Expression Profile of Genes Regulating Steroid Biosynthesis and Metabolism in Human Ovarian Granulosa Cells—A Primary Culture Approach

 , , , and
, , , and
Abstract
:1. Introduction
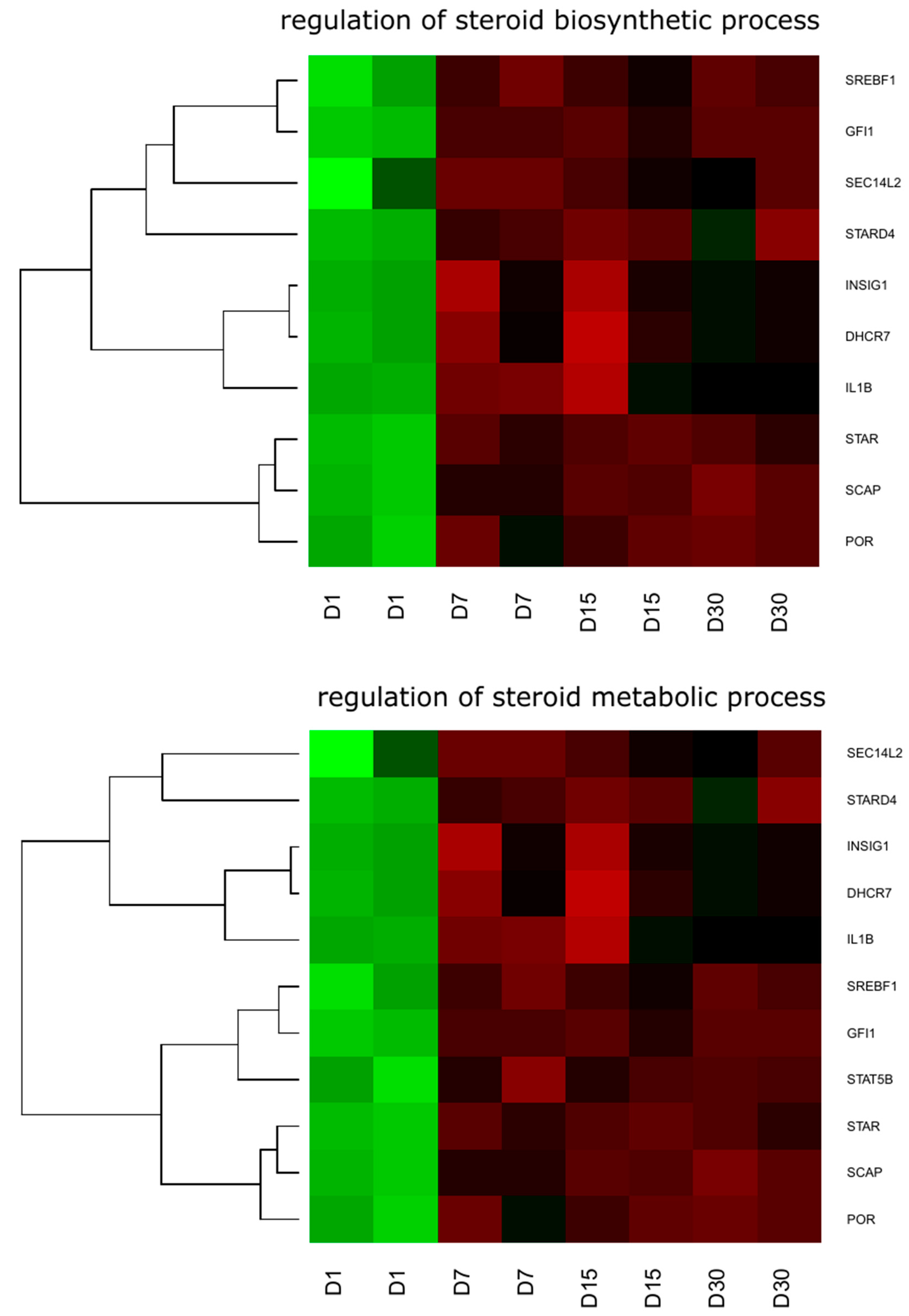
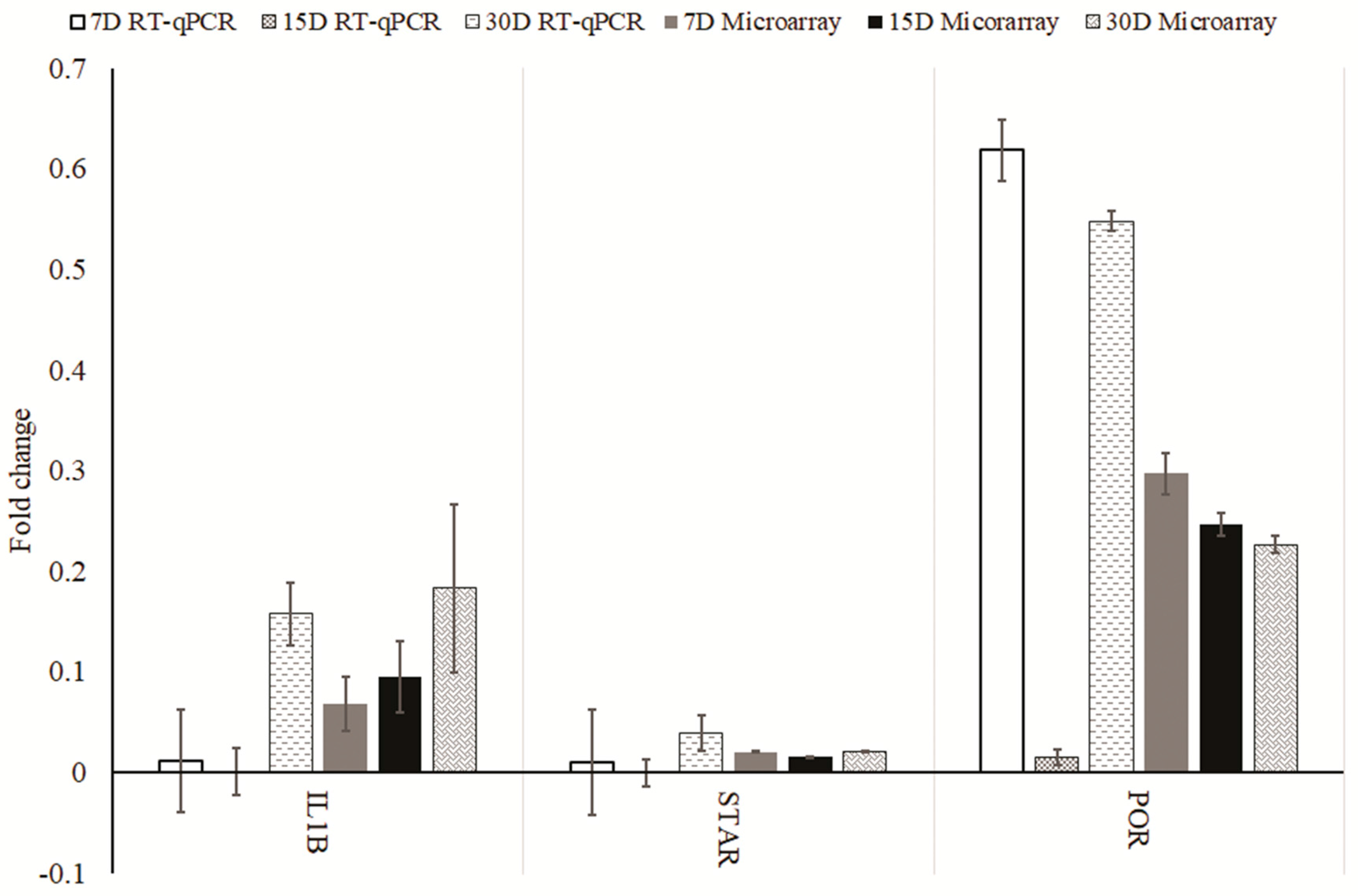
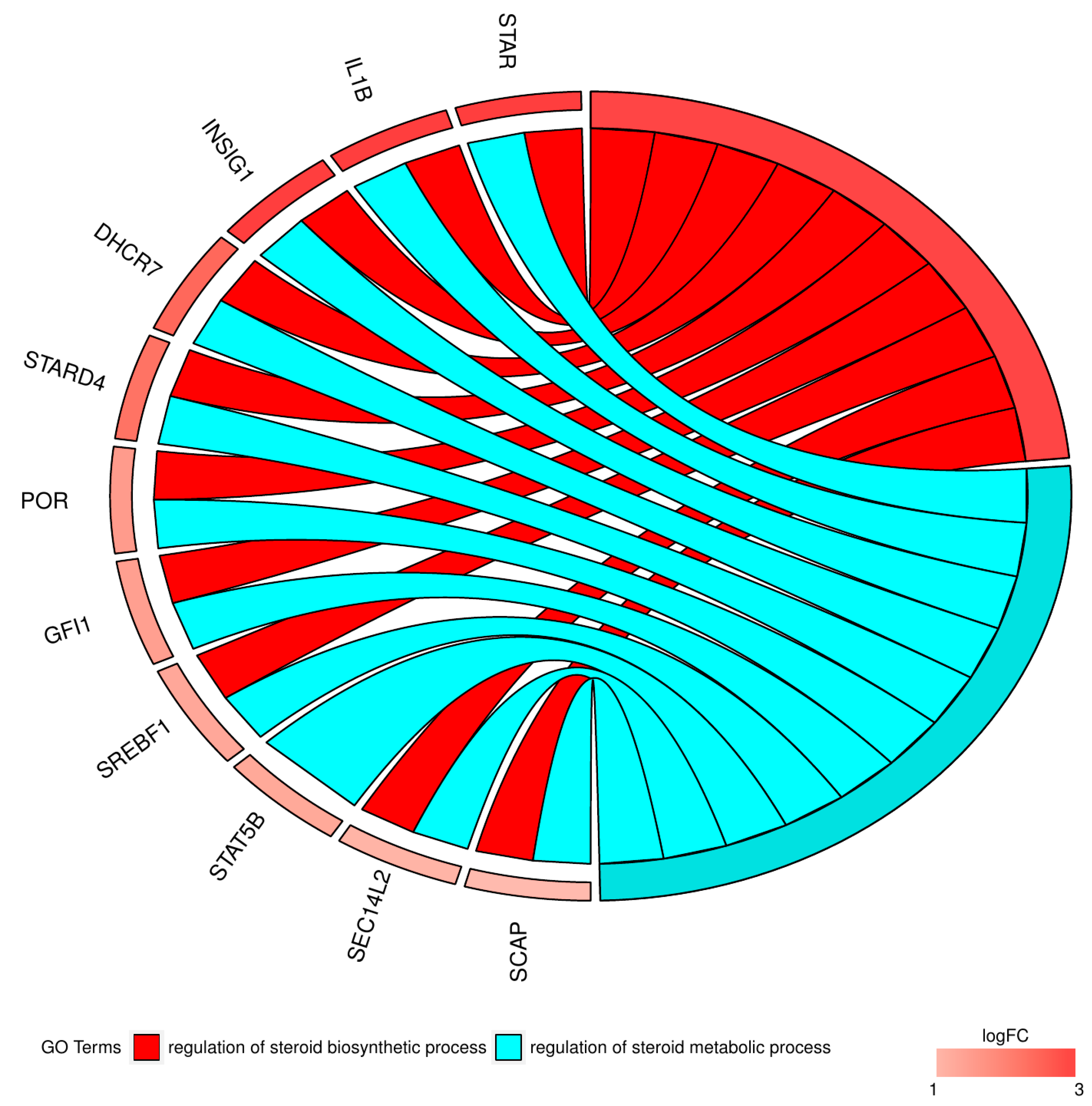
2. Results
3. Discussion
4. Material and Methods
4.1. Patients and Collection of Granulosa Cells
4.2. Primary Cell Culture
4.3. Total RNA Isolation
4.4. Microarray Expression Analysis and Statistics
4.5. Real-Time Quantitative Polymerase Chain Reaction (RT-qPCR) Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Kranc, W.; Budna, J.; Kahan, R.; Chachuła, A.; Bryja, A.; Ciesiółka, S.; Borys, S.; Antosik, M.P.; Bukowska, D.; Brussow, K.P.; et al. Molecular Basis of Growth, Proliferation, and Differentiation of Mammalian Follicular Granulosa Cells. J. Biol. Regul. Homeost. Agents 2017, 31, 1–8. [Google Scholar] [PubMed]
- Kranc, W.; Budna, J.; Dudek, M.; Bryja, A.; Chachuła, A.; Ciesiółka, S.; Borys, S.; Dyszkiewicz-Konwińska, M.; Jeseta, M.; Porowski, L.; et al. The Origin, in Vitro Differentiation, and Stemness Specificity of Progenitor Cells. J. Biol. Regul. Homeost. Agents 2017, 31, 365–369. [Google Scholar] [PubMed]
- Budna, J.; Celichowski, P.; Karimi, P.; Kranc, W.; Bryja, A.; Ciesiółka, S.; Rybska, M.; Borys, S.; Jeseta, M.; Bukowska, D.; et al. Does Porcine Oocytes Maturation in Vitro Is Regulated by Genes Involved in Transforming Growth Factor Beta Receptor Signaling Pathway? Adv. Cell Biol. 2017, 5, 1–14. [Google Scholar] [CrossRef]
- Kranc, W.; Celichowski, P.; Budna, J.; Khozmi, R.; Bryja, A.; Ciesiółka, S.; Rybska, M.; Borys, S.; Jeseta, M.; Bukowska, D.; et al. Positive Regulation Of Macromolecule Metabolic Process Belongs To The Main Mechanisms Crucial For Porcine Ooocytes Maturation. Adv. Cell Biol. 2017, 5, 15–31. [Google Scholar] [CrossRef]
