
The Complete Structure of the Core Oligosaccharide from Edwardsiella tarda EIB 202 Lipopolysaccharide


 ,
,
Abstract
:
1. Introduction
2. Results
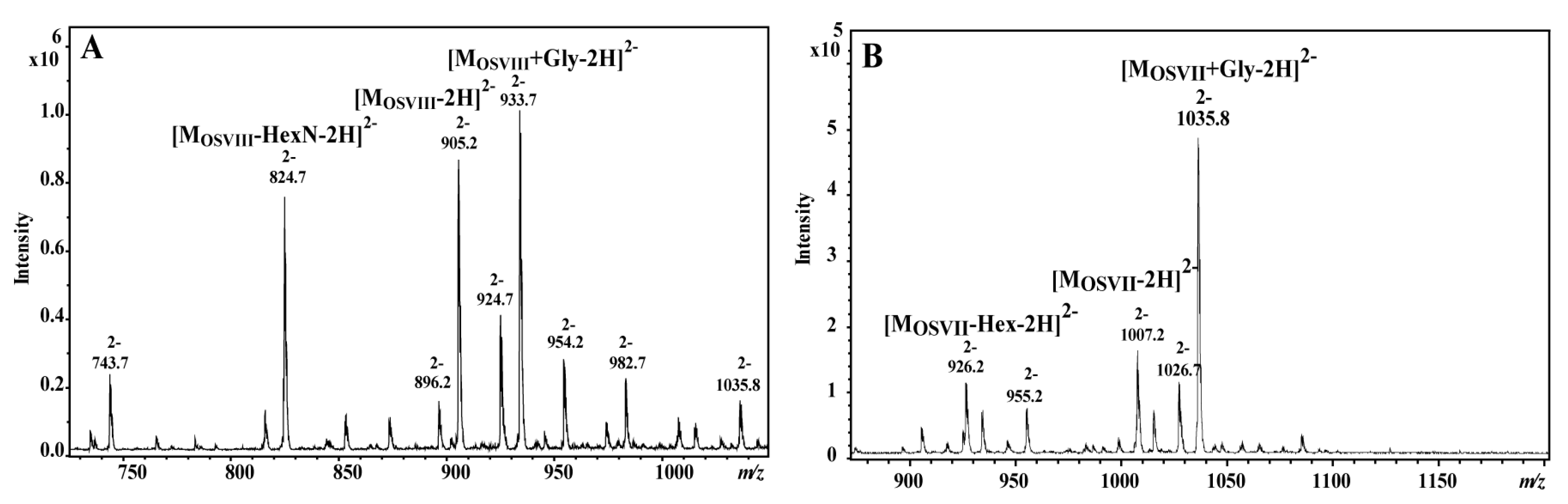
2.1. Isolation of the Core Oligosaccharide
2.2. Structure Analysis of the Core Oligosaccharide Fractions
2.3. Organization of the E. tarda Strain EIB 202 waa Gene Cluster
3. Discussion
4. Materials and Methods
4.1. Growth Conditions and Isolation of the Lipopolysaccharide and the Polysaccharide
4.2. Chemical Methods
4.3. Instrumental Methods
4.4. Comparative Genomics
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Matsuyama, T.; Kamaishi, T.; Ooseko, N.; Kurohara, K.; Iida, T. Pathogenicity of motile and non-motile Edwardsiella tarda to some marine fish. Fish Pathol. 2005, 40, 133–135. [Google Scholar] [CrossRef]
- Mohanty, B.R.; Sahoo, P.K. Edwardsiellosis in fish: A brief review. J. Biosci. 2007, 32, 1331–1344. [Google Scholar] [CrossRef] [PubMed]
- Janda, J.M.; Abbott, S.L. Infections associated with the genus Edwardsiella: The role of Edwardsiella tarda in human disease. Clin. Infect. Dis. 1993, 17, 742–748. [Google Scholar] [PubMed]
- Nelson, J.J.; Nelson, C.A.; Carter, J.E. Extraintestinal manifestations of Edwardsiella tarda infection: A 10-year retrospective review. J. La. State Med. Soc. 2009, 161, 103–106. [Google Scholar] [PubMed]
- Leung, K.Y.; Siame, B.A.; Tenkink, B.J.; Noort, R.J.; Mok, Y.K. Edwardsiella tarda virulence mechanisms of an emerging gastroenteritis pathogen. Microbes Infect. 2012, 14, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Jin, R.P.; Hu, Y.H.; Sun, B.G.; Zhang, X.H.; Sun, L. Edwardsiella tarda sialidase: Pathogenicity involvement and vaccine potential. Fish Shellfish Immunol. 2012, 33, 514–521. [Google Scholar] [CrossRef] [PubMed]
- Aquilini, E.; Tomás, J.M. (Eds.) Lipopolysaccharides (Endotoxins); Reference Module in Biomedical Sciences; Elsevier: Amsterdam, The Netherlands, 2015. [Google Scholar] [CrossRef]
- Katzenellenbogen, E.; Kocharova, N.A.; Shashkov, A.S.; Gorska-Fraczek, S.; Gamian, A.; Knirel, Y.A. Structure of the O-polysaccharide of Edwardsiella tarda PCM 1156. Carbohydr. Res. 2013, 477, 45–48. [Google Scholar] [CrossRef] [PubMed]
- Katzenellenbogen, E.; Kocharova, N.A.; Shashkov, A.S.; Gorska-Fraczek, S.; Bogulska, M.; Gamian, A.; Knirel, Y.A. Structure of the O-polysaccharide of Edwardsiella tarda PCM 1150 containing an amide of d-glucuronic acid with l-alanine. Carbohydr. Res. 2013, 347, 84–88. [Google Scholar] [CrossRef] [PubMed]
