Comparative Proteomic Profiling Reveals Molecular Characteristics Associated with Oogenesis and Oocyte Maturation during Ovarian Development of Bactrocera dorsalis (Hendel)

 , and
, and
Abstract
:
1. Introduction
2. Results and Discussion
2.1. Proteome-Wide Identification of Proteins in Ovaries
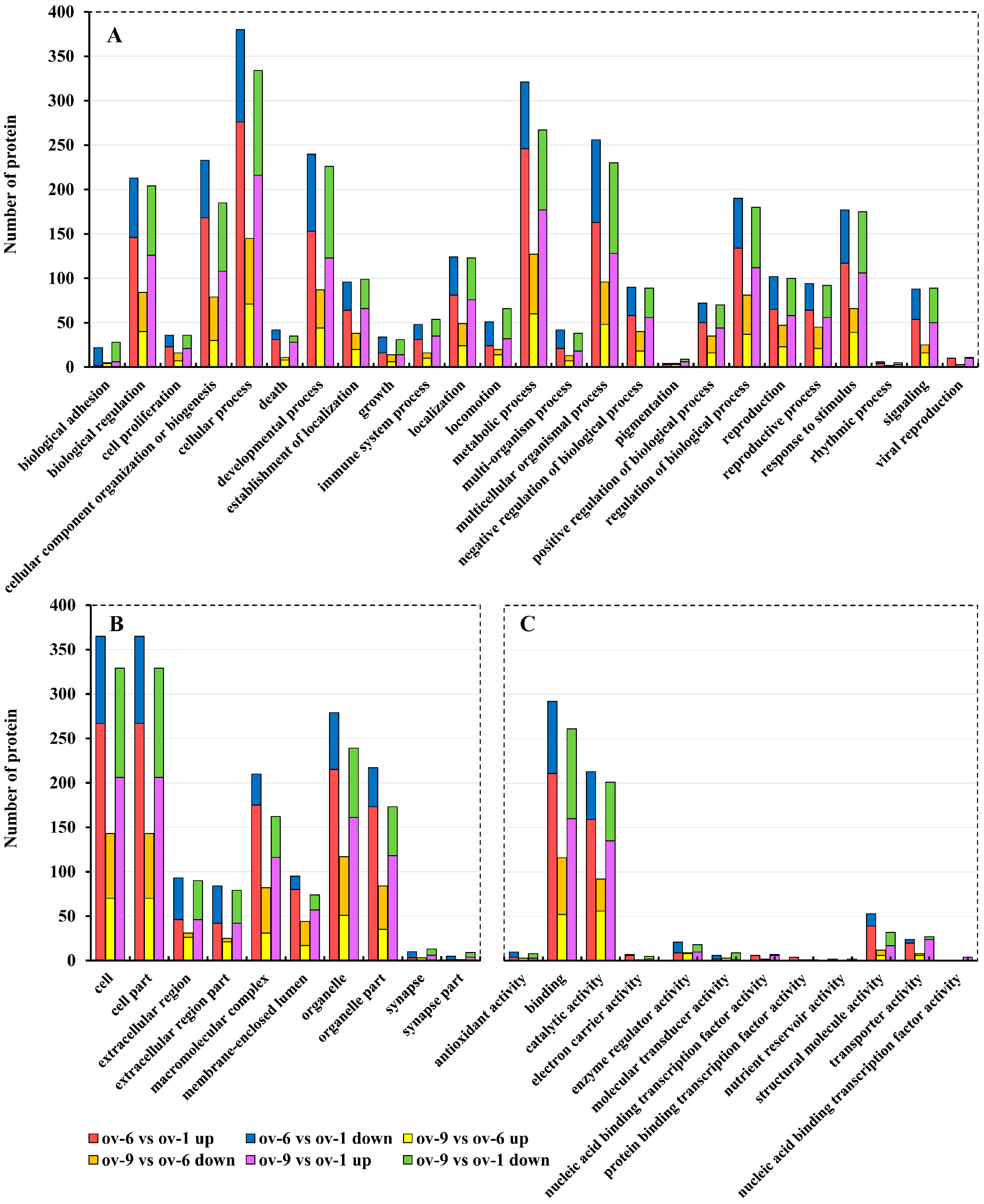