- Nawrocki, M.J.; Budna, J.; Celichowski, P.; Khozmi, R.; Bryja, A.; Kranc, W.; Borys, S.; Ciesiółka, S.; Knap, S.; Jeseta, M.; et al. Analysis of Fructose and Mannose—Regulatory Peptides Signaling Pathway in Porcine Epithelial Oviductal Cells (OECs) Primary Cultured Long-Term in Vitro. Adv. Cell Biol. 2017, 5, 129–135. [Google Scholar] [CrossRef]
- Ciesiółka, S.; Budna, J.; Jopek, K.; Bryja, A.; Kranc, W.; Borys, S.; Jeseta, M.; Chachuła, A.; Ziółkowska, A.; Antosik, P.; et al. Time- and Dose-Dependent Effects of 17 Beta-Estradiol on Short-Term, Real-Time Proliferation and Gene Expression in Porcine Granulosa Cells. Biomed. Res. Int. 2017, 2017, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.-G.; Barad, D.H.; Kushnir, V.A.; Lazzaroni, E.; Wang, Q.; Albertini, D.F.; Gleicher, N. Aging-Related Premature Luteinization of Granulosa Cells Is Avoided by Early Oocyte Retrieval. J. Endocrinol. 2015, 226, 167–180. [Google Scholar] [CrossRef] [PubMed]
- Borys, S.; Khozmi, R.; Kranc, W.; Bryja, A.; Dyszkiewicz-Konwińska, M.; Jeseta, M.; Kempisty, B. Recent Findings of the Types of Programmed Cell Death. Adv. Cell Biol. 2017, 5, 43–49. [Google Scholar] [CrossRef]
- Ciesiółka, S.; Bryja, A.; Budna, J.; Kranc, W.; Chachuła, A.; Bukowska, D.; Piotrowska, H.; Porowski, L.; Antosik, P.; Bruska, M.; et al. Epithelialization and Stromalization of Porcine Follicular Granulosa Cells during Real-Time Proliferation—A Primary Cell Culture Approach. J. Biol. Regul. Homeost. Agents 2016, 30, 693–702. [Google Scholar] [PubMed]
- Kossowska-Tomaszczuk, K.; De Geyter, C.; De Geyter, M.; Martin, I.; Holzgreve, W.; Scherberich, A.; Zhang, H. The Multipotency of Luteinizing Granulosa Cells Collected from Mature Ovarian Follicles. Stem. Cells 2009, 27, 210–219. [Google Scholar] [CrossRef] [PubMed]
- Borys, S.; Khozmi, R.; Kranc, W.; Bryja, A.; Jeseta, M.; Kempisty, B. Resveratrol and Its Analogues—Is It a New Strategy of Anticancer Therapy? Adv. Cell Biol. 2017, 5, 32–42. [Google Scholar] [CrossRef]
- Alpy, F.; Tomasetto, C. Give Lipids a START: The StAR-Related Lipid Transfer (START) Domain in Mammals. J. Cell Sci. 2005, 118, 2791–2801. [Google Scholar] [CrossRef] [PubMed]
