Comparative Morphology, Transcription, and Proteomics Study Revealing the Key Molecular Mechanism of Camphor on the Potato Tuber Sprouting Effect

Abstract
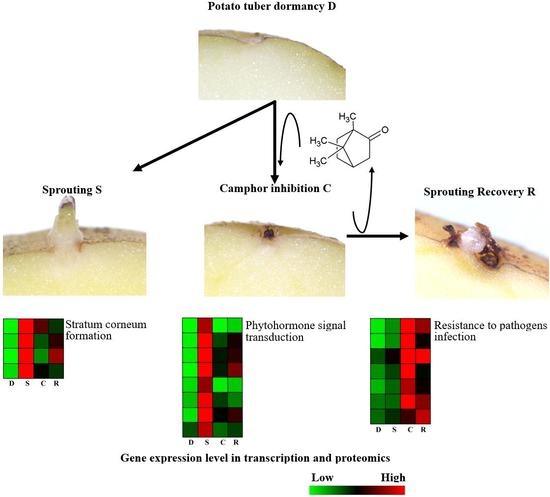
:
1. Introduction
2. Results
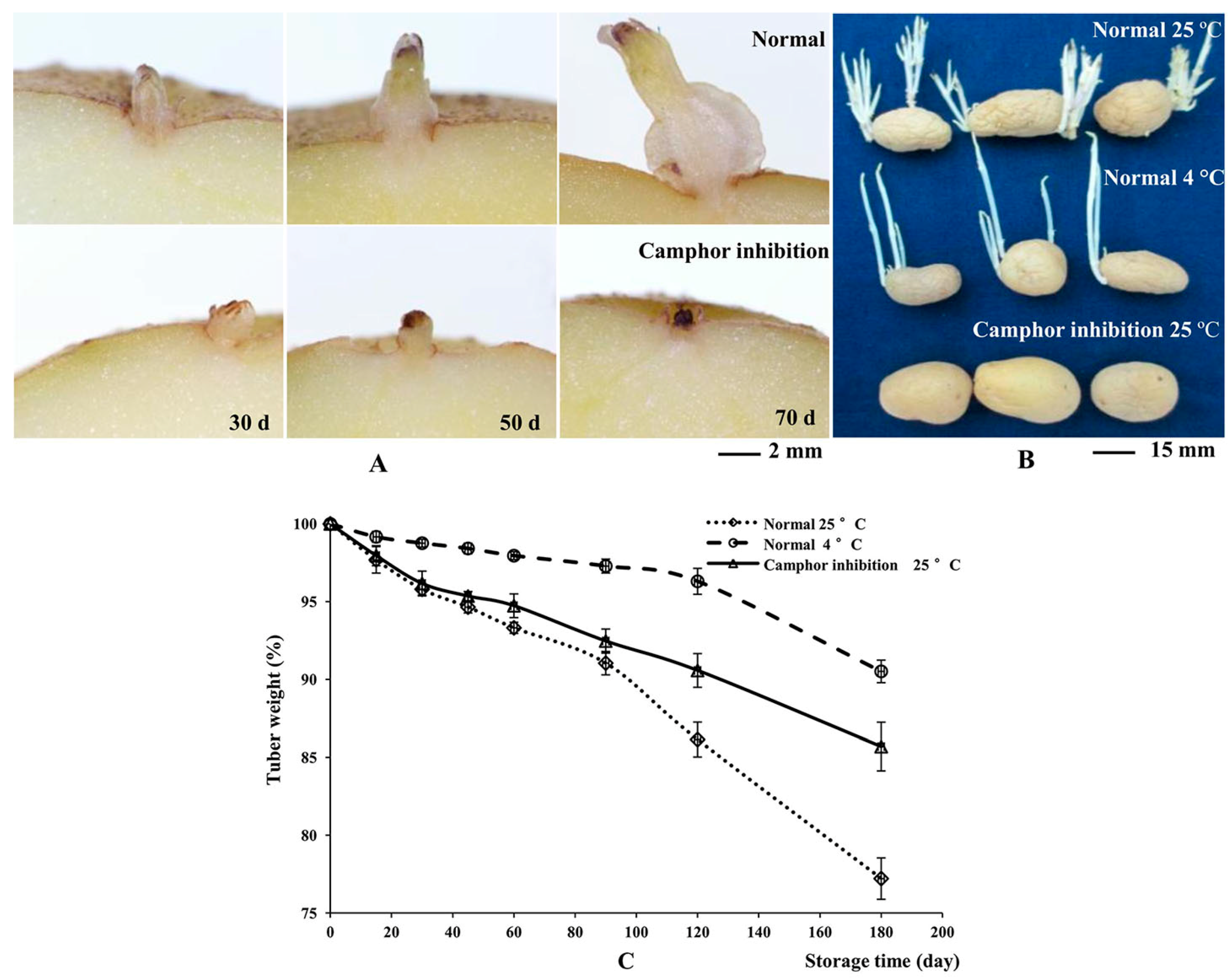
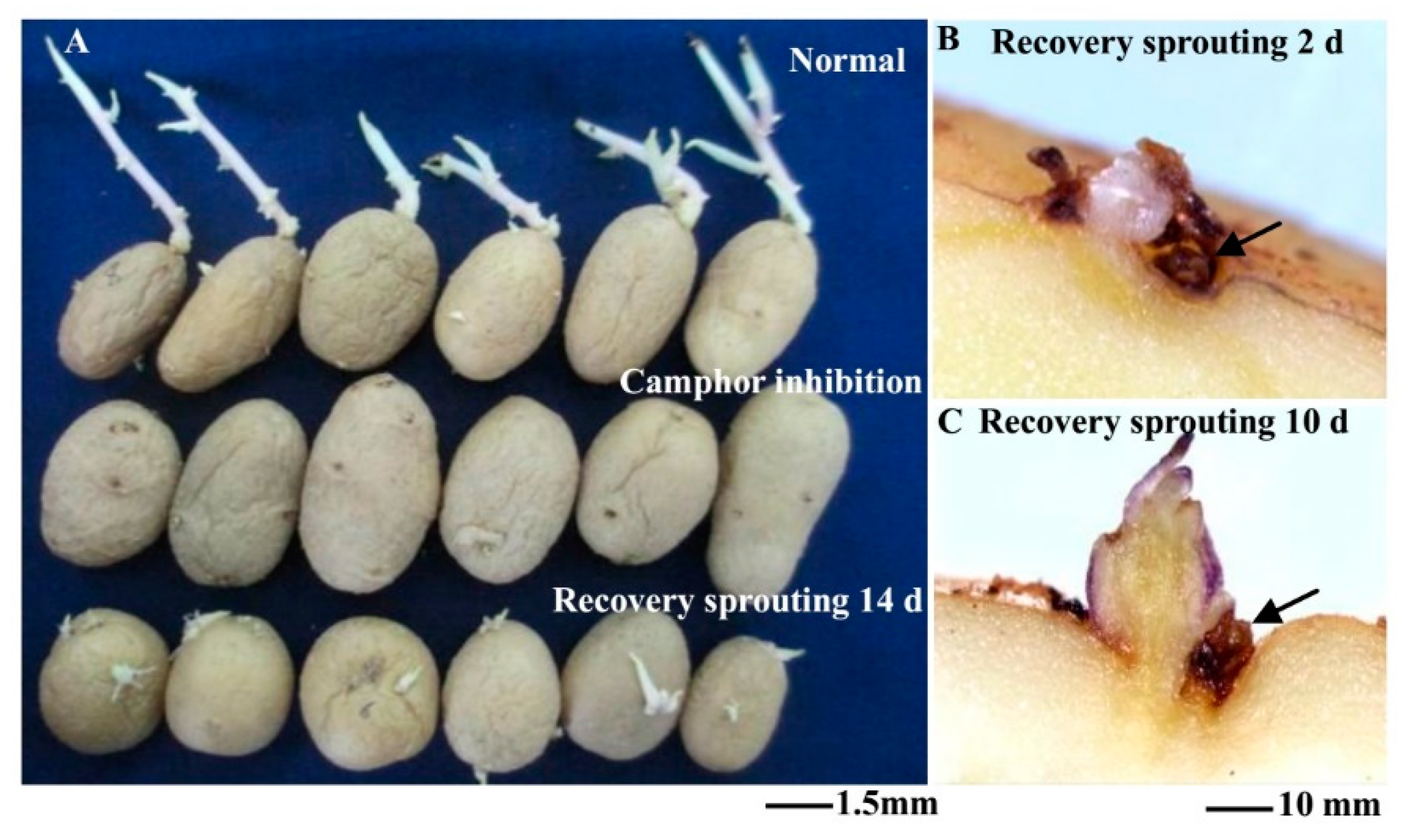
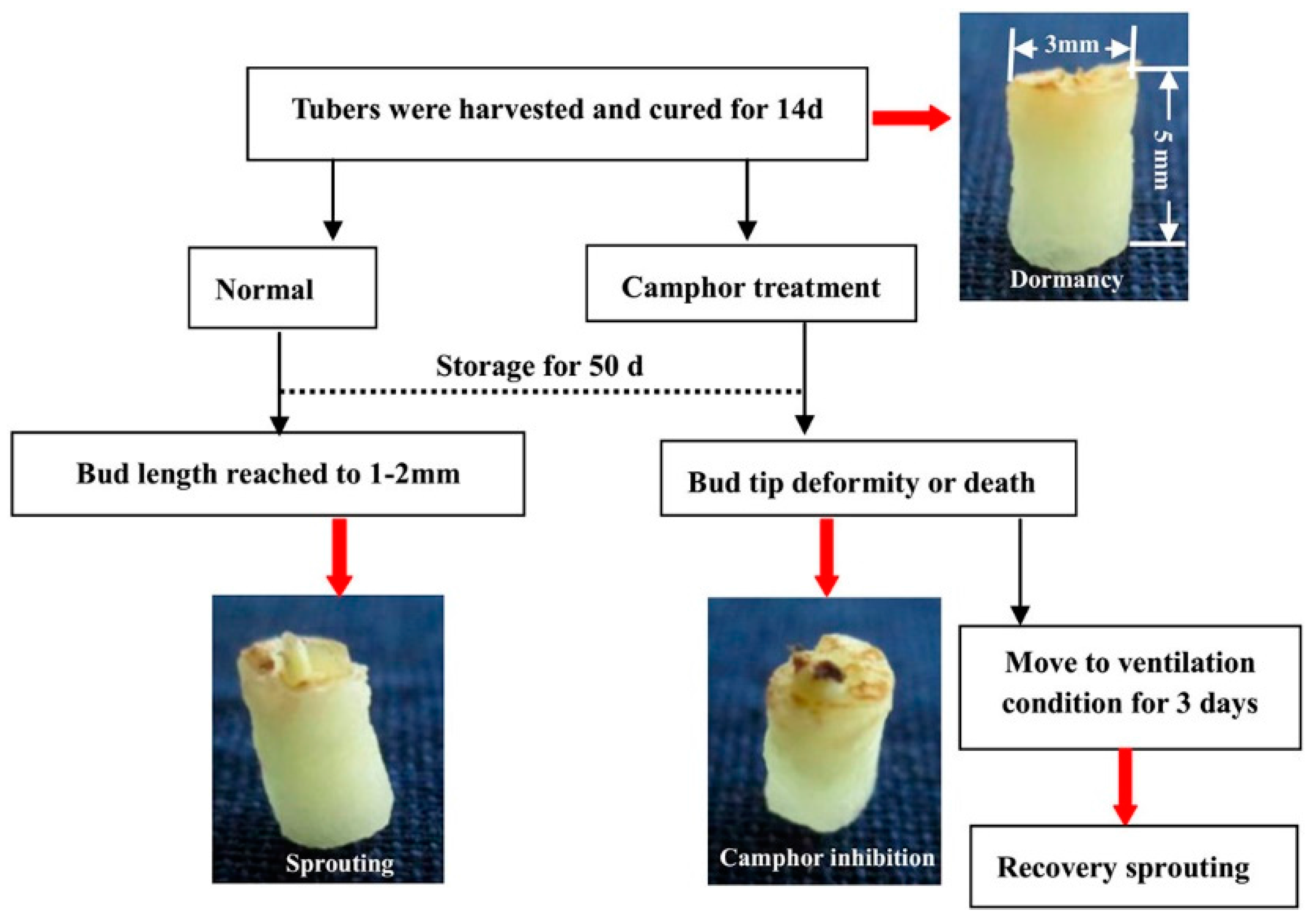
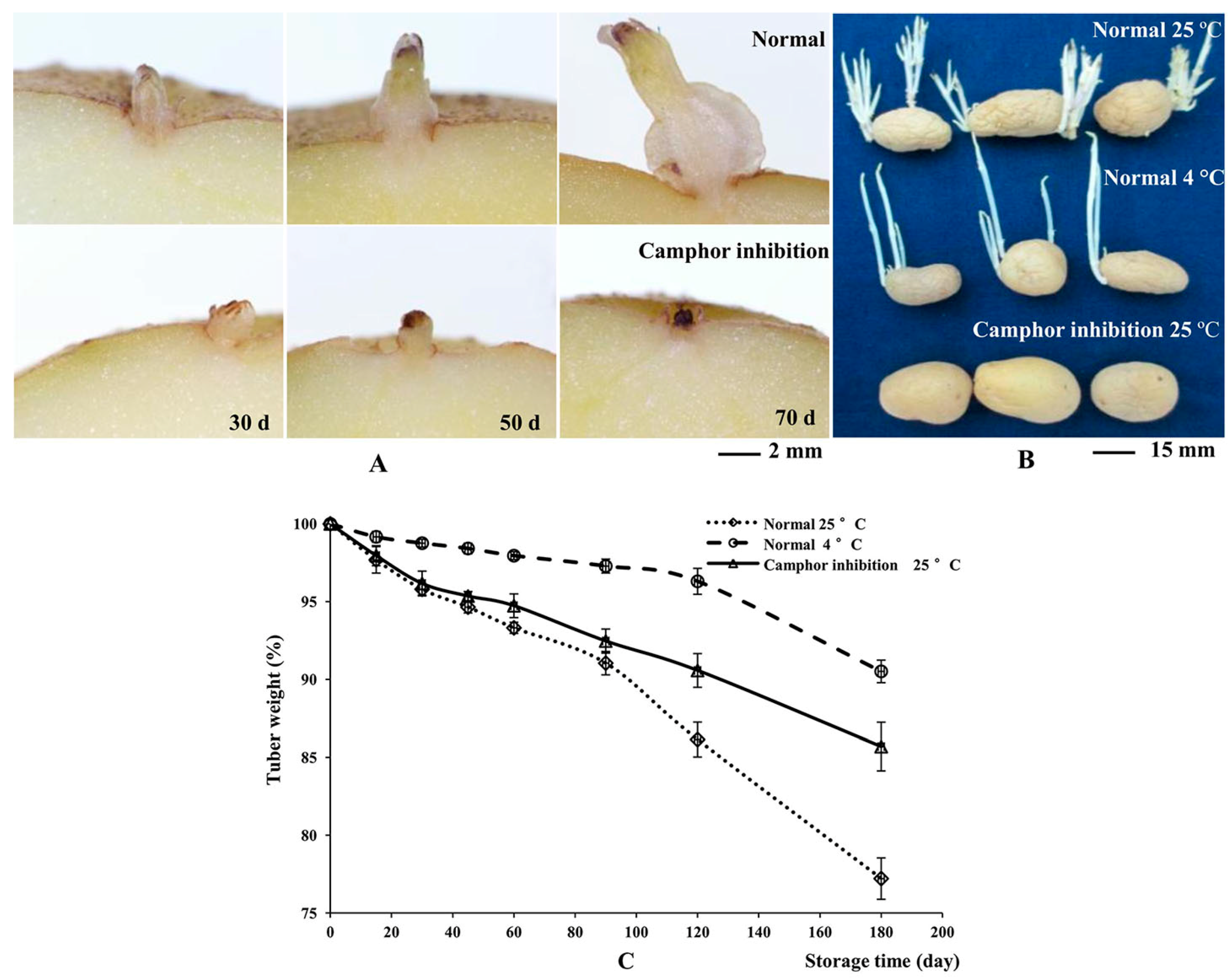
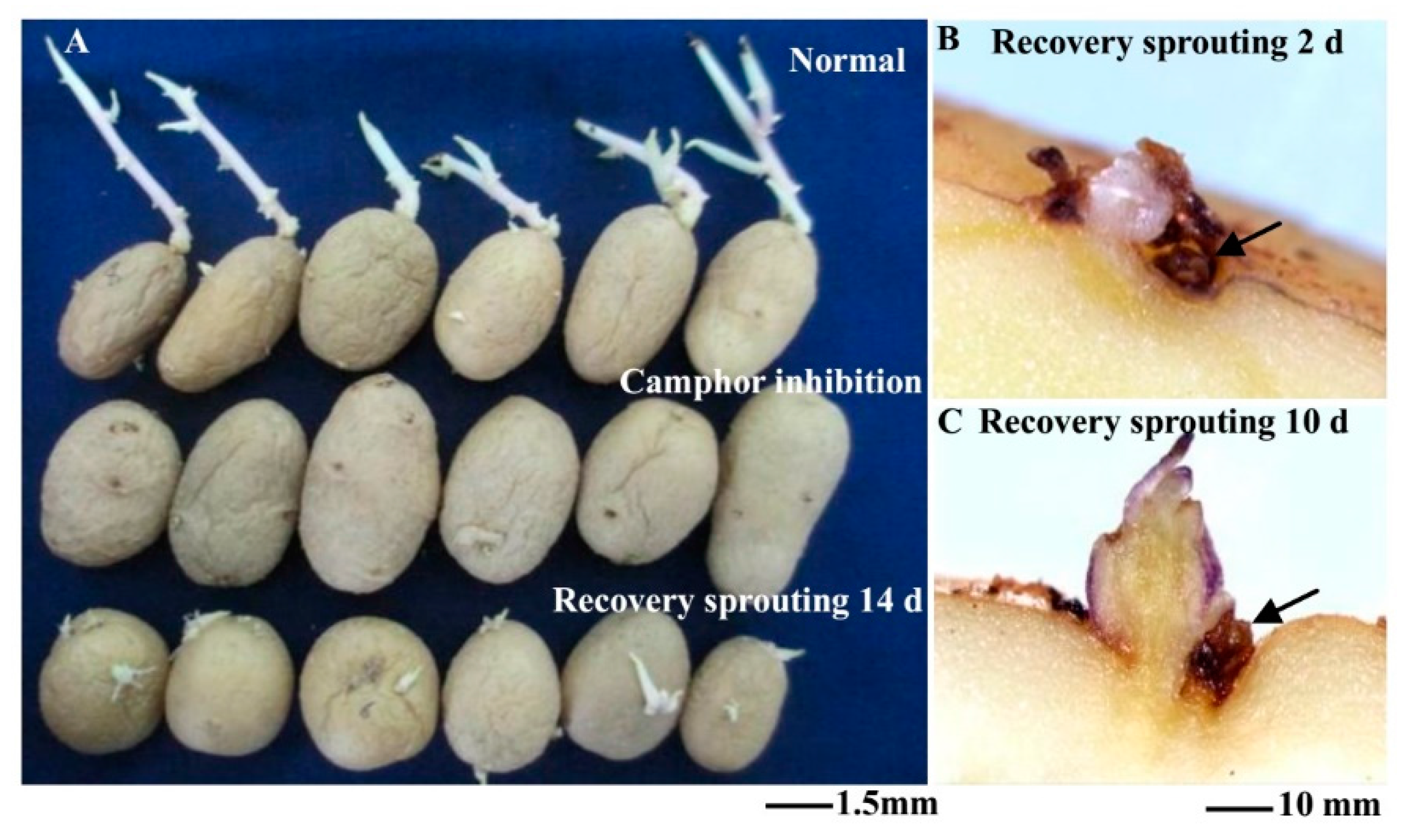
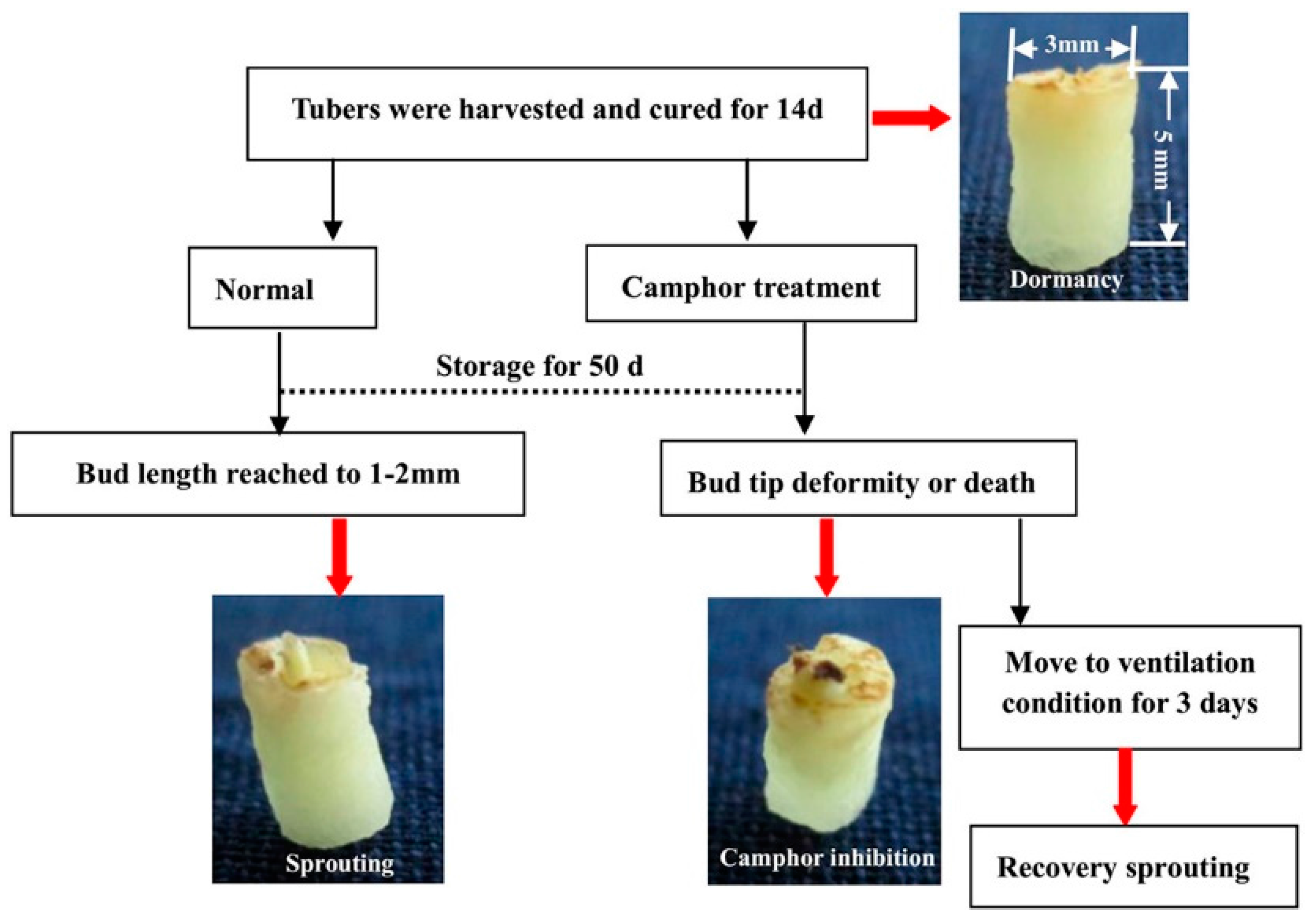
2.1. Flexible Inhibition of Tuber Sprouting Using Camphor
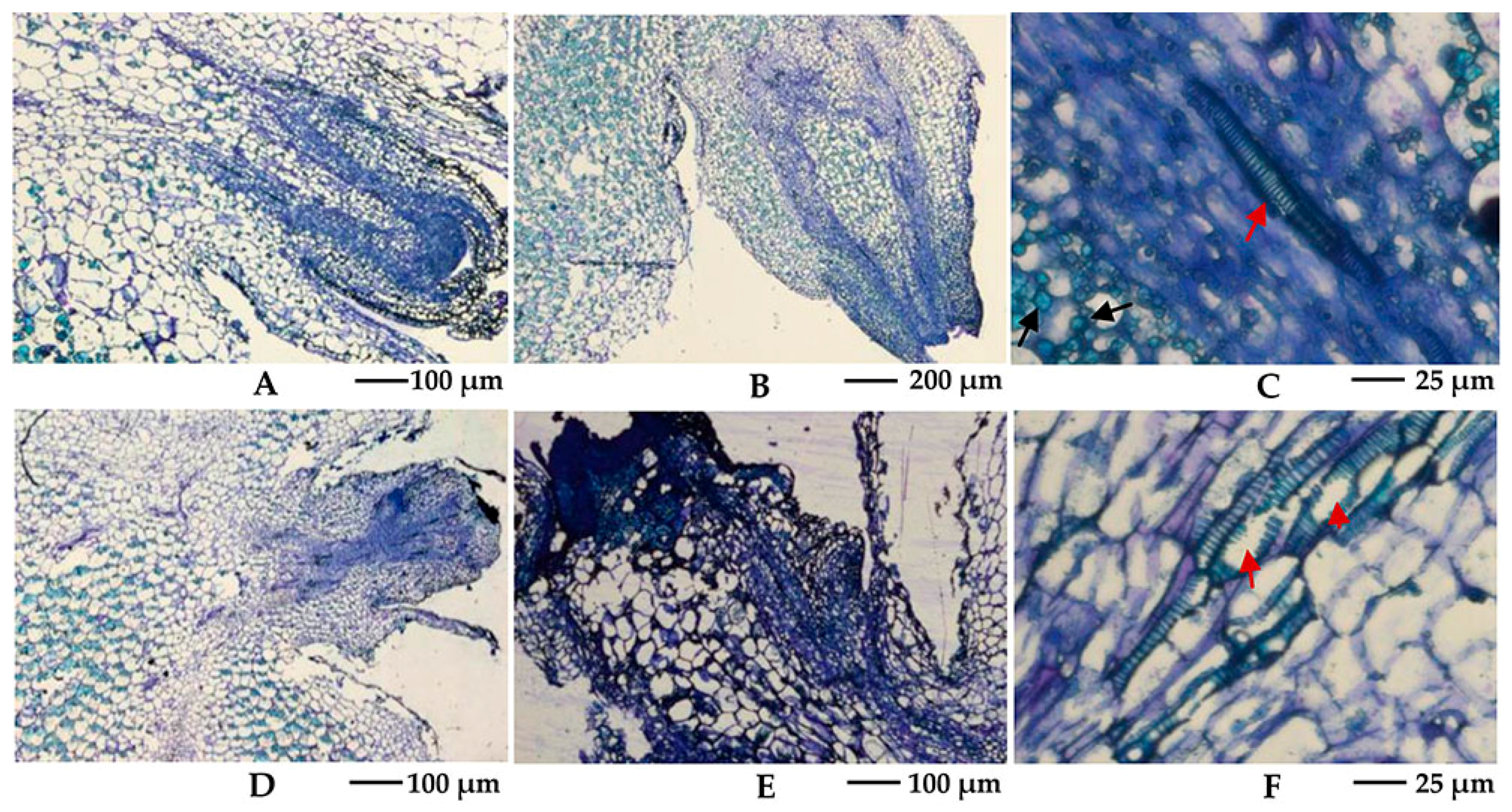
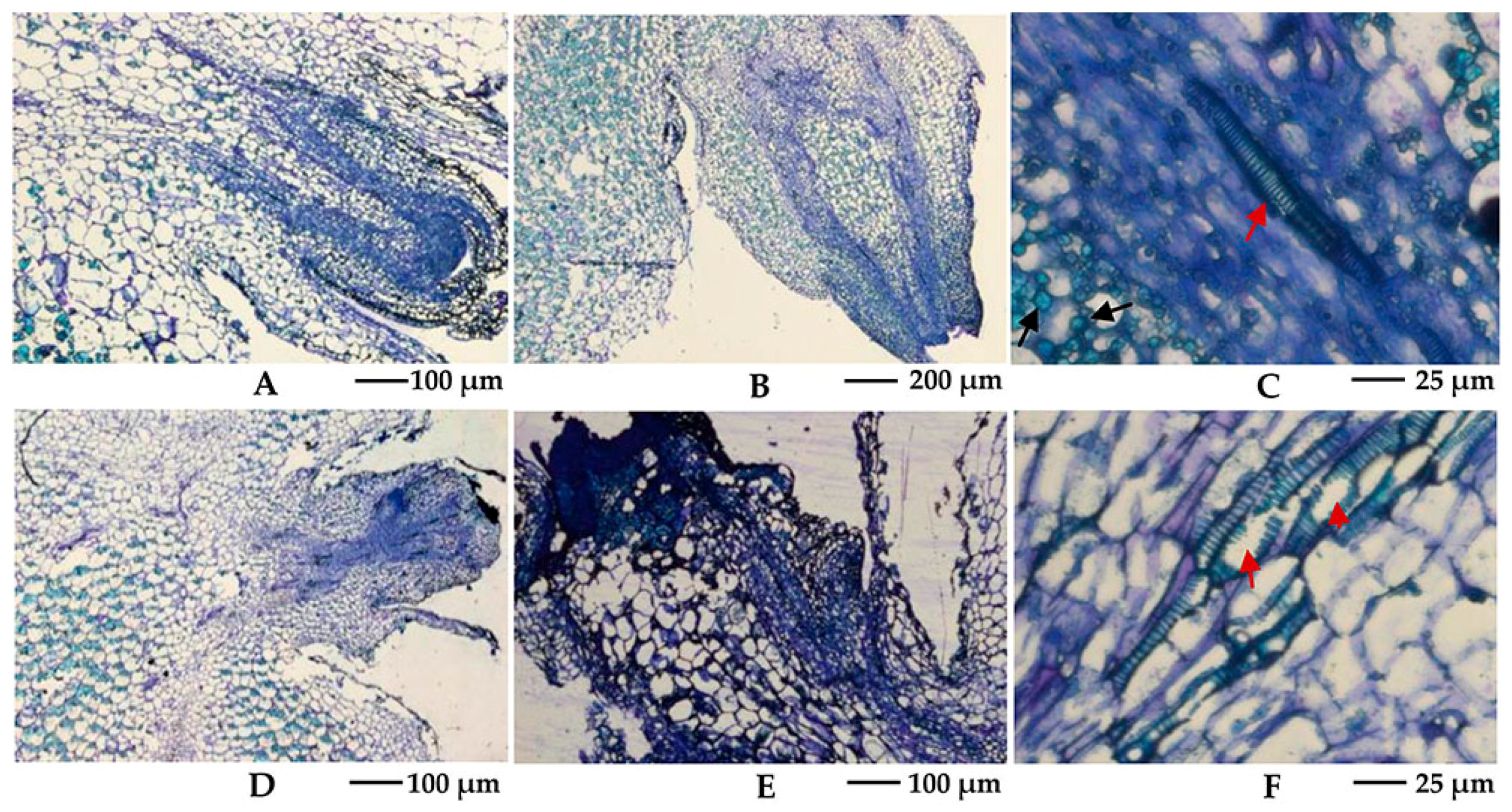
2.2. Effects of Camphor Inhibition on Histological Structures in the Bud Eye Region
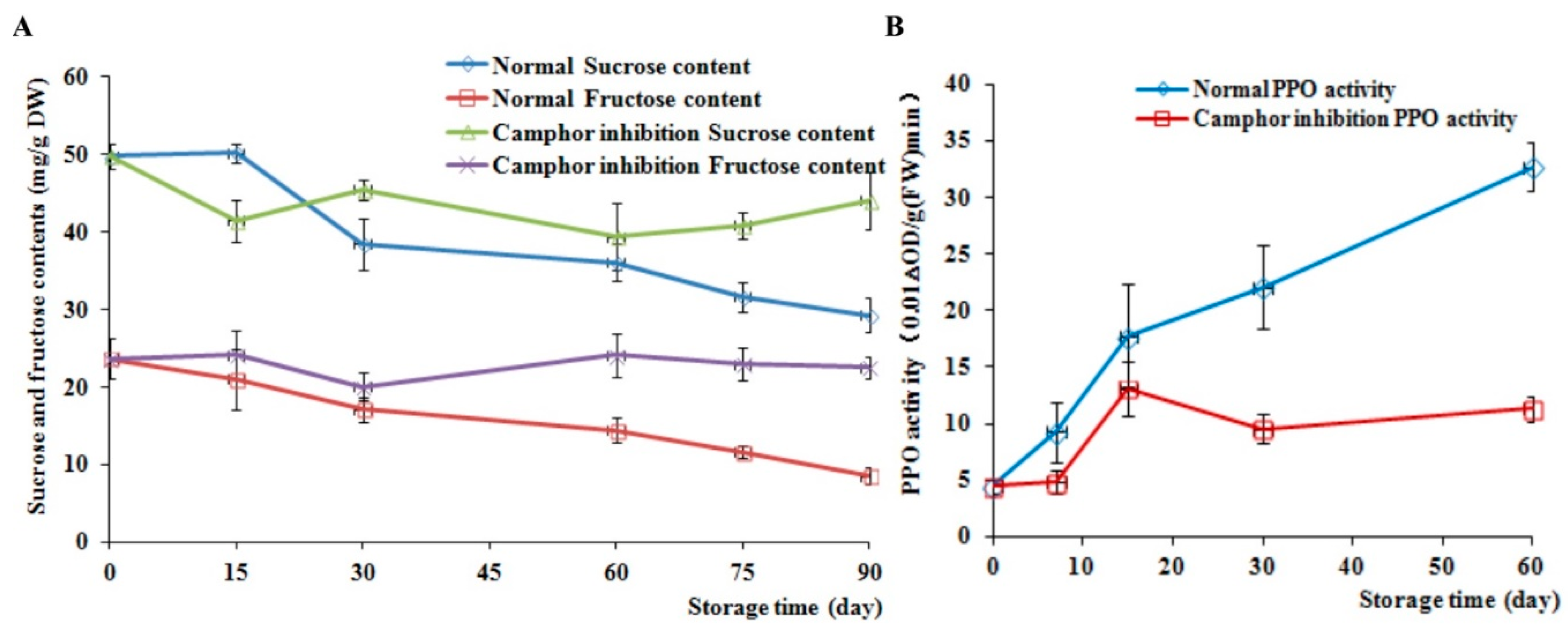
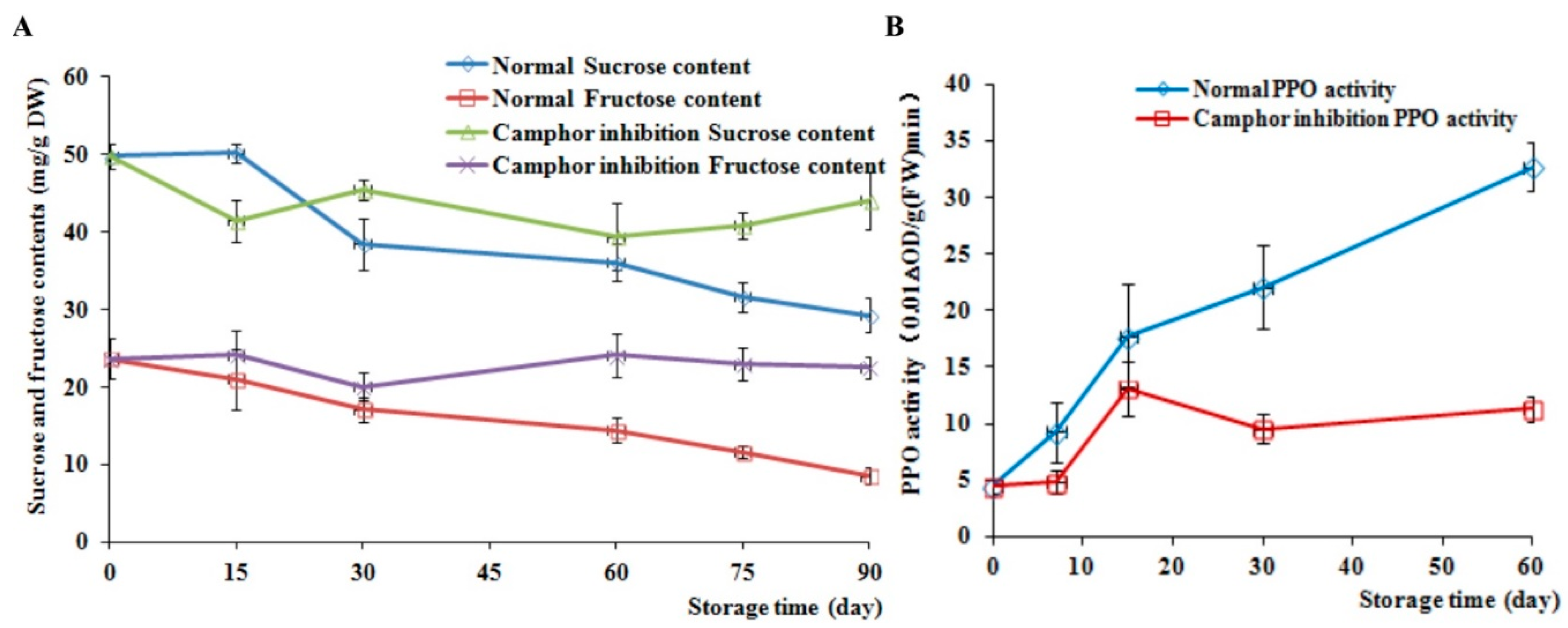
2.3. Analysis of Sucrose and Fructose Content and PPO Activity
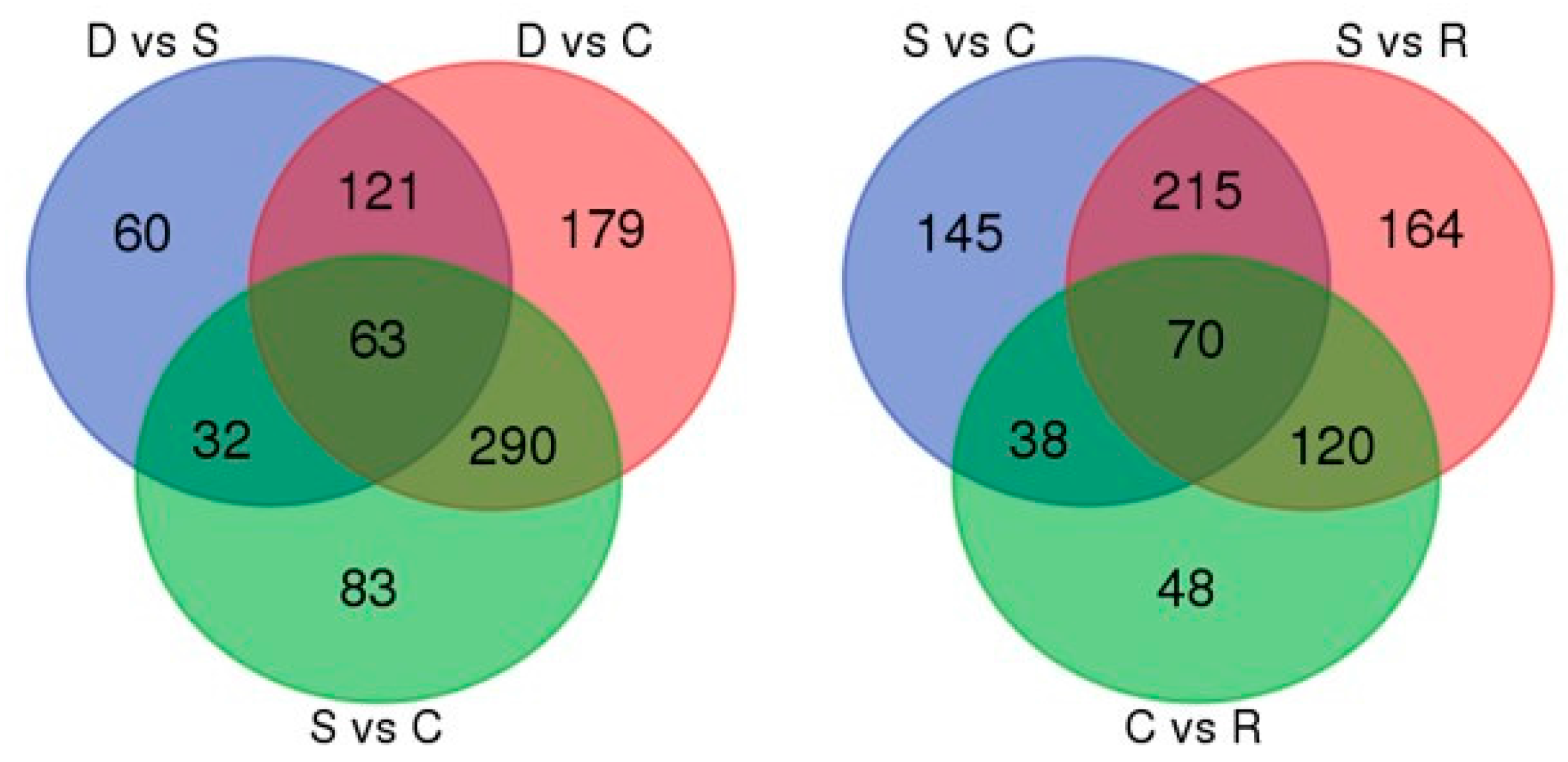
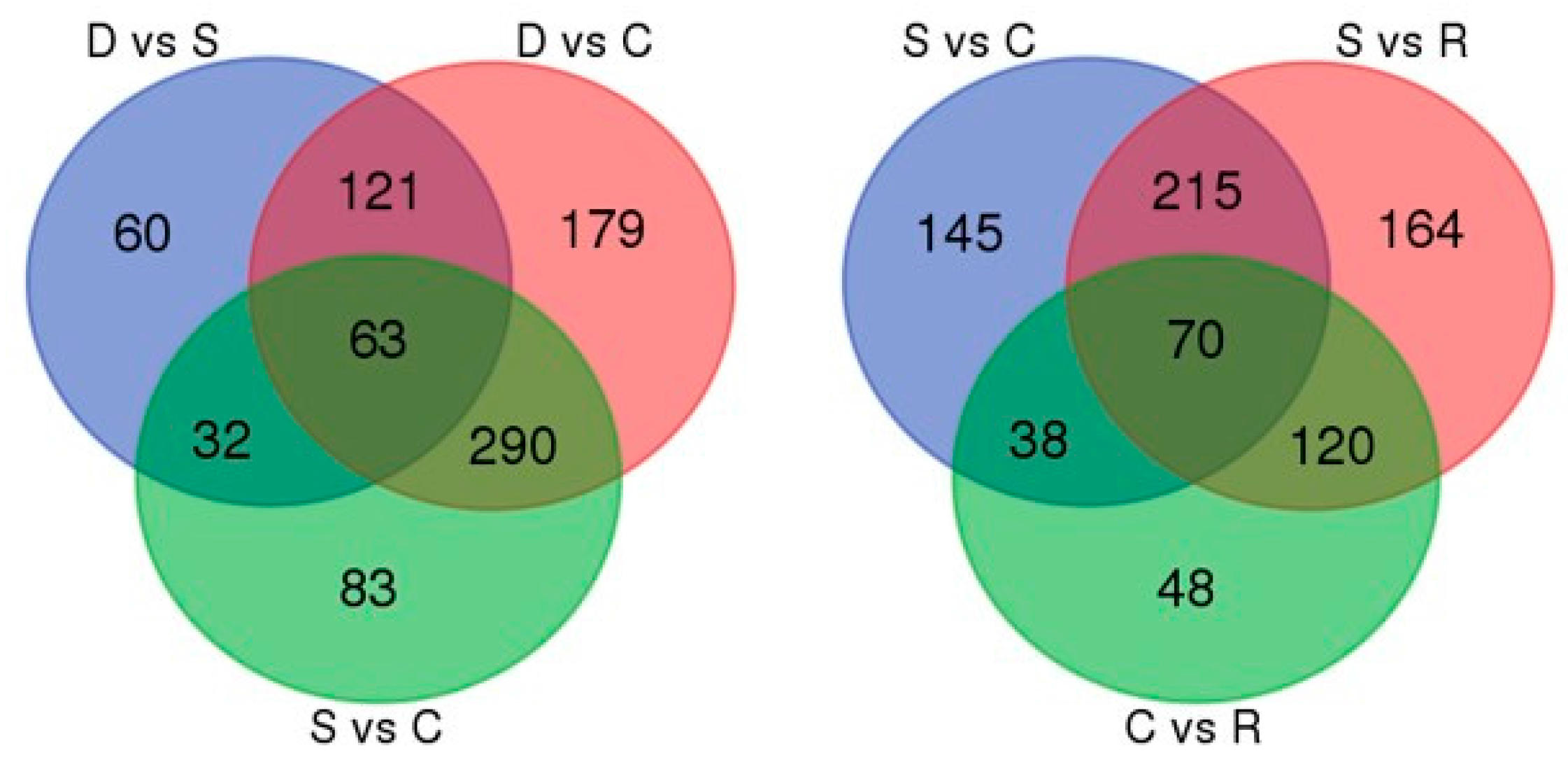
2.4. Evaluation of Differentially Transcribed Genes
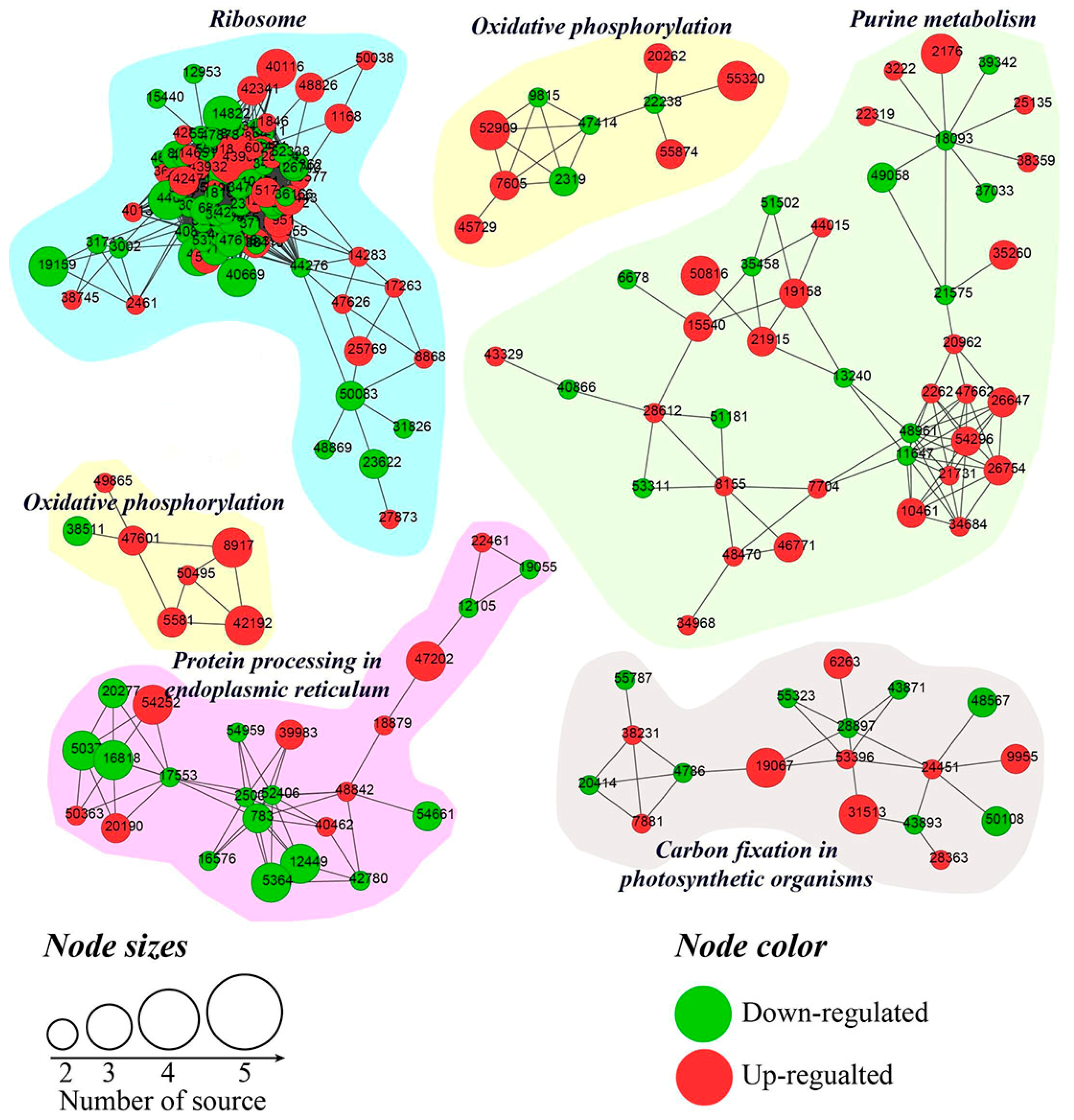
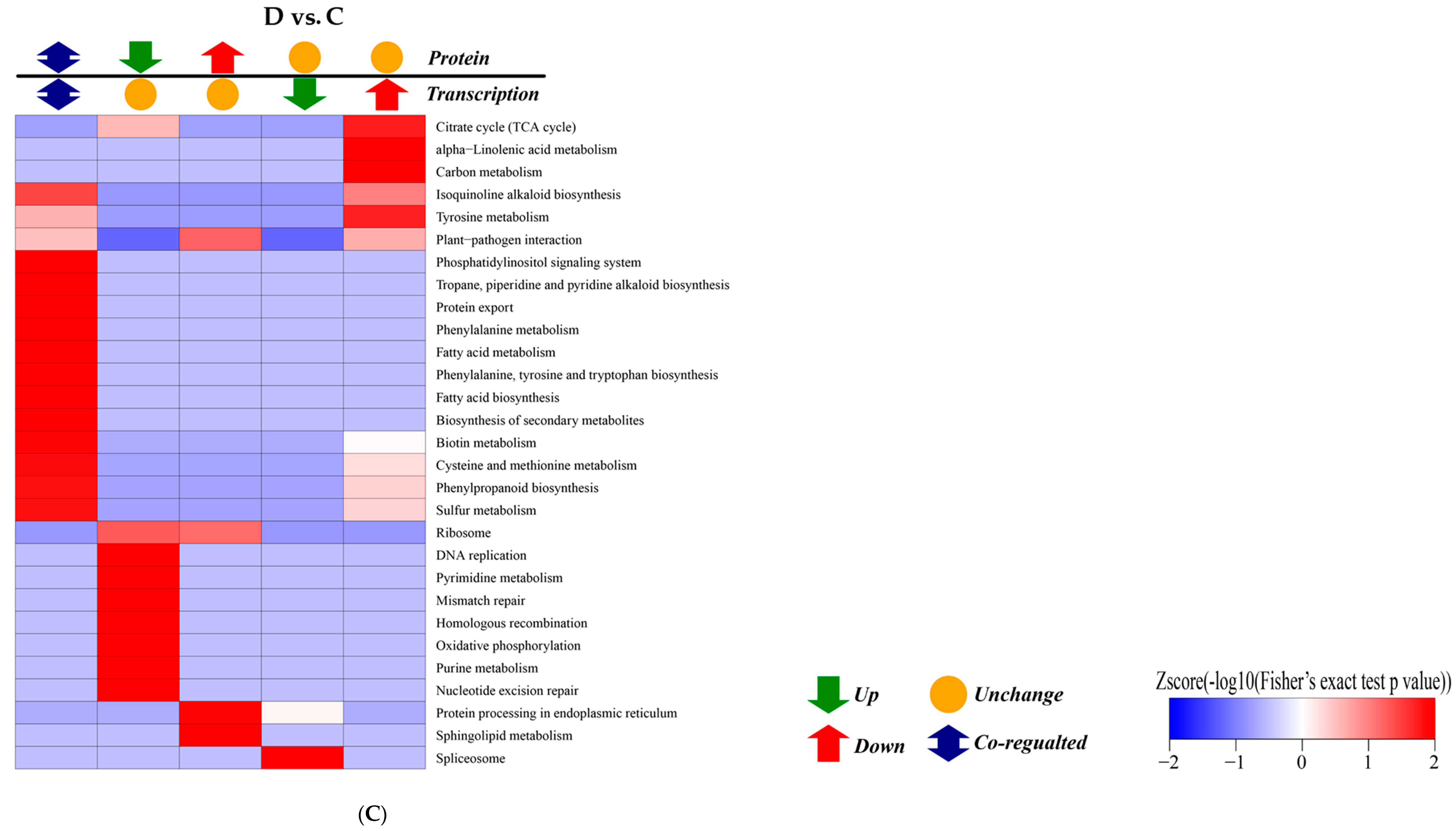
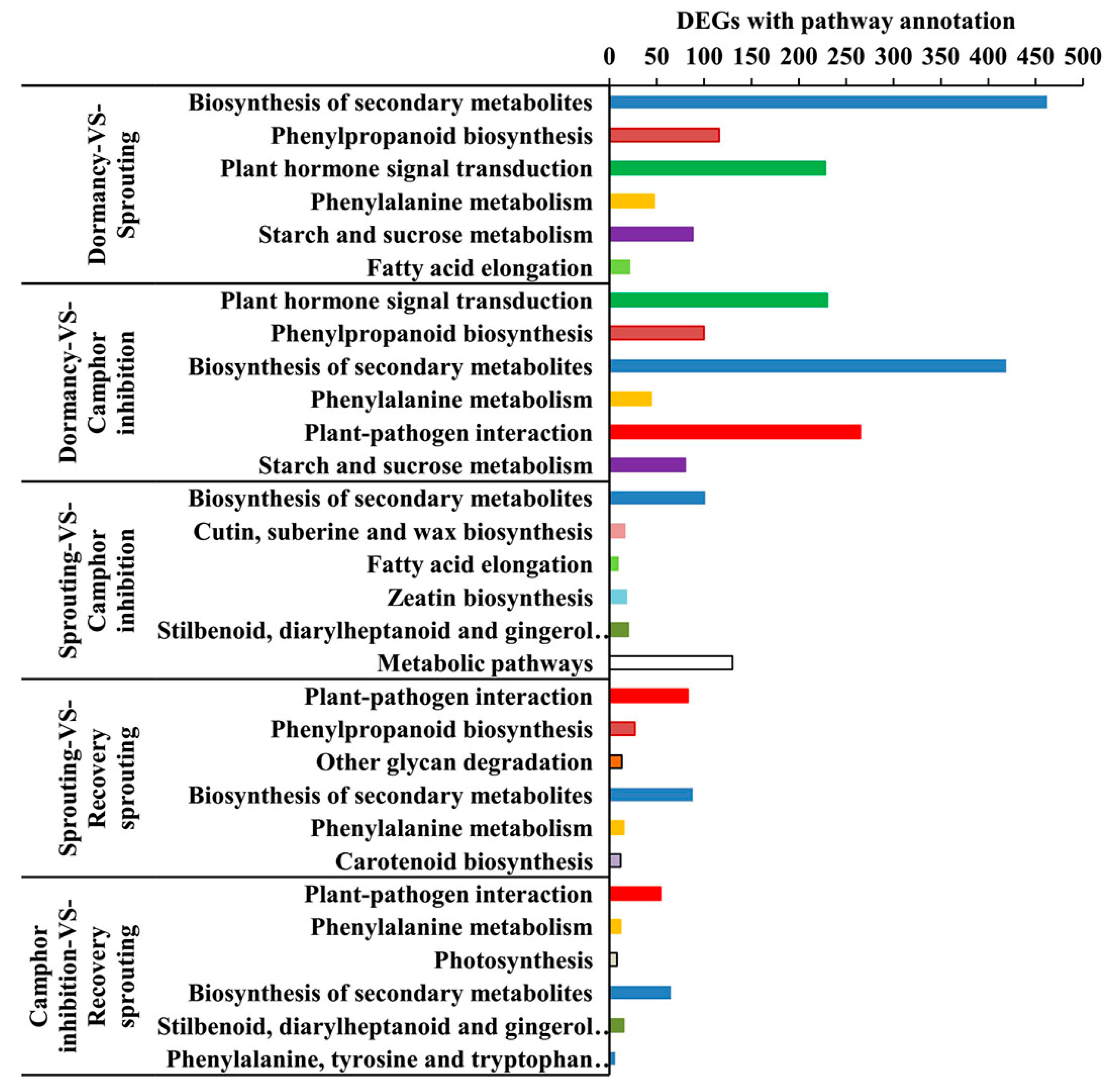
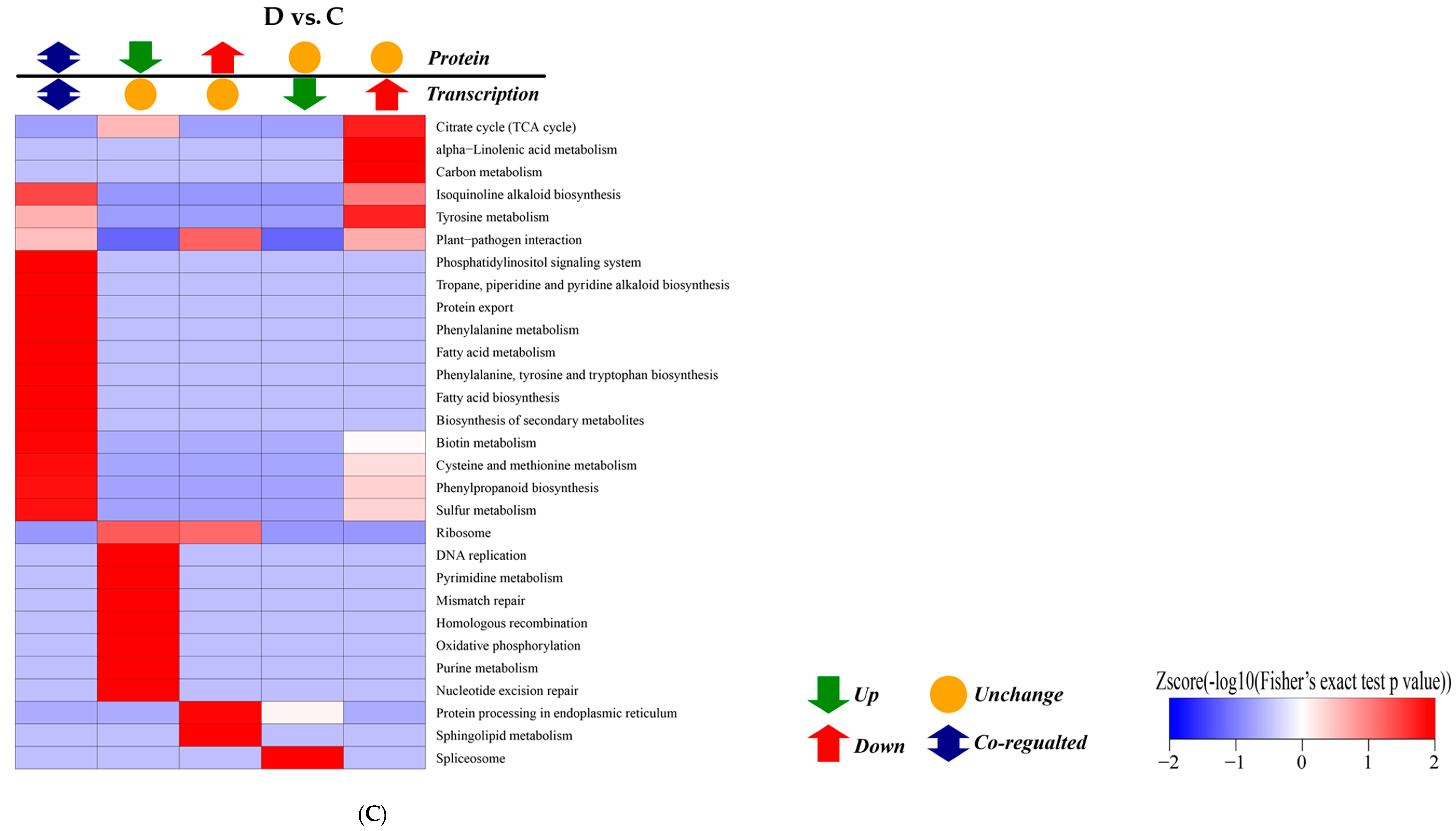
2.5. KEGG Pathway Enrichment Analysis of DEGs
2.6. Typical Expression Patterns of DEGs under Four Samples
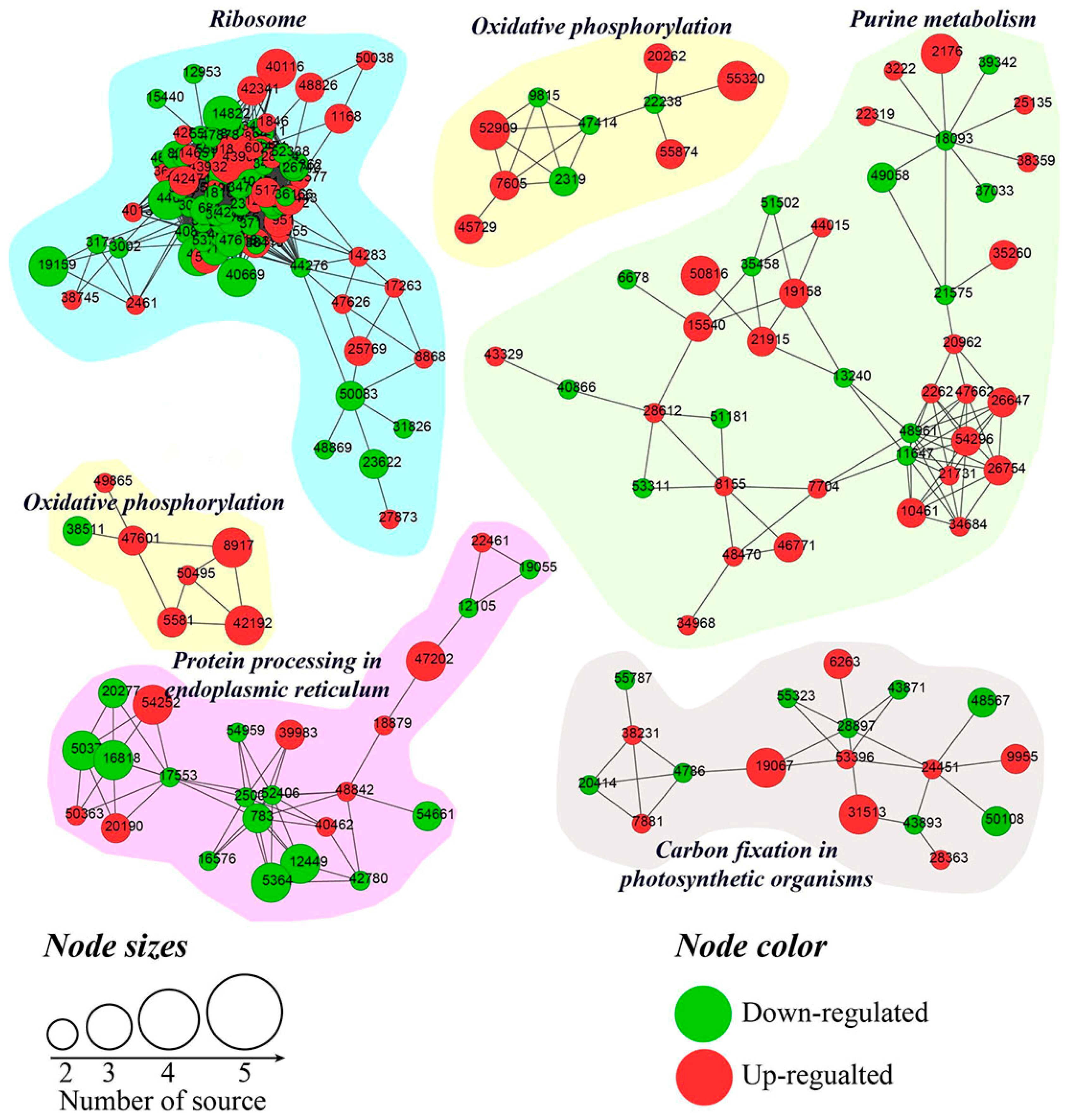
2.7. Analysis of Differentially Abundant Proteins (DAPs)
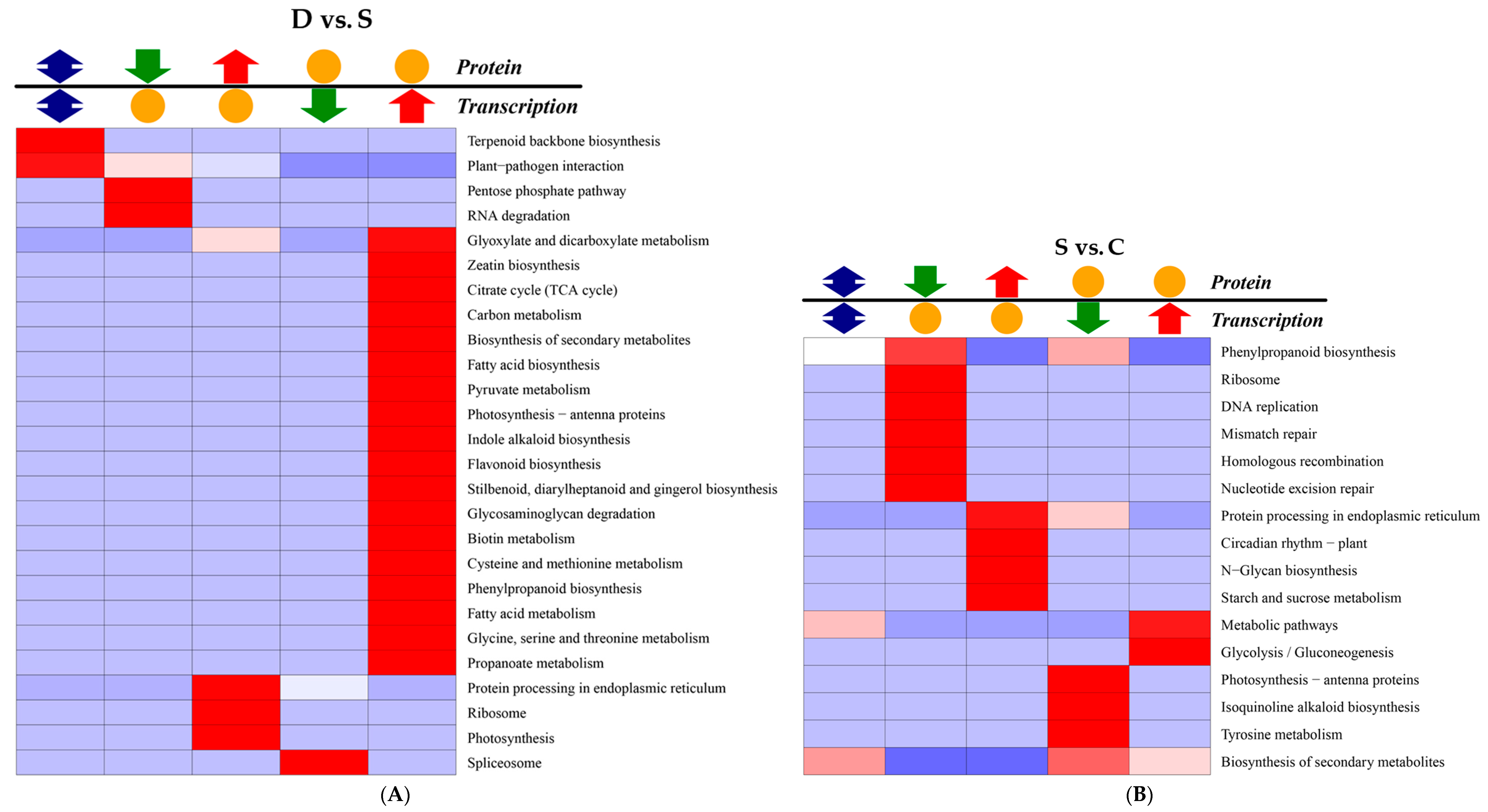
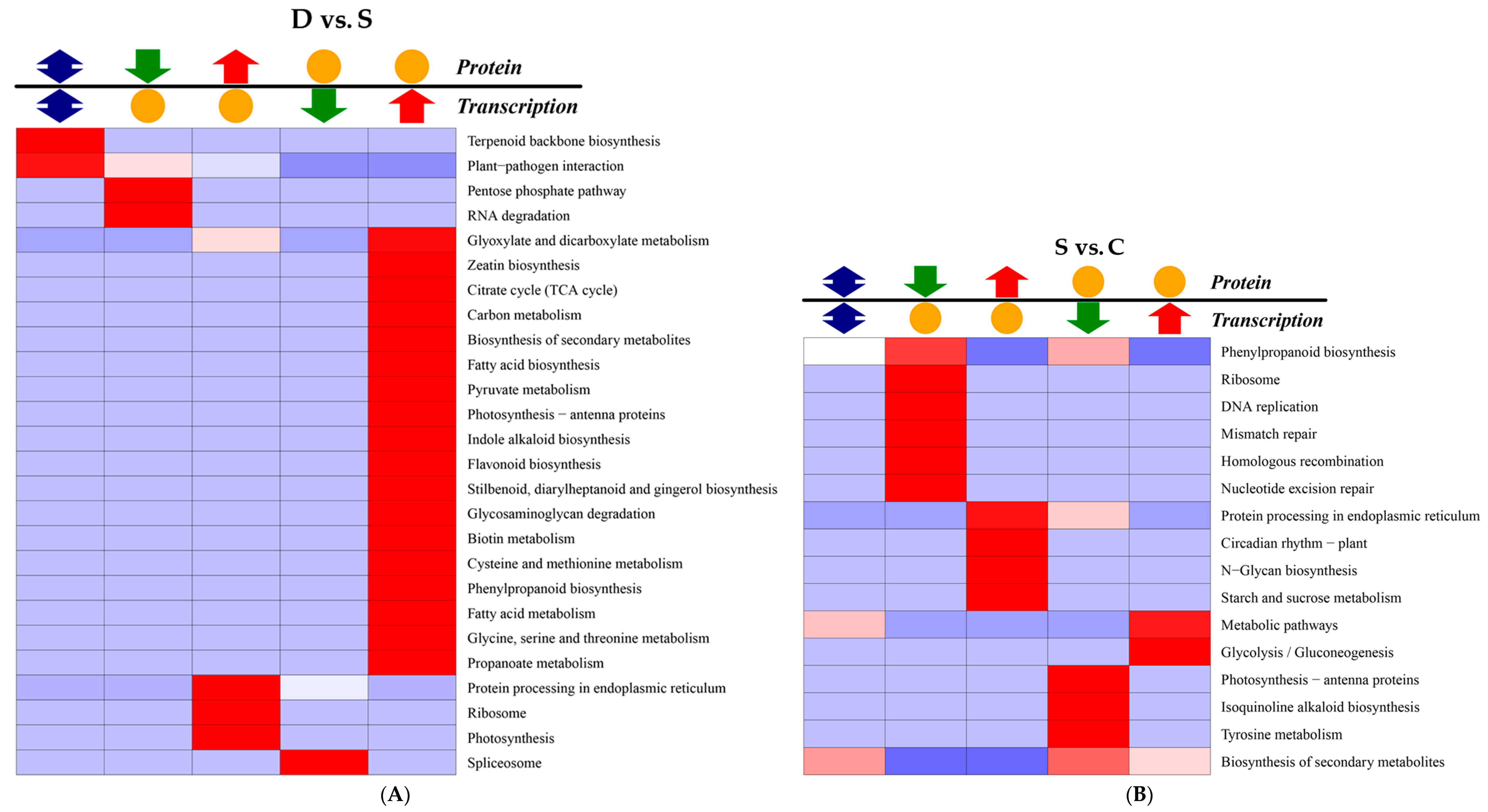
2.8. Correlation Analysis between Transcription and Proteomics
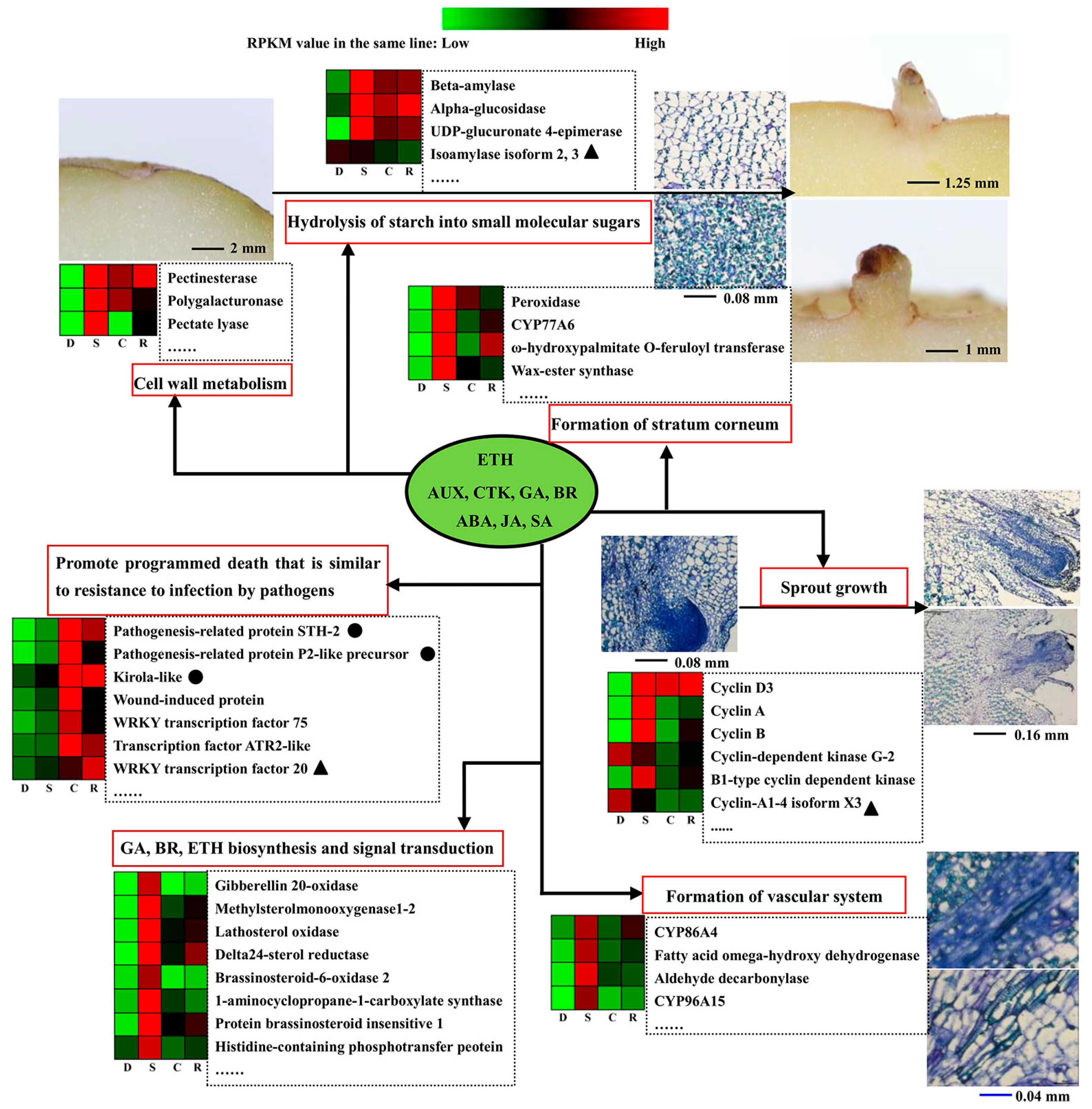
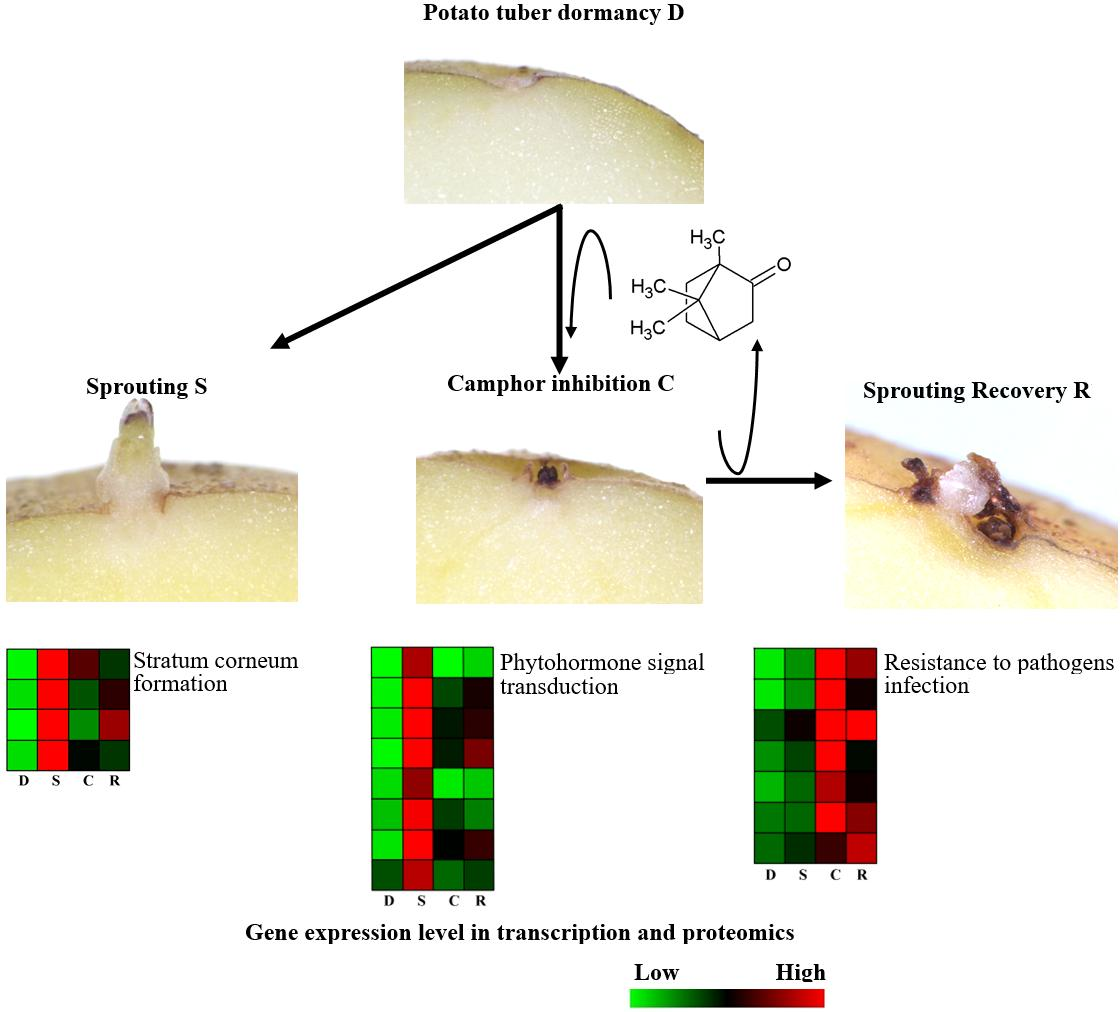
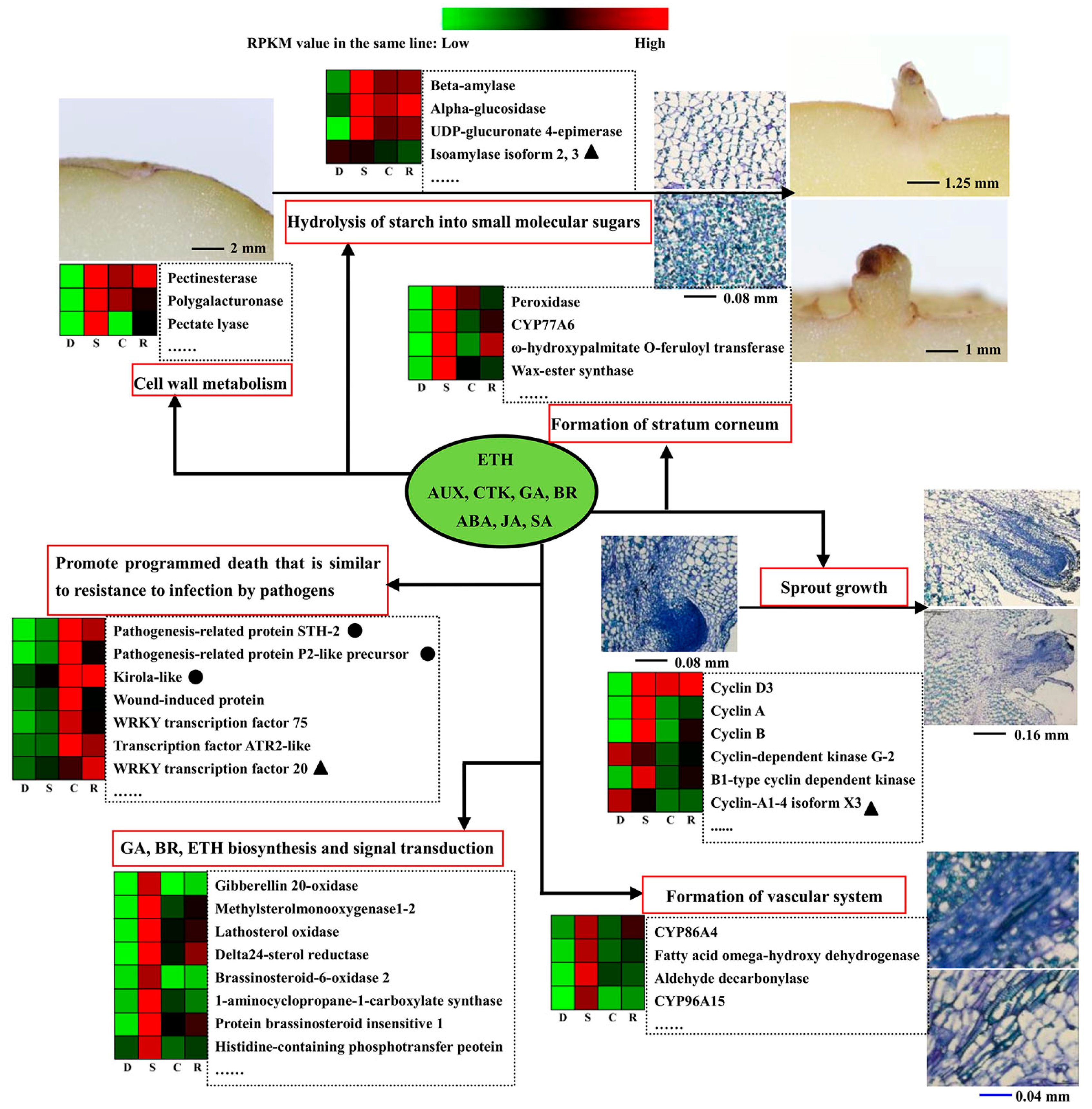
3. Discussion
3.1. Insight into the Transcription and Proteomics Analysis for Dormancy vs. Sprouting
3.2. Camphor Effect on Potato Tuber Sprouting
3.3. Camphor Effect on the Physiological State of Potato Tuber
3.4. Analysis on Uncoupling of Transcript Level and Protein Abundances
4. Materials and Methods
4.1. Plant Material Treatment and Physiological Index Analysis
4.2. Library Preparation and Sequencing
4.3. Identification of Different Expression Genes and Bioinformatics Analysis
4.4. Quantitative RT-PCR
4.5. Protein Extraction, Digestion, and iTRAQ Labelling
4.6. Liquid Chromatography-Mass Spectrometry and Database Search