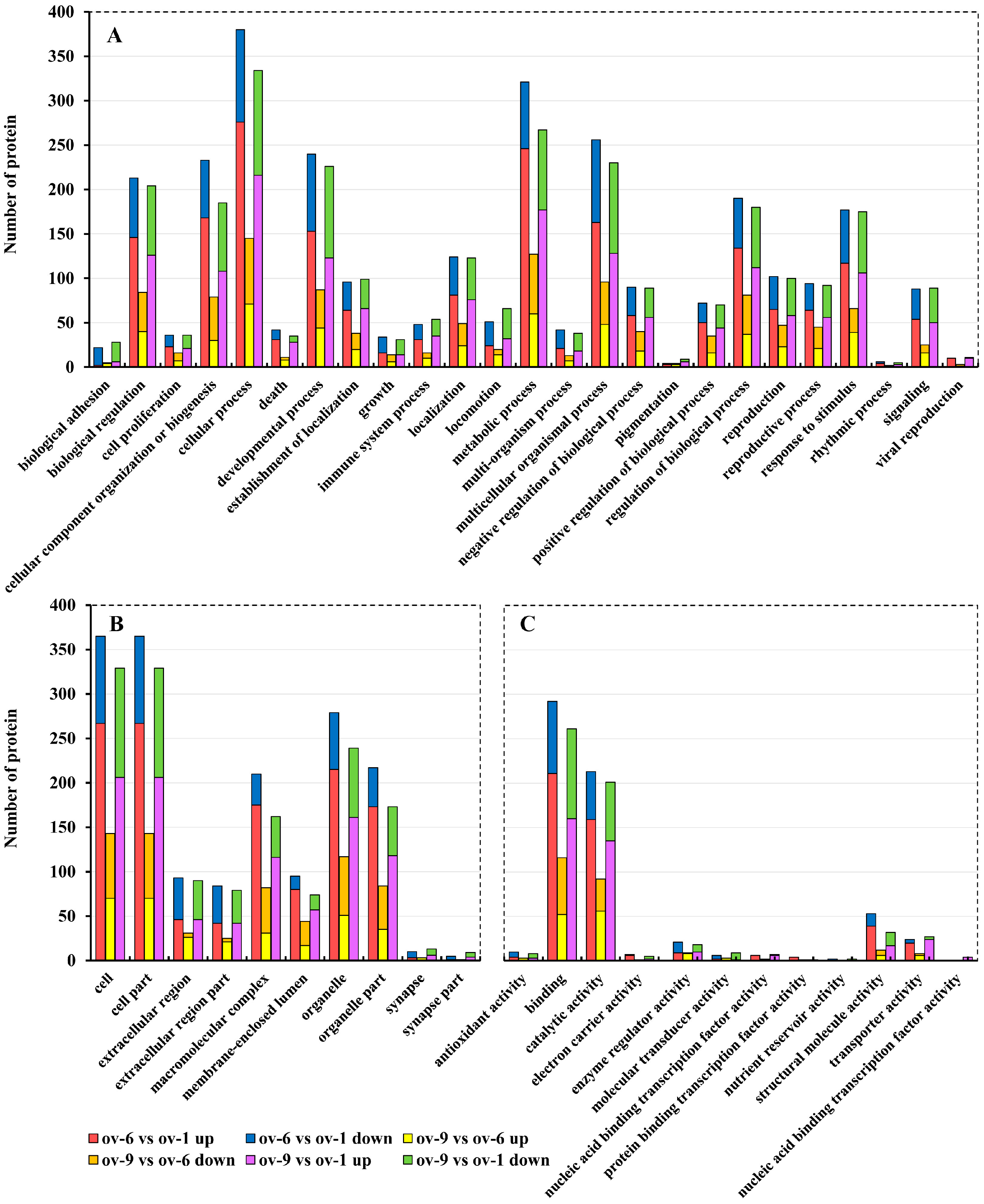
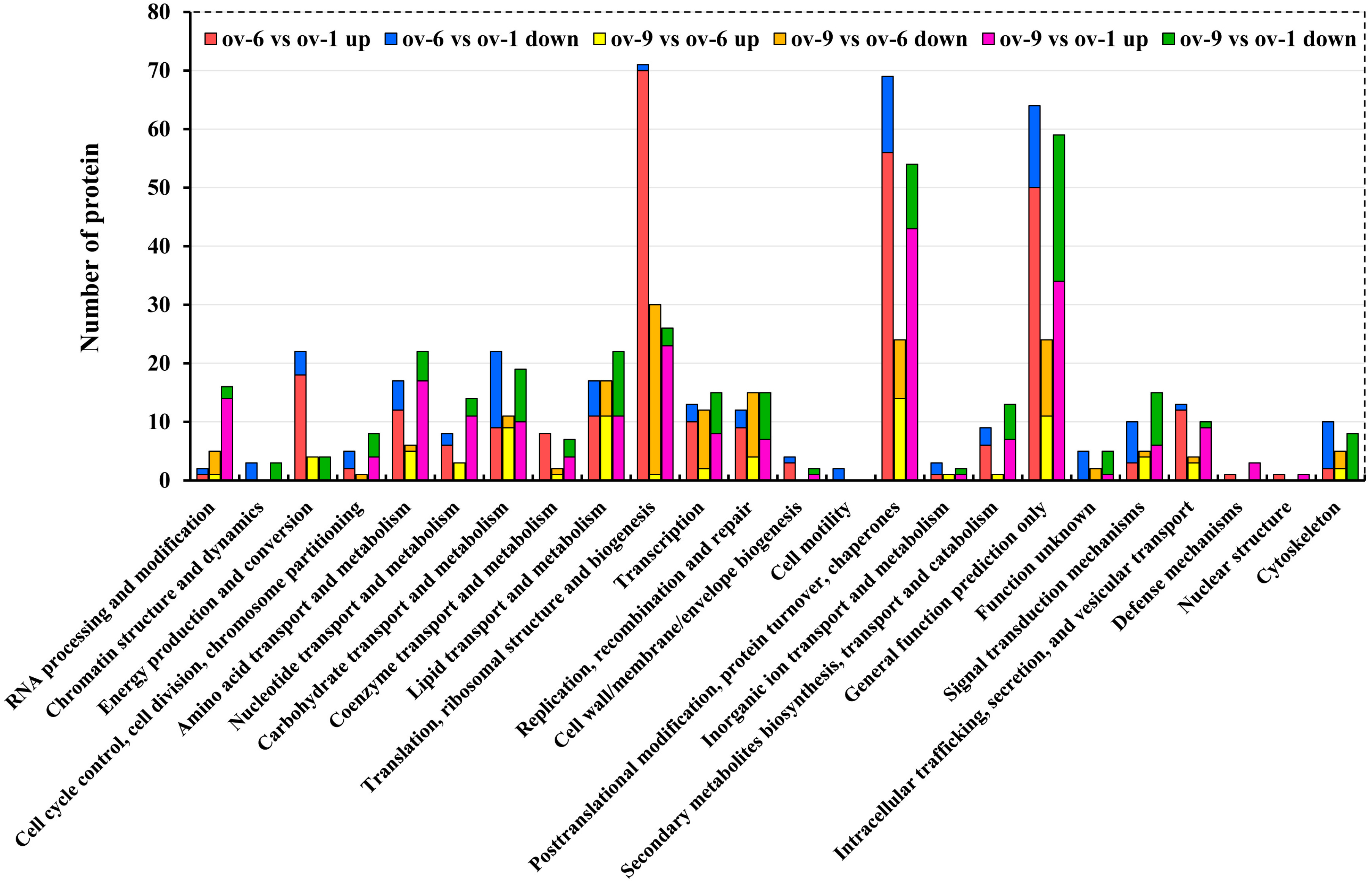
2.2. Functional