- Stocco, D.M. Steroidogenic Acute Regulatory Protein. Vitam. Horm. 1998, 55, 399–441. [Google Scholar]
- Clark, B.J.; Soo, S.C.; Caron, K.M.; Ikeda, Y.; Parker, K.L.; Stocco, D.M. Hormonal and Developmental Regulation of the Steroidogenic Acute Regulatory Protein. Mol. Endocrinol. 1995, 9, 1346–1355. [Google Scholar] [PubMed]
- Caron, K.M.; Soo, S.C.; Wetsel, W.C.; Stocco, D.M.; Clark, B.J.; Parker, K.L. Targeted Disruption of the Mouse Gene Encoding Steroidogenic Acute Regulatory Protein Provides Insights into Congenital Lipoid Adrenal Hyperplasia. Proc. Natl. Acad. Sci. USA 1997, 94, 11540–11545. [Google Scholar] [CrossRef] [PubMed]
- Bose, H.S.; Sugawara, T.; Strauss, J.F.; Miller, W.L. The Pathophysiology and Genetics of Congenital Lipoid Adrenal Hyperplasia. N. Engl. J. Med. 1996, 335, 1870–1879. [Google Scholar] [CrossRef] [PubMed]
- Soccio, R.E.; Adams, R.M.; Romanowski, M.J.; Sehayek, E.; Burley, S.K.; Breslow, J.L. The Cholesterol-Regulated StarD4 Gene Encodes a StAR-Related Lipid Transfer Protein with Two Closely Related Homologues, StarD5 and StarD6. Proc. Natl. Acad. Sci. USA 2002, 99, 6943–6948. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Agudo, D.; Calderon-Dominguez, M.; Ren, S.; Marques, D.; Redford, K.; Medina-Torres, M.A.; Hylemon, P.; Gil, G.; Pandak, W.M. Subcellular Localization and Regulation of StarD4 Protein in Macrophages and Fibroblasts. Biochim. Biophys. Acta 2011, 1811, 597–606. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Sato, R.; Brown, M.S.; Hua, X.; Goldstein, J.L. SREBP-1, a Membrane-Bound Transcription Factor Released by Sterol-Regulated Proteolysis. Cell 1994, 77, 53–62. [Google Scholar] [CrossRef]
- Brown, M.S.; Goldstein, J.L. The SREBP Pathway: Regulation of Cholesterol Metabolism by Proteolysis of a Membrane-Bound Transcription Factor. Cell 1997, 89, 331–340. [Google Scholar] [CrossRef]
- Shimano, H.; Yahagi, N.; Amemiya-Kudo, M.; Hasty, A.H.; Osuga, J.; Tamura, Y.; Shionoiri, F.; Iizuka, Y.; Ohashi, K.; Harada, K.; et al. Sterol Regulatory Element-Binding Protein-1 as a Key Transcription Factor for Nutritional Induction of Lipogenic Enzyme Genes. J. Biol. Chem. 1999, 274, 35832–35839. [Google Scholar] [CrossRef] [PubMed]