4.7. Statistical Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Moll, A. Effect of the physiological age of seed tubers on growth and yield of potato cultivars of different maturity classes. Potato Res. 1985, 28, 233–250. [Google Scholar] [CrossRef]
- Hartmann, A.; Senning, M.; Hedden, P.; Sonnewald, U.; Sonnewald, S. Reactivation of meristem activity and sprout growth in potato tubers require both cytokinin and gibberellin. Plant Physiol. 2011, 155, 776–796. [Google Scholar] [CrossRef] [PubMed]
- Daniels-Lake, B.J.; Pruski, K.; Prange, R.K. Using ethylene gas and chlorpropham potato sprout inhibitors together. Potato Res. 2011, 54, 223–236. [Google Scholar] [CrossRef]
- Campbell, M.A.; Gleichsner, A.; Hilldorfer, L.; Horvath, D.; Suttle, J. The sprout inhibitor 1,4-dimethylnaphthalene induces the expression of the cell cycle inhibitors KRP1 and KRP2 in potatoes. Funct. Integr. Genom. 2012, 12, 533–541. [Google Scholar] [CrossRef] [PubMed]
- Weerd, D.J.W.; Thornton, M.K.; Shafii, B. Sprout suppressing residue levels of 1,4-dimethylnaphthalene (1,4DMN) in potato cultivars. Am. J. Potato Res. 2010, 87, 434–445. [Google Scholar] [CrossRef]
- Gómez-Castillo, D.; Cruz, E.; Iguaz, A.; Arroqui, C.; Vírseda, P. Effects of essential oils on sprout suppression and quality of potato cultivars. Postharvest Biol. Technol. 2013, 82, 15–21. [Google Scholar] [CrossRef]
- Hartmans, K.J. The use of carvone in agriculture: Sprout suppression of potatoes and antifungal activity against potato tuber and other plant diseases. Ind. Crops Prod. Appl. 1995, 4, 3–13. [Google Scholar] [CrossRef]
- Oosterhaven, K.; Poolman, B.; Smid, E.J. S-carvone as a natural potato sprout inhibiting, fungistatic and bacteristatic compound. Potato Res. 1995, 4, 23–31. [Google Scholar] [CrossRef]
- Bamnolker, T.P.; Dudai, N.; Fischer, R.; Belausov, E.; Zemach, H.; Shoseyov, O. Mint essential oil can induce or inhibit potato sprouting by differential alteration of apical meristem. Planta 2010, 232, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Guo, G.; Hu, X.; Zhang, Y.; Li, Q.; Li, R. Deep RNA sequencing at single base-pair resolution reveals high complexity of the rice transcriptome. Genome Res. 2010, 20, 646–654. