Y.; Oda, H. Advanced glycation end products suppress lysyl oxidase and induce bone collagen degradation in a rat model of renal osteodystrophy. Lab. Investig. 2013, 93, 1170–1183. [Google Scholar] [CrossRef] [PubMed]
- Akagawa, M.; Shigemitsu, T.; Suyama, K. Oxidative deamination of benzylamine and lysine residue in bovine serum albumin by green tea, black tea, and coffee. J. Agric. Food Chem. 2005, 53, 8019–8124. [Google Scholar] [CrossRef] [PubMed]
- Adamopoulos, C.; Piperi, C.; Gargalionis, A.N.; Dalagiorgou, G.; Spilioti, E.; Korkolopoulou, P.; Diamanti-Kandarakis, E.; Papavassiliou, A.G. Advanced glycation end products upregulate lysyl oxidase and endothelin-1 in human aortic endothelial cells via parallel activation of ERK1/2-NF-kappaB and JNK-AP-1 signaling pathways. Cell. Mol. Life Sci. 2016, 73, 1685–1698. [Google Scholar] [CrossRef] [PubMed]
- Song, F.; Hurtado del Pozo, C.; Rosario, R.; Zou, Y.S.; Ananthakrishnan, R.; Xu, X.; Petel, P.R.; Benoit, V.M.; Yan, S.F.; Li, H.; et al. RAGE regulates the metabolic and inflammatory response to high-fat feeding in mice. Diabetes 2014, 63, 1948–1965. [Google Scholar] [CrossRef] [PubMed]
- Kong, S.Y.; Takeuchi, M.; Hyogo, H.; McKeown-Eyssen, G.; Yamagishi, S.; Chayama, K.; O’Brien, P.J.; Ferrari, P.; Overvad, K.; Olsen, A.; et al. The association between glyceraldehyde-derived advanced glycation end-products and colorectal cancer risk. Cancer Epidemiol. Biomark. Prev. 2015, 24, 1855–1863. [Google Scholar] [CrossRef] [PubMed]
- Jiao, L.; Weinstein, S.J.; Albanes, D.; Taylor, P.R.; Graubard, B.I.; Virtamo, J.; Stolzenberg-Solomon, R.Z. Evidence that serum levels of the soluble receptor for advanced glycation end products are inversely associated with pancreatic cancer risk: A prospective study. Cancer Res. 2011, 71, 3582–3589. [Google Scholar] [CrossRef] [PubMed]
- Ajith, T.A.; Vinodkumar, P. Advanced glycation end products: Association with the pathogenesis of diseases and the current therapeutic advances. Curr. Clin. Pharmacol. 2016, 11, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Papachroni, K.K.; Piperi, C.; Levidou, G.; Korkolopoulou, P.; Pawelczyk, L.; Diamanti-Kandarakis, E.; Papavassiliou, A.G. Lysyl oxidase interacts with AGE signalling to modulate collagen synthesis in polycystic ovarian tissue. J. Cell. Mol. Med. 2010, 14, 2460–2469. [Google Scholar] [CrossRef] [PubMed]
- Di Ferrante, N.; Leachman, R.D.; Angelini, P.; Donnelly, P.V.; Francis, G.; Almazan, A.; Segni, G.; Franzblau, C.; Jorden, R.E. Ehlers-Danlos type V (X-linked form): A lysyl oxidase deficiency. Birth Defects Orig. Artic. Ser. 1975, 11, 31–37. [Google Scholar] [PubMed]
- Kuivaniemi, H.; Peltonen, L.; Kivirikko, K.I.; Type, I.X. Ehlers-Danlos syndrome and Menkes syndrome: The decrease in lysyl oxidase activity is associated with a corresponding deficiency in the enzyme protein. Am. J. Hum. Genet. 1985, 37, 798–808. [Google Scholar] [PubMed]