- Edwards, P.; Tabor, D.; Kast, H.R.; Venkateswaran, A. Regulation of Gene Expression by SREBP and SCAP. Biochim. Biophys. Acta 2000, 1529, 103–113. [Google Scholar] [CrossRef]
- Nohturfft, A.; DeBose-Boyd, R.A.; Scheek, S.; Goldstein, J.L.; Brown, M.S. Sterols Regulate Cycling of SREBP Cleavage-Activating Protein (SCAP) between Endoplasmic Reticulum and Golgi. Proc. Natl. Acad. Sci. USA 1999, 96, 11235–11240. [Google Scholar] [CrossRef] [PubMed]
- Shao, W.; Espenshade, P.J. Sterol Regulatory Element-Binding Protein (SREBP) Cleavage Regulates Golgi-to-Endoplasmic Reticulum Recycling of SREBP Cleavage-Activating Protein (SCAP). J. Biol. Chem. 2014, 289, 7547–7557. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Espenshade, P.J.; Wright, M.E.; Yabe, D.; Gong, Y.; Aebersold, R.; Goldstein, J.L.; Brown, M.S. Crucial Step in Cholesterol Homeostasis: Sterols Promote Binding of SCAP to INSIG-1, a Membrane Protein That Facilitates Retention of SREBPs in ER. Cell 2002, 110, 489–500. [Google Scholar] [CrossRef]
- Huang, N.; Pandey, A.V.; Agrawal, V.; Reardon, W.; Lapunzina, P.D.; Mowat, D.; Jabs, E.W.; Vliet, G.; Van Sack, J.; Flück, C.E.; et al. Diversity and Function of Mutations in P450 Oxidoreductase in Patients with Antley-Bixler Syndrome and Disordered Steroidogenesis. Am. J. Hum. Genet. 2005, 76, 729–749. [Google Scholar] [CrossRef] [PubMed]
- Hart, S.N.; Zhong, X. P450 Oxidoreductase: Genetic Polymorphisms and Implications for Drug Metabolism and Toxicity. Expert Opin. Drug Metab. Toxicol. 2008, 4, 439–452. [Google Scholar] [CrossRef] [PubMed]
- Arlt, W.; Walker, E.A.; Draper, N.; Ivison, H.E.; Ride, J.P.; Hammer, F.; Chalder, S.M.; Borucka-Mankiewicz, M.; Hauffa, B.P.; Malunowicz, E.M.; et al. Congenital Adrenal Hyperplasia Caused by Mutant P450 Oxidoreductase and Human Androgen Synthesis: Analytical Study. Lancet 2004, 363, 2128–2135. [Google Scholar] [CrossRef]