Annotation of Proteins
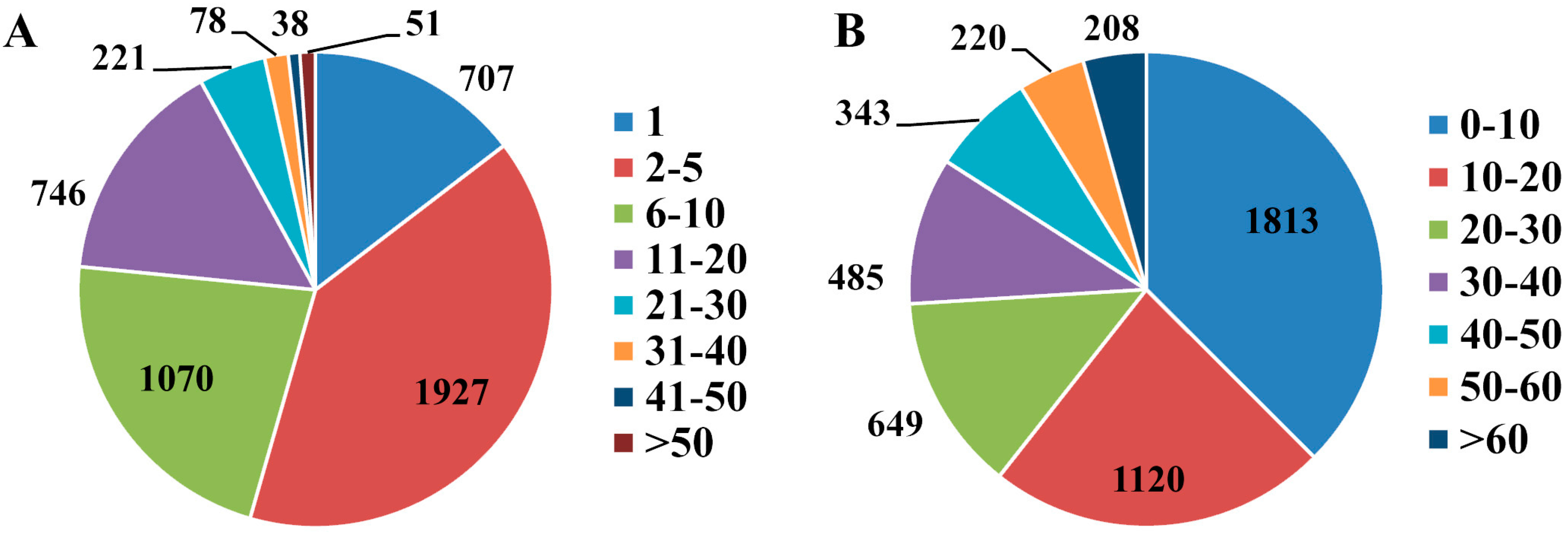
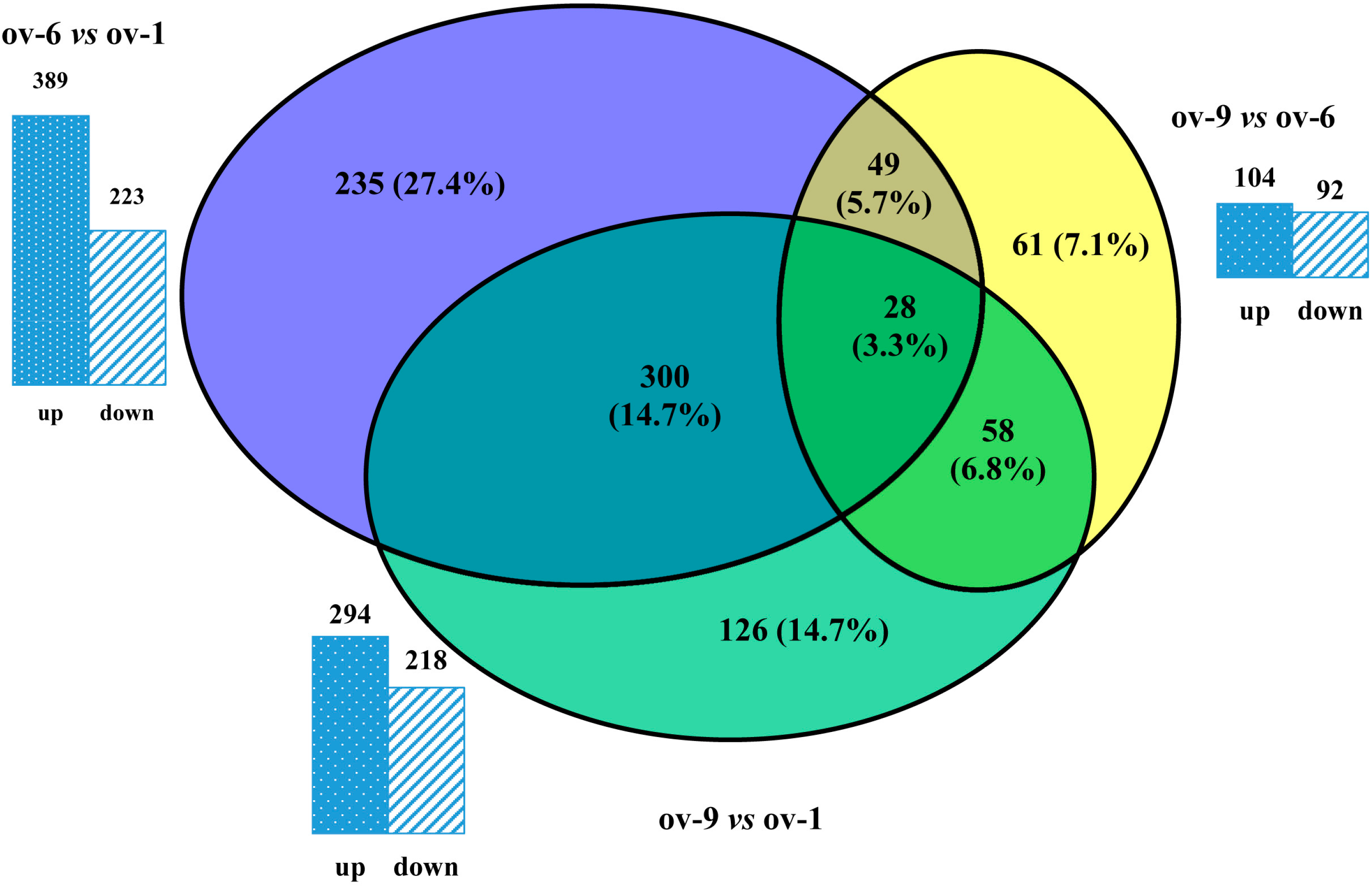