- Preethi, K.C.; Siveen, K.S.; Kuttan, R.; Kuttan, G. Inhibition of metastasis of B16F-10 melanoma cells in C57BL/6 mice by an extract of Calendula officinalis L. flowers. Asian Pac. J. Cancer Prev. 2010, 11, 1773–1779. [Google Scholar] [PubMed]
- Akiri, G.; Sabo, E.; Dafni, H.; Vadasz, Z.; Kartvelishvily, Y.; Gan, N.; Kessler, O.; Cohen, T.; Resnick, M.; Neeman, M.; et al. Lysyl oxidase-related protein-1 promotes tumor fibrosis and tumor progression in vivo. Cancer Res. 2003, 63, 1657–1666. [Google Scholar] [PubMed]
- Zibadi, S.; Vazquez, R.; Moore, D.; Larson, D.F.; Watson, R.R. Myocardial lysyl oxidase regulation of cardiac remodeling in a murine model of diet-induced metabolic syndrome. Am. J. Physiol. Heart Circ. Physiol. 2009, 297, H976–H982. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Martinez, E.; Rodriguez, C.; Galan, M.; Miana, M.; Jurado-Lopez, R.; Bartolome, M.V.; Luaces, M.; Islas, F.; Martinez-Gonzales, J.; Lopez-Andres, N.; et al. The lysyl oxidase inhibitor (β-aminopropionitrile) reduces leptin profibrotic effects and ameliorates cardiovascular remodeling in diet-induced obesity in rats. J. Mol. Cell. Cardiol. 2016, 92, 96–104. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, C.; Rodriguez-Sinovas, A.; Martinez-Gonzalez, J. Lysyl oxidase as a potential therapeutic target. Drug News Perspect. 2008, 21, 218–224. [Google Scholar] [CrossRef] [PubMed]
- Le Calve, B.; Griveau, A.; Vindrieux, D.; Marechal, R.; Wiel, C.; Svrcek, M.; Gout, J.; Azzi, L.; Payen, L.; Cros, J.; et al. Lysyl oxidase family activity promotes resistance of pancreatic ductal adenocarcinoma to chemotherapy by limiting the intratumoral anticancer drug distribution. Oncotarget 2016, 7, 32100–32112. [Google Scholar] [PubMed]
- Schutze, F.; Rohrig, F.; Vorlova, S.; Gatzner, S.; Kuhn, A.; Ergun, S.; Henke, E. Inhibition of lysyl oxidases improves drug diffusion and increases efficacy of cytotoxic treatment in 3D tumor models. Sci. Rep. 2015, 5, 17576. [Google Scholar] [CrossRef] [PubMed]
- Kanapathipillai, M.; Mammoto, A.; Mammoto, T.; Kang, J.H.; Jiang, E.; Ghosh, K.; Korin, N.; Gibbs, A.; Mannix, R.; Ingber, G.E. Inhibition of mammary tumor growth using lysyl oxidase-targeting nanoparticles to modify extracellular matrix. Nano Lett. 2012, 12, 3213–3217. [Google Scholar] [CrossRef] [PubMed]
- Gong, C.; Gu, R.; Jin, H.; Sun, Y.; Li, Z.; Chen, J.; Wu, G. Lysyl oxidase mediates hypoxia-induced radioresistance in non-small cell lung cancer A549 cells. Exp. Biol. Med. 2016, 241, 387–395. [Google Scholar] [CrossRef] [PubMed]

© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, T.-H.; Hsia, S.-M.; Shieh, T.-M. Lysyl Oxidase and the Tumor Microenvironment. Int. J. Mol. Sci. 2017, 18, 62. https://doi.org/10.3390/ijms18010062
Wang T-H, Hsia S-M, Shieh T-M. Lysyl Oxidase and the Tumor Microenvironment. International Journal of Molecular Sciences. 2017; 18(1):62. https://doi.org/10.3390/ijms18010062
Chicago/Turabian StyleWang, Tong-Hong, Shih-Min Hsia, and Tzong-Ming Shieh. 2017. "Lysyl Oxidase and the Tumor Microenvironment" International Journal of Molecular Sciences 18, no. 1: 62. https://doi.org/10.3390/ijms18010062