- Flück, C.E.; Tajima, T.; Pandey, A.V.; Arlt, W.; Okuhara, K.; Verge, C.F.; Jabs, E.W.; Mendonça, B.B.; Fujieda, K.; Miller, W.L. Mutant P450 Oxidoreductase Causes Disordered Steroidogenesis with and without Antley-Bixler Syndrome. Nat. Genet. 2004, 36, 228–230. [Google Scholar]
- Waterham, H.; Wanders, R.J. Biochemical and Genetic Aspects of 7-Dehydrocholesterol Reductase and Smith-Lemli-Opitz Syndrome. Biochim. Biophys. Acta 2000, 1529, 340–356. [Google Scholar] [CrossRef]
- Witsch-Baumgartner, M.; Löffler, J.; Utermann, G. Mutations in the Human DHCR7 Gene. Hum. Mutat. 2001, 17, 172–182. [Google Scholar] [CrossRef] [PubMed]
- Saeed, M.; Andreo, U.; Chung, H.-Y.; Espiritu, C.; Branch, A.D.; Silva, J.M.; Rice, C.M. SEC14L2 Enables Pan-Genotype HCV Replication in Cell Culture. Nature 2015, 524, 471–475. [Google Scholar] [CrossRef] [PubMed]
- Zingg, J.-M.; Libinaki, R.; Meydani, M.; Azzi, A. Modulation of Phosphorylation of Tocopherol and Phosphatidylinositol by hTAP1/SEC14L2-Mediated Lipid Exchange. PLoS ONE 2014, 9, e101550. [Google Scholar] [CrossRef] [PubMed]
- Fechtner, S.; Singh, A.; Chourasia, M.; Ahmed, S. Molecular Insights into the Differences in Anti-Inflammatory Activities of Green Tea Catechins on IL-1β Signaling in Rheumatoid Arthritis Synovial Fibroblasts. Toxicol. Appl. Pharmacol. 2017, 329, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Miao, E.A.; Alpuche-Aranda, C.M.; Dors, M.; Clark, A.E.; Bader, M.W.; Miller, S.I.; Aderem, A. Cytoplasmic Flagellin Activates Caspase-1 and Secretion of Interleukin 1[beta] via Ipaf. Nat. Immunol. 2006, 7, 569–576. [Google Scholar] [CrossRef] [PubMed]
- Ben-Sasson, S.Z.; Hu-Li, J.; Quiel, J.; Cauchetaux, S.; Ratner, M.; Shapira, I.; Dinarello, C.A.; Paul, W.E. IL-1 Acts Directly on CD4 T Cells to Enhance Their Antigen-Driven Expansion and Differentiation. Proc. Natl. Acad. Sci. USA 2009, 106, 7119–7124. [Google Scholar] [CrossRef] [PubMed]