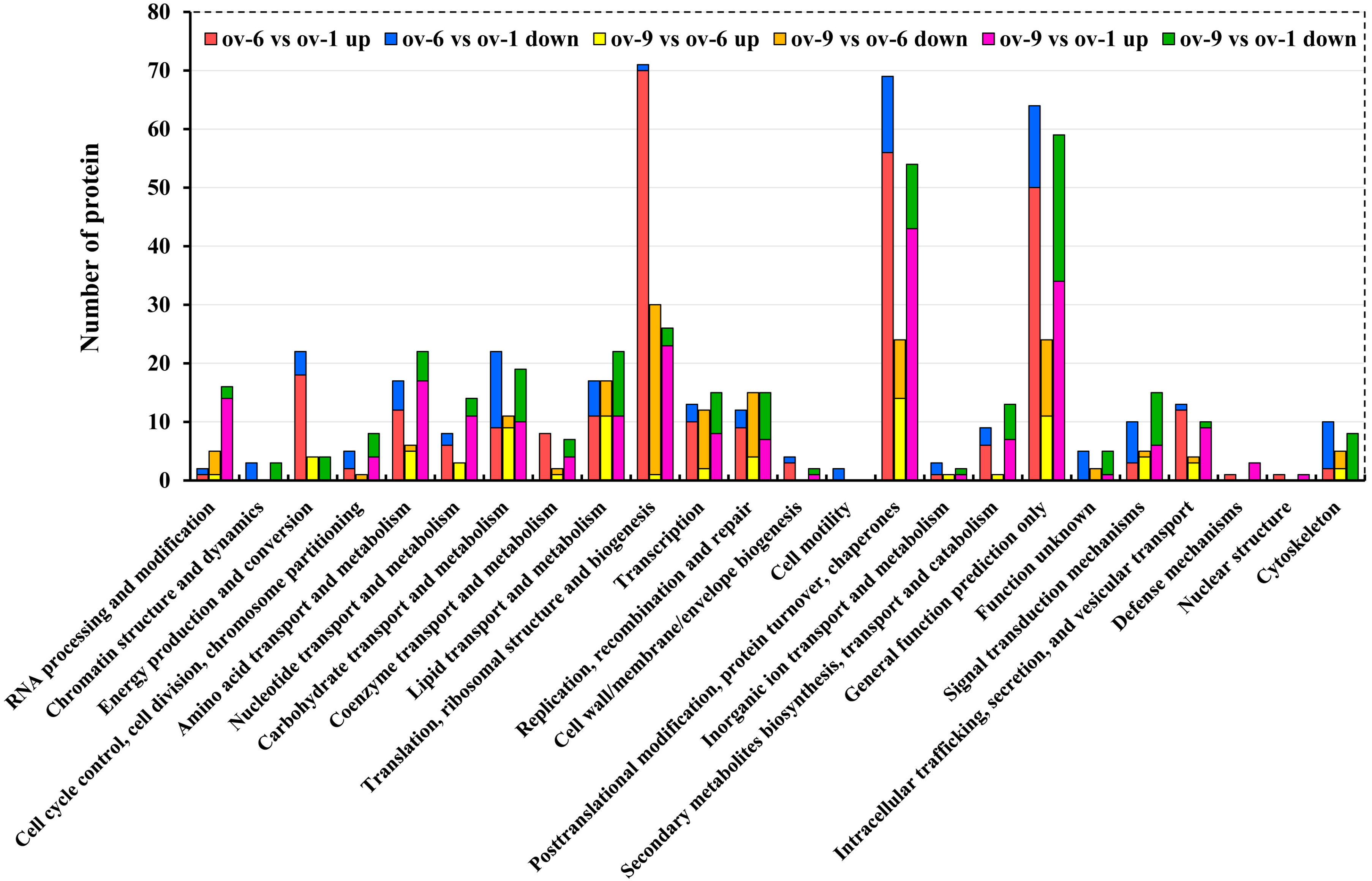
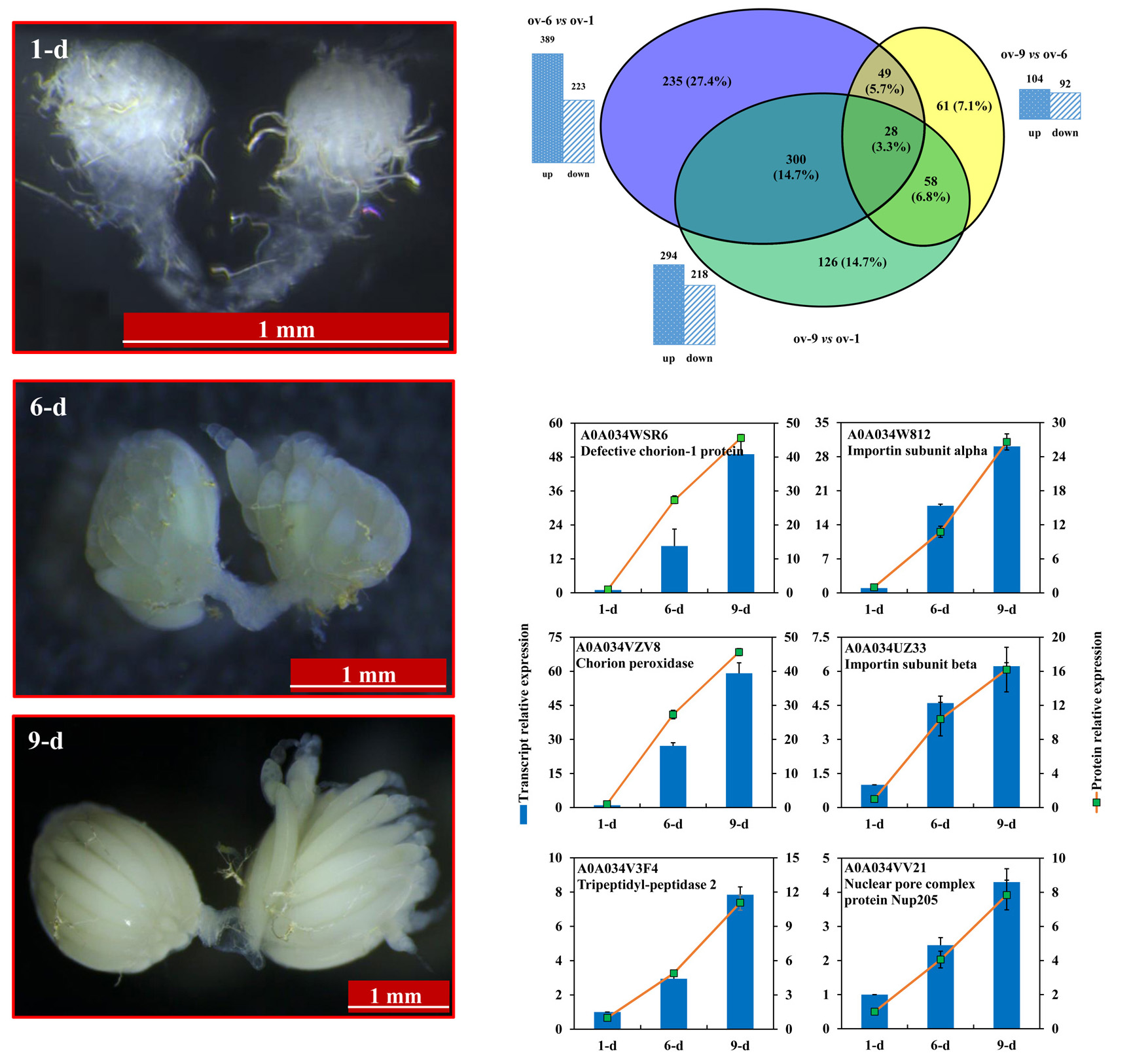
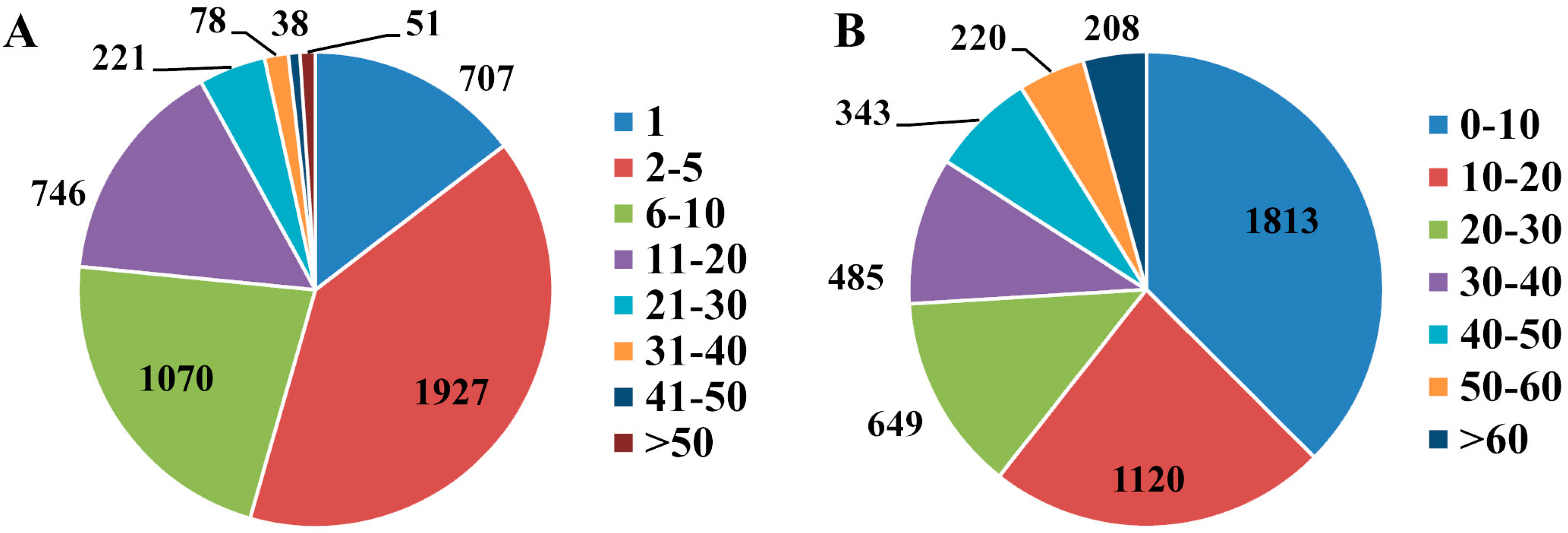