- Katzenellenbogen, E.; Kocharova, N.A.; Toukach, P.V.; Gorska, S.; Bogulska, M.; Gamian, A.; Knirel, Y.A. Structures of a unique O-polysaccharide of Edwardsiella tarda PCM 1153 containing an amide of galacturonic acid with 2-aminopropane-1,3-diol and an abequose-containing O-polysaccharide shared by E. tarda PCM 1145, PCM 1151 and PCM 1158. Carbohydr. Res. 2012, 355, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Vinogradov, E.; Nossova, L.; Perry, M.B.; Kay, W.W. Structural characterization of the O-polysaccharide antigen of Edwardsiella tarda MT 108. Carbohydr. Res. 2005, 340, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Heinrichs, D.E.; Yethon, J.A.; Whitfield, C. Molecular basis for structural diversity in the core regions of the lipopolysaccharides of Escherichia coli and Salmonella enterica. Mol. Microbiol. 1998, 30, 221–232. [Google Scholar] [CrossRef] [PubMed]
- Regué, M.; Climent, N.; Abitiu, N.; Coderch, N.; Merino, S.; Izquierdo, L.; Altarriba, M.; Tomás, J.M. Genetic characterization of the Klebsiella pneumoniae waa gene cluster, involved in core lipopolysaccharide biosynthesis. J. Bacteriol. 2001, 183, 3564–3573. [Google Scholar] [CrossRef] [PubMed]
- Knirel, Y.A.; Dentovskaya, S.V.; Bystrova, O.V.; Kocharova, N.A.; Senchenkova, S.N.; Shaikhutdinova, R.Z.; Titareva, G.M.; Bakhteeva, I.V.; Lindner, B.; Pier, G.B.; et al. Relationship of the lipopolysaccharide structure of Yersinia pestis to resistance to antimicrobial factors. Adv. Exp. Med. Biol. 2007, 603, 88–96. [Google Scholar] [PubMed]
- Aquilini, E.; Azevedo, J.; Jimenez, N.; Bouamama, L.; Tomás, J.M.; Regué, M. Functional Identification of the Proteus mirabilis core lipopolysaccharide biosynthetic genes. J. Bacteriol. 2010, 192, 4413–4424. [Google Scholar] [CrossRef] [PubMed]
- Aquilini, E.; Merino, S.; Regué, M.; Tomás, J.M. Genomic and proteomic studies of Plesiomonas shigelloides lipopolysaccharide core biosynthesis. J. Bacteriol. 2014, 196, 556–567. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Wang, Q.; Xiao, J.; Liu, Q.; Wang, X.; Chen, T.; Zhang, Y. Characterization of Edwardsiella tarda waaL: Roles in lipopolysaccharide biosynthesis, stress adaptation, and virulence toward fish. Arch. Microbiol. 2010, 192, 1039–1047. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.F.; Wang, Q.Y.; Liu, Q.; Wang, X.; Liu, H.; Zhang, Y.X. Isolation and identification of fish pathogen Edwardsiella tarda from mariculture in China. Aquac. Res. 2008, 40, 13–17. [Google Scholar] [CrossRef]
- Wang, Q.; Yang, M.; Xiao, J.; Wu, H.; Wang, X.; Lu, Y.; Xu, L.; Zheng, H.; Wang, S.; Zhao, G.; et al. Genome sequence of the versatile fish pathogen Edwardsiella tarda provides insights into its adaptation to broad host ranges and intracellular niches. PLoS ONE 2009, 4, e7646. [Google Scholar] [CrossRef] [PubMed]
- Geerlof, A.; Lexendon, A.; Shaw, V. Purification and characterization of phosphopantheine adenyltransferase from Escherichia coli. J. Biol. Chem. 1999, 274, 27105–27111. [Google Scholar] [CrossRef] [PubMed]