- Bartelmez, S.H.; Bradley, T.R.; Bertoncello, I.; Mochizuki, D.Y.; Tushinski, R.J.; Stanley, E.R.; Hapel, A.J.; Young, I.G.; Kriegler, A.B.; Hodgson, G.S. Interleukin 1 plus Interleukin 3 plus Colony-Stimulating Factor 1 Are Essential for Clonal Proliferation of Primitive Myeloid Bone Marrow Cells. Exp. Hematol. 1989, 17, 240–245. [Google Scholar] [PubMed]
- Mangan, D.F.; Welch, G.R.; Wahl, S.M. Lipopolysaccharide, Tumor Necrosis Factor-Alpha, and IL-1 Beta Prevent Programmed Cell Death (Apoptosis) in Human Peripheral Blood Monocytes. J. Immunol. 1991, 146, 1541–1546. [Google Scholar] [PubMed]
- Zhu, J.; Guo, L.; Min, B.; Watson, C.J.; Hu-Li, J.; Young, H.A.; Tsichlis, P.N.; Paul, W.E. Growth Factor Independent-1 Induced by IL-4 Regulates Th2 Cell Proliferation. Immunity 2002, 16, 733–744. [Google Scholar] [CrossRef]
- Hock, H.; Hamblen, M.J.; Rooke, H.M.; Schindler, J.W.; Saleque, S.; Fujiwara, Y.; Orkin, S.H. Gfi-1 Restricts Proliferation and Preserves Functional Integrity of Haematopoietic Stem Cells. Nature 2004, 431, 1002–1007. [Google Scholar] [CrossRef] [PubMed]
- Person, R.E.; Li, F.-Q.; Duan, Z.; Benson, K.F.; Wechsler, J.; Papadaki, H.A.; Eliopoulos, G.; Kaufman, C.; Bertolone, S.J.; Nakamoto, B.; et al. Mutations in Proto-Oncogene GFI1 Cause Human Neutropenia and Target ELA2. Nat. Genet. 2003, 34, 308–312. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Stravopodis, D.; Teglund, S.; Kitazawa, J.; Ihle, J.N. Naturally Occurring Dominant Negative Variants of Stat5. Mol. Cell. Biol. 1996, 16, 6141–6148. [Google Scholar] [CrossRef] [PubMed]
- Ambrosioa, R.; Fimiania, G.; Monfregolaa, J.; Sanzaria, E.; Felicea, N.; De Salernob, M.C.; Pignatab, C.; D’Ursoa, M.; Valeria Ursini, M. The Structure of Human STAT5A and B Genes Reveals Two Regions of Nearly Identical Sequence and an Alternative Tissue Specific STAT5B Promoter. Gene 2002, 285, 311–318. [Google Scholar] [CrossRef]
- Lin, J.X.; Mietz, J.; Modi, W.S.; John, S.; Leonard, W.J. Cloning of Human Stat5B. Reconstitution of Interleukin-2-Induced Stat5A and Stat5B DNA Binding Activity in COS-7 Cells. J. Biol. Chem. 1996, 271, 10738–10744. [Google Scholar] [CrossRef] [PubMed]