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Cao, L.; Wang, S.; Li, Y.; Shi, X.; Liu, H.; Li, L.; Zhang, Z.; Fowke, L.C.; Wang, H.; et al. Down regulation of multiple CDK inhibitor ICK/KRP genes upregulates the E2F pathway and increases cell proliferation, and organ and seed sizes in Arabidopsis. Plant J. 2013, 75, 642–655. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Zhang, N.; Wen, Y.; Jin, X.; Yang, J.; Si, H. Transcriptomic changes during tuber dormancy release process revealed by RNA sequencing in potato. J. Biotechnol. 2015, 198, 17–30. [Google Scholar] [CrossRef] [PubMed]
- Wilm, M. Quantitative proteomics in biological research. Proteomics 2009, 9, 4590–4605. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Qiang, X.; Owsiany, K.; Zhang, S.; Thannhauser, T.W.; Li, L. Evaluation of different multidimensional LC-MS/MS pipelines for isobaric tags for relative and absolute quantitation (iTRAQ)-based proteomic analysis of potato tubers in response to cold storage. J. Proteome Res. 2011, 10, 4647–4660. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Zhang, N.; Zhao, S.; Chang, J.; Wang, Z.; Zhang, G. Proteomic changes during tuber dormancy release process revealed by iTRAQ quantitative proteomics in potato. Plant Physiol. Biochem. 2015, 86, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Mani, F.; Bettaieb, T.; Doudech, N.; Hannachi, C.; Institut, H.A.; Chott-Mariem, P.O.B. Physiological mechanisms for potato dormancy release and sprouting a review. Afr. Crop Sci. J. 2014, 22, 155–174. [Google Scholar]
- Sonnewald, S.; Sonnewald, U. Regulation of potato tuber sprouting. Planta 2014, 239, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Wilson, N.S.; Rueschhoff, E.E.; Bhatti, H.; Franks, R.G. Synergistic disruptions in seuss cyp85A2 double mutants reveal a role for brassinolide synthesis during gynoecium and ovule development. BMC Plant Biol. 2010, 10, 198. [Google Scholar]
- Reinecke, D.M.; Wickramarathna, A.D.; Ozga, J.A.; Kurepin, L.V.; Jin, A.L.; Good, A.G. Gibberellin 3-oxidase gene expression patterns influence gibberellin biosynthesis, growth, and development in pea. Plant Physiol. 2013, 163, 929–945. [Google Scholar] [CrossRef] [PubMed]
- Quan, J. The Role of Terpenoids in Plants and Its Application. Bot. Res. 2013, 106–108. [Google Scholar]