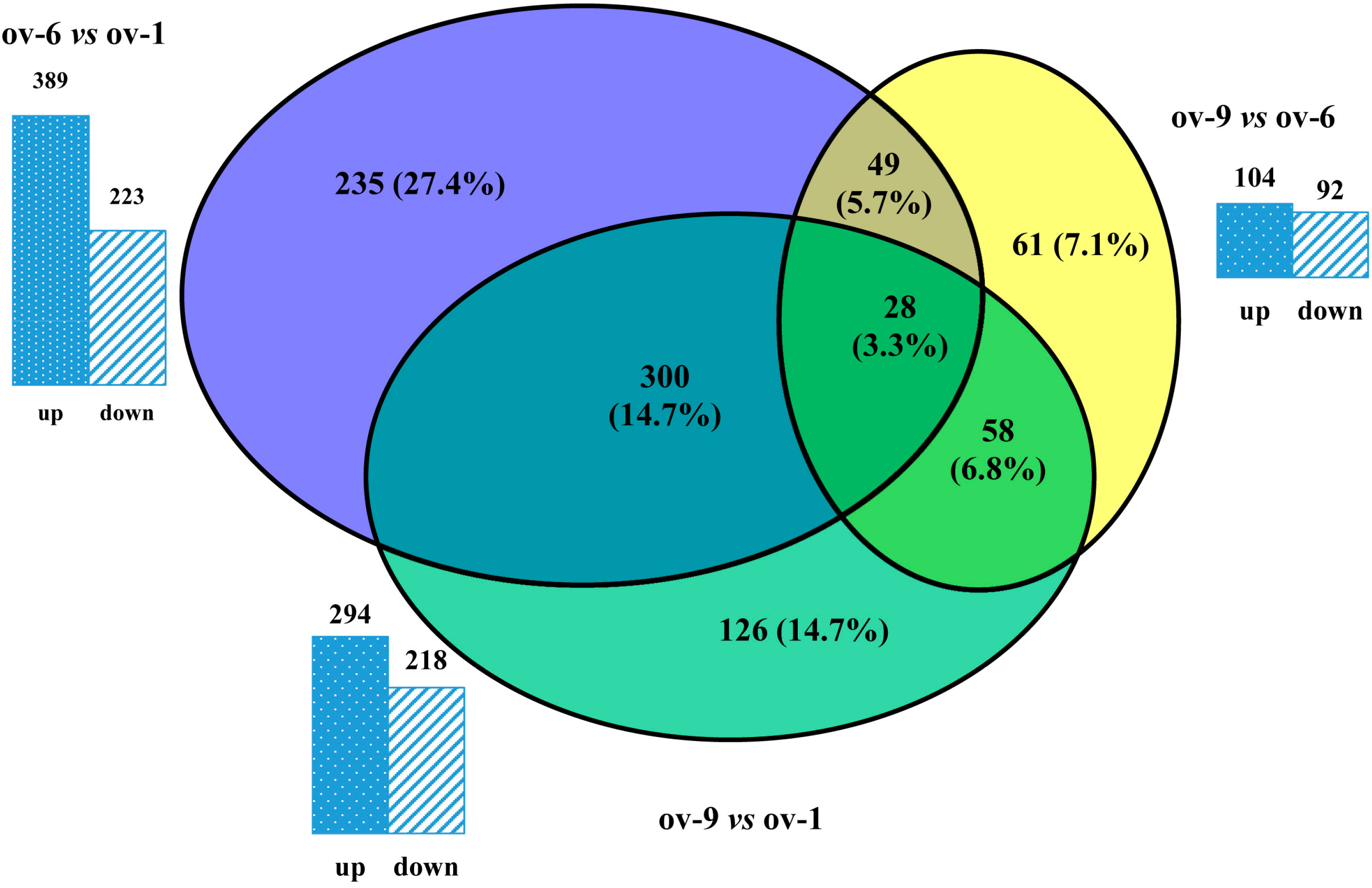
2.3. Differentially Expressed Proteins
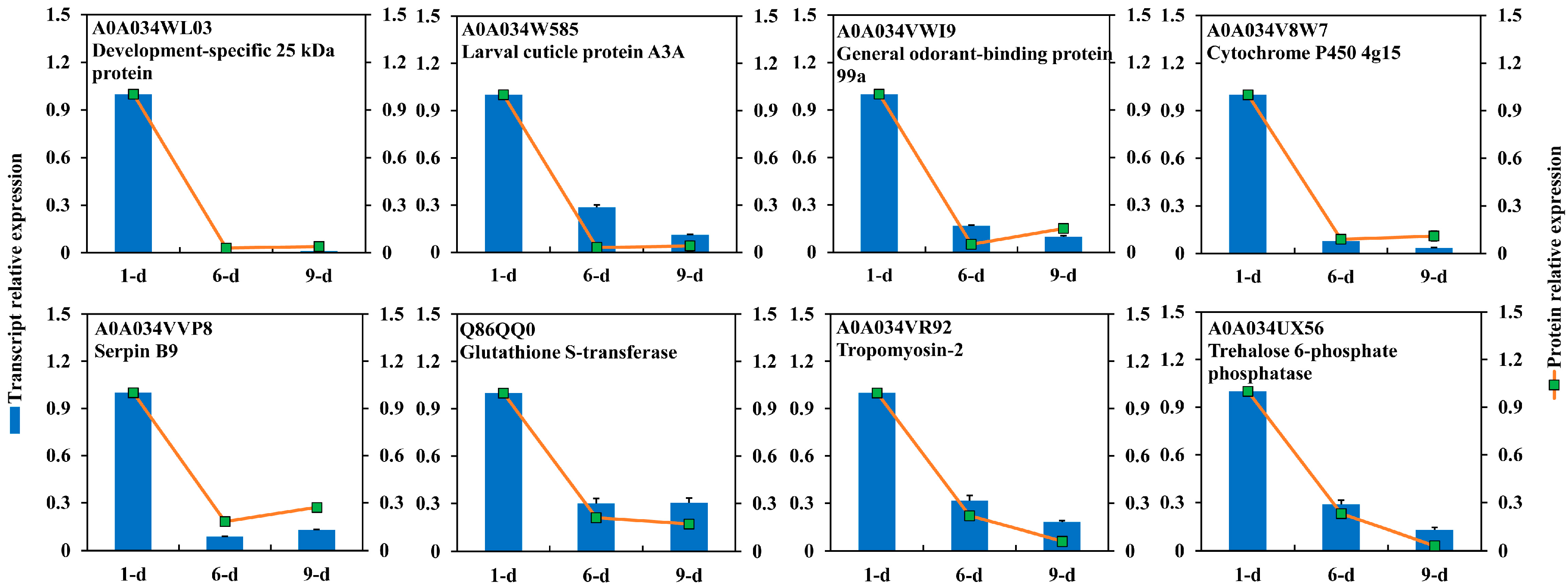