- Miyoshi, K.; Shillingford, J.M.; Smith, G.H.; Grimm, S.L.; Wagner, K.U.; Oka, T.; Rosen, J.M.; Robinson, G.W.; Hennighausen, L. Signal Transducer and Activator of Transcription (Stat) 5 Controls the Proliferation and Differentiation of Mammary Alveolar Epithelium. J. Cell Biol. 2001, 155, 531–542. [Google Scholar] [CrossRef] [PubMed]
- Chomczynski, P.; Sacchi, N. Single-Step Method of RNA Isolation by Acid Guanidinium Thiocyanate-Phenol-Chloroform Extraction. Anal. Biochem. 1987, 162, 156–159. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; Tan, Q.; Kir, J.; Liu, D.; Bryant, D.; Guo, Y.; Stephens, R.; Baseler, M.W.; Lane, H.C.; et al. DAVID Bioinformatics Resources: Expanded Annotation Database and Novel Algorithms to Better Extract Biology from Large Gene Lists. Nucleic Acids Res. 2007, 35, W169–W175. [Google Scholar] [CrossRef] [PubMed]
- Walter, W.; Sánchez-Cabo, F.; Ricote, M. GOplot: An R Package for Visually Combining Expression Data with Functional Analysis: Figure 1. Bioinformatics 2015, 31, 2912–2914. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Official Gene Symbol | Fold Change D1/D7 | Fold Change D1/D15 | Fold Change D1/D30 | adj.P.Val. D1/D7 | adj.P.Val. D1/D15 | adj.P.Val. D1/D30 | Entrez Gene ID |
|---|---|---|---|---|---|---|---|
| DHCR7 | 0.174701154 | 0.135435372 | 0.287884941 | 0.039383847 | 0.023551624 | 0.084599974 | 1717 |
| GFI1 | 0.310857826 | 0.318291331 | 0.286701236 | 0.001572194 | 0.001472925 | 0.000931728 | 2672 |
| IL1B | 0.068991209 | 0.095213232 | 0.183379317 | 0.027149434 | 0.035799475 | 0.083315453 | 3553 |
| INSIG1 | 0.112401766 | 0.111379044 | 0.231549573 | 0.040421878 | 0.035709593 | 0.102319923 | 3638 |
| POR | 0.297205384 | 0.246609697 | 0.226675727 | 0.02007617 | 0.011465695 | 0.008491922 | 5447 |
| SCAP | 0.428465981 | 0.37245418 | 0.350801297 | 0.00234869 | 0.001405483 | 0.000934051 | 22937 |
| SEC14L2 | 0.393655866 | 0.47190842 | 0.474492018 | 0.049169113 | 0.080230709 | 0.075670468 | 23541 |
| SREBF1 | 0.342373023 | 0.40104925 | 0.351769946 | 0.004735431 | 0.007059315 | 0.004002643 | 6720 |