- Yang, X.; Wang, S.S.; Wang, M.; Qiao, Z.; Bao, C.C.; Zhang, W. Arabidopsis thaliana calmodulin-like protein CML24 regulates pollen tube growth by modulating the actin cytoskeleton and controlling the cytosolic Ca2+ concentration. Plant Mol. Biol. 2014, 86, 225–236. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Qiao, Z.; Liu, H.; Acharya, B.R.; Li, C.; Zhang, W. CML20, an Arabidopsis Calmodulin-like Protein, Negatively Regulates Guard Cell ABA Signaling and Drought Stress Tolerance. Front. Plant Sci. 2017, 8, 824. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Zhang, N.; Wen, Y.; Si, H.; Wang, D. Identification of differentially expressed genes in potato associated with tuber dormancy release. Mol. Biol. Rep. 2012, 39, 11277–11287. [Google Scholar] [CrossRef] [PubMed]
- Petrovova, M.; Tkadlec, J.; Dvoracek, L.; Streitova, E.; Licha, I. NAD(P)H-hydrate dehydratase- a metabolic repair enzyme and its role in Bacillus subtilis stress adaptation. PLoS ONE 2014, 9, e112590. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Olszewski, N.E. Gibberellin and abscisic acid regulate GAST1 expression at the level of transcription. Plant Mol. Biol. 1998, 38, 1053–1060. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, Z.; Xu, Y.; Joo, S.H.; Kim, S.K.; Xue, Z. OsGSR1 is involved in crosstalk between gibberellins and brassinosteroids in rice. Plant J. 2009, 57, 498–510. [Google Scholar] [CrossRef] [PubMed]
- Campbell, M.A.; Gleichsner, A.; Alsbury, R.; Horvath, D.; Suttle, J. The sprout inhibitors chlorpropham and 1,4-dimethylnaphthalene elicit different transcriptional profiles and do not suppress growth through a prolongation of the dormant state. Plant Mol. Biol. 2010, 73, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Fan, Z.J.; Liu, X.F.; Liu, F.L.; Bao, L.L.; Zhang, Y.G. Progress of researches on induced resistance of plant activator. Acta Phytophylacica Sin. 2005, 32, 87–92. [Google Scholar]
- Marineau, C.; Matton, D.P.; Brisson, N. Differential accumulation of potato tuber mRNAs during the hypersensitive response induced by arachidonic acid elicitor. Plant Mol. Biol. 1987, 9, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Constabel, C.P.; Brisson, N. The defense-related STH-2 gene product of potato shows race-specific accumulation after inoculation with low concentrations of Phytophthora infestans zoospores. Planta 1992, 188, 289–295. [Google Scholar] [CrossRef] [PubMed]