- Kaszowska, M.; Jachymek, W.; Jachymek, W.; Lukasiewicz, J.; Niedziala, T.; Kenne, L.; Lugowski, C. The unique structure of complete lipopolysaccharide isolated from semi-rough Plesiomonas shigelloides O37 (strain CNCTC 39/89) containing (2S)-O-(4-oxopentanoic acid)-α-d-Glcp (α-d-lenose). Carbohydr. Res. 2013, 378, 98–107. [Google Scholar] [CrossRef] [PubMed]
- Westphal, O.; Jann, K. Bacterial lipopolysaccharide: Extraction with phenol-water and further application of the procedure. Methods Carbohydr. Chem. 1965, 5, 83–89. [Google Scholar]
- Ciucanu, I.; Kerek, F. A simple and rapid method for the permethylation of carbohydrates. Carbohydr. Res. 1984, 131, 209–217. [Google Scholar] [CrossRef]
- Gerwig, G.J.; Kamerling, J.P.; Vliegenthart, J.F.G. Determination of the D and L configuration of neutral monosaccharides by high-resolution capillary g.l.c. Carbohydr. Res. 1978, 62, 349–357. [Google Scholar] [CrossRef]
- Goddard, T.D.; Kneller, D.G. SPARKY 3; University of California: San Francisco, CA, USA, 2001. Available online: https://www.cgl.ucsf.edu/home/sparky/ (accessed on 30 May 2017).




{kind=link}
{kind=link}
{kind=link}
| Residues | Oligosaccharide | Chemical Shifts (ppm) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| OSVIII | OSVII | H1/C1 | H2/C2 | H3(ax,eq)/C3 | H4/C4 | H5/C5 | H6,6`/C6 | H7,7`/C7 | H8,8`/C8 (NAc) | |
| A →5)-Kdo | * | * | - | - | 1.96, 2.29 | 4.21 | 4.25 | 3.81 | 3.97 | 3.69, 3.92 |
| nd | 97.7 | 34.7 | 66.5 | 75.7 | 70.1 | 66.9 | 64.3 | |||
| B →3,4)-l-glycero-α-d-manno-Hepp-(1→ | * | * | 5.19 | 4.15 | 4.25 | 4.35 | 4.26 | 4.18 | 3.80 | |
| 101.7 | 71.3 | 74.7 | 74.4 | 72.2 | 69.4 | 63.9 | ||||
| C →β-d-Glcp-(1→ | * | * | 4.51 | 3.36 | 3.55 | 3.56 | 3.55 | 3.87, 3.95 | ||
| 103.3 | 73.9 | 76.2 | 69.9 | 76.2 | 61.5 | |||||
| D →2,3,7)-l-glycero-α-d-manno-Hepp-(1→ | * | * | 5.44 | 4.34 | 4.19 | 4.12 | 3.67 | 4.28 | 3.62, 4.01 | |
| 99.6 | 78.6 | 80.0 | 66.6 | 73.4 | 69.3 | 73.3 | ||||
| E →7)-l-glycero-α-d-manno-Hepp-(1→ | * | * | 4.98 | 4.05 | 3.95 | 3.93 | 3.72 | 4.23 | 3.69, 4.88 | |
| 103.2 | 71.1 | 72.3 | 71.4 | 73.4 | 69.1 | 72.0 | ||||
| F →4)-α-d-GalpA-(1→ | * | * | 5.12 | 4.05 | 4.16 | 4.62 | 4.41 | |||
| 99.8 | 69.0 | 69.9 | 77.9 | 70.9 | 176.7 | |||||
| G α-d-GlcpN-(1→ | * | * | 5.33 | 3.34 | 4.02 | 3.60 | 4.19 | 3.91a | ||
| 95.6 | 55.1 | 70.4 | 76.6 | 73.2 | 60.0 | |||||
| H α-d-Glcp-(1→ | * | * | 4.63 | 3.40 | 3.56 | 3.33 | 3.60 | 3.69, 3.98 | ||
| 103.2 | 74.5 | 76.3 | 71.6 | 76.5 | 62.7 | |||||
| I →4)-α-d-GalpA-(1→ | * | * | 5.47 | 3.93 | 4.26 | 4.48 | 4.67 | - | ||
| 102.5 | 69.6 | 72.2 | 80.9 | 72.5 | 175.6 | |||||