| STAR | 0.020791115 | 0.015709262 | 0.021002203 | 0.000945846 | 0.000687578 | 0.000705428 | 6770 |
| STARD4 | 0.192889636 | 0.155104665 | 0.215393387 | 0.037708661 | 0.023368014 | 0.037965777 | 134429 |
| STAT5B | 0.343238914 | 0.381840346 | 0.358794257 | 0.008451029 | 0.010289799 | 0.007445427 | 6777 |
| Gene | Gene Accession Number | Primer Sequence (5′-3′) | Product Size (bp) |
|---|---|---|---|
| STAR | NM_000349.2 | GGCATCCTTAGCAACCAAGA TCTCCTTGACATTGGGGTTC | 199 |
| Il1B | NM_000576.2 | GGGCCTCAAGGAAAAGAATC TTCTGCTTGAGAGGTGCTGA | 205 |
| POR | NM_000941.2 | CACAAGGTCTACGTCCAGCA GCCACGATGTCGTAGAAGGT | 143 |
| GAPDH | NM_002046 | TCAGCCGCATCTTCTTTTGC ACGACCAAATCCGTTGACTC | 90 |
| ACTB | NM_001101 | AAAGACCTGTACGCCAACAC CTCAGGAGGAGCAATGATCTTG | 132 |
| HPRT | NM_000194 | TGGCGTCGTGATTAGTGATG ACATCTCGAGCAAGACGTTC | 141 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kranc, W.; Brązert, M.; Ożegowska, K.; Nawrocki, M.J.; Budna, J.; Celichowski, P.; Dyszkiewicz-Konwińska, M.; Jankowski, M.; Jeseta, M.; Pawelczyk, L.; et al. Expression Profile of Genes Regulating Steroid Biosynthesis and Metabolism in Human Ovarian Granulosa Cells—A Primary Culture Approach. Int. J. Mol. Sci. 2017, 18, 2673. https://doi.org/10.3390/ijms18122673
Kranc W, Brązert M, Ożegowska K, Nawrocki MJ, Budna J, Celichowski P, Dyszkiewicz-Konwińska M, Jankowski M, Jeseta M, Pawelczyk L, et al. Expression Profile of Genes Regulating Steroid Biosynthesis and Metabolism in Human Ovarian Granulosa Cells—A Primary Culture Approach. International Journal of Molecular Sciences. 2017; 18(12):2673. https://doi.org/10.3390/ijms18122673
Chicago/Turabian StyleKranc, Wiesława, Maciej Brązert, Katarzyna Ożegowska, Mariusz J. Nawrocki, Joanna Budna, Piotr Celichowski, Marta Dyszkiewicz-Konwińska, Maurycy Jankowski, Michal Jeseta, Leszek Pawelczyk, and et al. 2017. "Expression Profile of Genes Regulating Steroid Biosynthesis and Metabolism in Human Ovarian Granulosa Cells—A Primary Culture Approach" International Journal of Molecular Sciences 18, no. 12: 2673. https://doi.org/10.3390/ijms18122673