- Matton, D.P.; Prescott, G.; Bertrand, C.; Camirand, A.; Brisson, N. Identification of cis-acting elements involved in the regulation of the pathogenesis-related gene STH-2 in potato. Plant Mol. Biol. 1993, 22, 279–291. [Google Scholar] [CrossRef] [PubMed]
- Constabel, C.P.; Bertrand, C.; Brisson, N. Transgenic potato plants overexpressing the pathogenesis-related STH-2 gene show unaltered susceptibility to Phytophthora infestans and potato virus X. Plant Mol. Biol. 1993, 22, 775–782. [Google Scholar] [CrossRef] [PubMed]
- Rehman, K.U.M.; Sun, L.; Li, C.X.; Faheem, M.; Wang, W.; Tao, J.M. Comparative RNA-seq based transcriptomic analysis of bud dormancy in grape. BMC Plant Biol. 2017, 17, 18. [Google Scholar]
- Ibañes, M.; Fabregas, N.; Chory, J.; Cano-Delgado, A.I. Brassinosteroid signaling and auxin transport are required to establish the periodic pattern of Arabidopsis shoot vascular bundles. Proc. Natl. Acad. Sci. USA 2009, 106, 13630–13635. [Google Scholar] [CrossRef] [PubMed]
- Albert, M.; Werner, M.; Proksch, P.; Fry, S.C.; Kaldenhoff, R. The cell wall-modifying xyloglucan endotransglycosylase/hydrolase LeXTH1 is expressed during the defence reaction of tomato against the plant parasite Cuscuta reflexa. Plant Biol. 2004, 6, 402–407. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.K.; Ali, S.A.; Nath, P.; Sane, V.A. Activation of ethylene-responsive p-hydroxyphenylpyruvate dioxygenase leads to increased tocopherol levels during ripening in mango. J. Exp. Bot. 2011, 62, 3375–3385. [Google Scholar] [CrossRef] [PubMed]
- Kolattukudy, P.E. Biopolyester membranes of plants: Cutin and suberin. Science 1980, 208, 990–1000. [Google Scholar] [CrossRef] [PubMed]
- Garcia, CD.; Mazuch, J.; Altmann, T.; Mussig, C. EXORDIUM regulates brassinosteroid-responsive genes. FEBS Lett. 2004, 563, 82–86. [Google Scholar] [CrossRef]
- Rautengarten, C.; Usadel, B.; Neumetzler, L.; Hartmann, J.; Bussis, D.; Altmann, T. A subtilisin-like serine protease essential for mucilage release from Arabidopsis seed coats. Plant J. 2008, 54, 466–480. [Google Scholar] [CrossRef] [PubMed]