| J α-d-GlcpN-(1→ | * | 5.29 | 3.39 | 3.99 | 3.63 | 4.27 | 3.69, 3.97 | |||
| 97.0 | 55.1 | 70.5 | 70.5 | 72.8 | 62.5 | |||||
| K α-d-GlcpNAc-(1→ | * | 5.06 | 4.02 | 3.83 | 3.63 | 4.28 | 3.90a | 2.13 | ||
| 99.8 | 54.6 | 71.6 | 70.2 | 73.2 | 61.0 | 23.2 | ||||
| L Gly | * | * | 3.90, 4.09 | |||||||
| 168.0 | 41.8 | |||||||||
| M →β-d-Galp-(1→ | * | 4.45 | 3.61 | 3.73 | 3.93 | 3.59 | 3.74, 3.80 | |||
| 103.6 | 71.0 | 72.7 | 71.4 | 75.1 | 63.2 | |||||
| Protein | Homologus Protein | Predicted Function | % Identity/Similarity |
|---|---|---|---|
| ETAE_0083 | HldE Enterobacteriaceae | ADP-l-glycero-d-manno Heptose-6-epimerase | 85/91 |
| ETAE_0082 | WaaF Enterobacteriaceae | ADP-heptose:LPS heptosyl transferase II | 76/86 |
| ETAE_0081 | WaaC Enterobacteriaceae | ADP-heptose:LPS heptosyl transferase I | 78/84 |
| ETAE_0080 | WaaL Klebsiella pneumoniae | O-antigen ligase | 29/46 |
| ETAE_0079 | Similar only to WabK Klebsiella pneumoniae | unknown | 34/53 |
| ETAE_0078 | WapC Plesiomonas shigelloides | UDP-galacturonic a transferase α(1→7) to HepIII acid | 76/89 |
| ETAE_0077 | WapB Plesiomonas shigelloides | UDP-N-acetyl glucosamine α(1→4) to GalAII | 61/82 |
| ETAE_0076 | WabN Klebsiella pneumoniae | Protein deacetilase | 77/89 |
| ETAE_0075 | WaaQ Klebsiella pneumoniae | ADP-heptose: LPS heptosyl | 72/83 |
| ETAE_0074 | WabG Klebsiella pneumoniae | UDP-galacturonic acid transferase α(1→3) to HepIII | 78/87 |
| ETAE_0073 | WabH Klebsiella pneumoniae | UDP-N-acetyl glucosamine transferase α(1→4) to GalAI | 70/83 |
| ETAE_0072 | WaaA Enterobacteriaceae | 3-deoxy-d-manno-octulosonic acid transferase | 88/96 |
| ETAE_0071 | WapE Plesiomonas shigelloides | UDP-galactose transferase β(1→4) to l-HepI | 82/97 |
| ETAE_0070 | CoaD Enterobacteriaceae | Phosphopantetehine adenylyltransferase | 81/89 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaszowska, M.; De Mendoza-Barberá, E.; Maciejewska, A.; Merino, S.; Lugowski, C.; Tomás, J.M. The Complete Structure of the Core Oligosaccharide from Edwardsiella tarda EIB 202 Lipopolysaccharide. Int. J. Mol. Sci. 2017, 18, 1163. https://doi.org/10.3390/ijms18061163
Kaszowska M, De Mendoza-Barberá E, Maciejewska A, Merino S, Lugowski C, Tomás JM. The Complete Structure of the Core Oligosaccharide from Edwardsiella tarda EIB 202 Lipopolysaccharide. International Journal of Molecular Sciences. 2017; 18(6):1163. https://doi.org/10.3390/ijms18061163
Chicago/Turabian StyleKaszowska, Marta, Elena De Mendoza-Barberá, Anna Maciejewska, Susana Merino, Czeslaw Lugowski, and Juan M. Tomás. 2017. "The Complete Structure of the Core Oligosaccharide from Edwardsiella tarda EIB 202 Lipopolysaccharide" International Journal of Molecular Sciences 18, no. 6: 1163. https://doi.org/10.3390/ijms18061163