- Kopka, J.; Pical, C.; Gray, J.E.; Muller-Rober, B. Molecular and enzymatic characterization of three phosphoinositide-specific phospholipase C isoforms from potato. Plant Physiol. 1998, 116, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Swidzinski, J.A.; Leaver, C.J.; Sweetlove, L.J. A proteomic analysis of plant programmed cell death. Phytochemistry 2004, 65, 1829–1838. [Google Scholar] [CrossRef] [PubMed]
- Muers, M. Gene expression: Transcriptome to proteome and back to genome. Nat. Rev. Genet. 2011, 12, 518. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, R.N. Structure, catalytic mechanism, and evolution of the glutathione transferases. Chem. Res. Toxicol. 1997, 10, 2–18. [Google Scholar] [CrossRef] [PubMed]
- Pou, A.; Jeanguenin, L.; Milhiet, T.; Batoko, H.; Chaumont, F.; Hachez, C. Salinity-mediated transcriptional and post-translational regulation of the Arabidopsis aquaporin PIP2;7. Plant Mol. Biol. 2016, 92, 731–744. [Google Scholar] [CrossRef] [PubMed]
- Klein, R.D.; Chidawanyika, T.; Tims, H.S.; Meulia, T.; Bouchard, R.A.; Pett, V.B. Chaperone function of two small heat shock proteins from maize. Plant Sci. 2014, 221, 48–58. [Google Scholar] [CrossRef] [PubMed]
- Hu, G.J.; Koh, J.; Yoo, M.J.; Grupp, K.; Chen, S.; Wendel, J.F. Proteomic profiling of developing cotton fibers from wild and domesticated Gossypium barbadense. New Phytol. 2013, 200, 570–582. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Niwa, H.; Yokota, N.; Kubota, K.; Inoue, H. Widespread tissue expression of nepenthesis-like aspartic protease genes in Arabidopsis thaliana. Plant Physiol. Biol. 2008, 46, 724–729. [Google Scholar] [CrossRef] [PubMed]
- Guillaumot, D.; Guillon, S.; Deplanque, T.; Vanhee, C.; Gumy, C.; Masquelier, D.; Morsomme, P.; Batoko, H. The Arabidopsis TSPO-related protein is a stress and abscisic acid-regulated, endoplasmic reticulum-Golgi-localized membrane protein. Plant J. 2009, 60, 242–256. [Google Scholar] [CrossRef] [PubMed]
- Cui, N.; Song, Z.; Yang, B.; Fan, L.M. AtTSPO, a translocator protein, in stress responses in Arabidopsis. Environ. Exp. Bot. 2016, 129, 13–22. [Google Scholar] [CrossRef]
- Chen, J.; Liu, S.S.; Kohler, A.; Yan, B.; Luo, H.M.; Chen, X.M.; Guo, S.X. iTRAQ and RNA-Seq analyses provide new insights into regulation mechanism of symbiotic germination of Dendrobium officinale seeds (Orchidaceae). J. Proteome Res, 2017, 16, 2174–2187. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Wu, Z.; Tang, D.; Luo, K.; Lu, H.; Liu, Y. Comparative transcriptome analysis reveals critical function of sucrose metabolism related-enzymes in starch accumulation in the storage root of sweet potato. Front. Plant Sci. 2017, 8, 914. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; You, X.; Li, L.; Peng, H.; Su, W.; Li, C. Effects of a phospholipase D inhibitor on postharvest enzymatic browning and oxidative stress of litchi fruit. Postharvest Biol. Technol. 2011, 62, 288–294. [Google Scholar] [CrossRef]
- The Potato Genome Sequencing Consortium. Genome sequence and analysis of the tuber crop potato. Nature 2011, 475, 189–195. [Google Scholar]
- Mortazavi, A.; Williams, B.A.; McCue, K.; Schaeffer, L.; Wold, B. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods 2008, 5, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Seed Tuber | Number of Seeding | Number of Tubers | Tuber Weight (g) |
|---|---|---|---|
| Normal | 1.33 ± 0.49 B | 2.40 ± 0.83 B | 24.16 ± 5.43 a |
| Recovery sprouting | 2.86 ± 1.61 A | 4.14 ± 1.75 A | 19.38 ± 6.11 b |
| Gene ID | D | S | C | R | Functional Annotation | |
|---|---|---|---|---|---|---|
 | N16546 | 0.01 | 50.35 | 19.09 | 21.41 | GDSL(Gly-Asp-Ser-Leu)-like lipase/acylhydrolase family protein |
| N55828 | 0.01 | 23.67 | 12.36 | 18.00 | E3 ubiquitin-protein ligase RMA1H1 | |
| N88323 | 0.01 | 15.47 | 13.51 | 18.48 | Aspartic proteinase nepenthesin-1 | |
| N30676 | 11.32 | 1242.54 | 500.20 | 886.62 | 2-oxoglutarate-dependent dioxygenase | |
| N05343 | 0.98 | 251.85 | 155.24 | 183.27 | Pectinesterase | |
| N30650 | 1.42 | 209.91 | 103.84 | 159.95 | β-solanine/β-chaconine rhamnosyltransferase | |
 | N76367 | 0.12 | 34.15 | 0.01 | 1.09 | Proline-rich protein 1 |
| N83077 | 0.24 | 20.90 | 1.10 | 13.96 | Gip1 | |
| N24372 | 0.01 | 5.78 | 0.01 | 3.23 | Hypothetical gene of unknown function | |
| N23187 | 0.01 | 19.75 | 1.42 | 16.42 | Xyloglucan-specific endoglucanase inhibitor 4 | |
| N30708 | 0.01 | 24.63 | 2.95 | 6.81 | GATA-type transcription factor 4-like or AG-motif binding protein-3 | |
| N81314 | 3.23 | 40.78 | 6.27 | 9.18 | Endo-1,4-beta-glucanase | |
 | N37745 | 4.71 | 0.01 | 0.01 | 0.30 | Conserved gene of unknown function |
| N53578 | 4.23 | 0.58 | 0.59 | 1.87 | Conserved gene of unknown function | |
| N41670 | 4.69 | 0.69 | 1.13 | 1.20 | Serine/threonine-protein kinase SAPK1 | |
| N69883 | 43.11 | 9.27 | 11.40 | 6.10 | Protein phosphatase 2c | |
| N07511 | 35.47 | 12.23 | 8.31 | 6.28 | Dehydration-responsive element binding protein | |
| N04046 | 2353.85 | 939.44 | 881.62 | 1155.99 | Snakin-2 | |
 | N24747 | 5.25 | 4.81 | 0.01 | 1.86 | Hypothetical gene of unknown function |
| N05722 | 5.88 | 6.55 | 0.96 | 2.49 | Hypothetical gene of unknown function | |
| N35281 | 5.21 | 4.76 | 0.65 | 2.00 | UDP(Uridine diphosphate)-sugar:glycosyltransferase | |
| N76038 | 16.88 | 12.91 | 2.36 | 8.70 | Hypothetical gene of unknown function | |
| N40696 | 18.21 | 12.56 | 3.12 | 5.88 | Leucine-rich repeat resistance protein | |
| N47882 | 14.98 | 10.02 | 2.70 | 3.06 | Hypothetical gene of unknown function | |
 | N47338 | 0.01 | 0.01 | 23.80 | 4.11 | Wound-induced protein |
| N52057 | 0.01 | 0.15 | 9.11 | 0.44 | WRKY DNA binding protein | |
| N26225 | 0.38 | 0.87 | 26.82 | 10.63 | Conserved gene of unknown function | |
| N07870 | 0.90 | 3.96 | 40.06 | 6.74 | Osmotin | |
| N47343 | 5.63 | 9.02 | 89.09 | 26.79 | Wound-induced protein | |
| N03936 | 7.76 | 43.00 | 420.05 | 279.97 | Pathogenesis-related protein STH-2 or TSI-1 protein |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, L.-Q.; Zou, X.; Deng, M.-S.; Peng, J.; Huang, X.-L.; Lu, X.; Fang, C.-C.; Wang, X.-Y. Comparative Morphology, Transcription, and Proteomics Study Revealing the Key Molecular Mechanism of Camphor on the Potato Tuber Sprouting Effect. Int. J. Mol. Sci. 2017, 18, 2280. https://doi.org/10.3390/ijms18112280
Li L-Q, Zou X, Deng M-S, Peng J, Huang X-L, Lu X, Fang C-C, Wang X-Y. Comparative Morphology, Transcription, and Proteomics Study Revealing the Key Molecular Mechanism of Camphor on the Potato Tuber Sprouting Effect. International Journal of Molecular Sciences. 2017; 18(11):2280. https://doi.org/10.3390/ijms18112280
Chicago/Turabian StyleLi, Li-Qin, Xue Zou, Meng-Sheng Deng, Jie Peng, Xue-Li Huang, Xue Lu, Chen-Cheng Fang, and Xi-Yao Wang. 2017. "Comparative Morphology, Transcription, and Proteomics Study Revealing the Key Molecular Mechanism of Camphor on the Potato Tuber Sprouting Effect" International Journal of Molecular Sciences 18, no. 11: 2280. https://doi.org/10.3390/ijms18112280