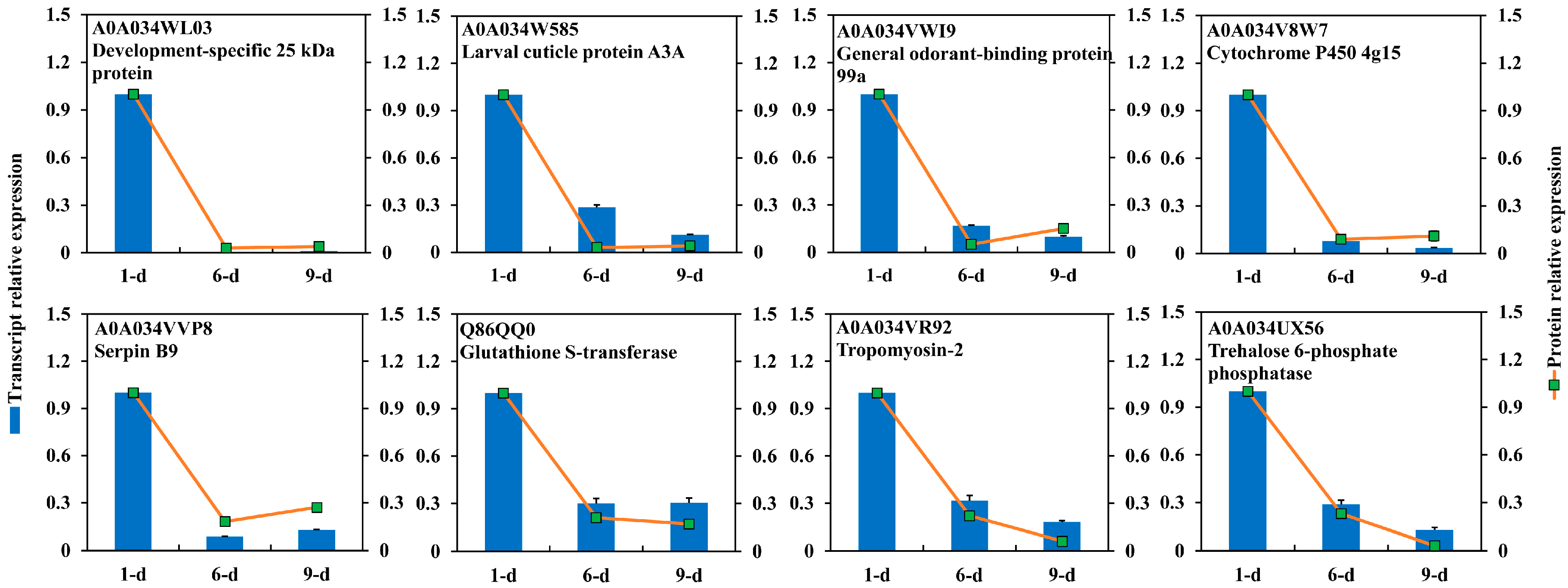
2.4. DEPs Highly Abundant Ovary of 1-Day-Old Adult
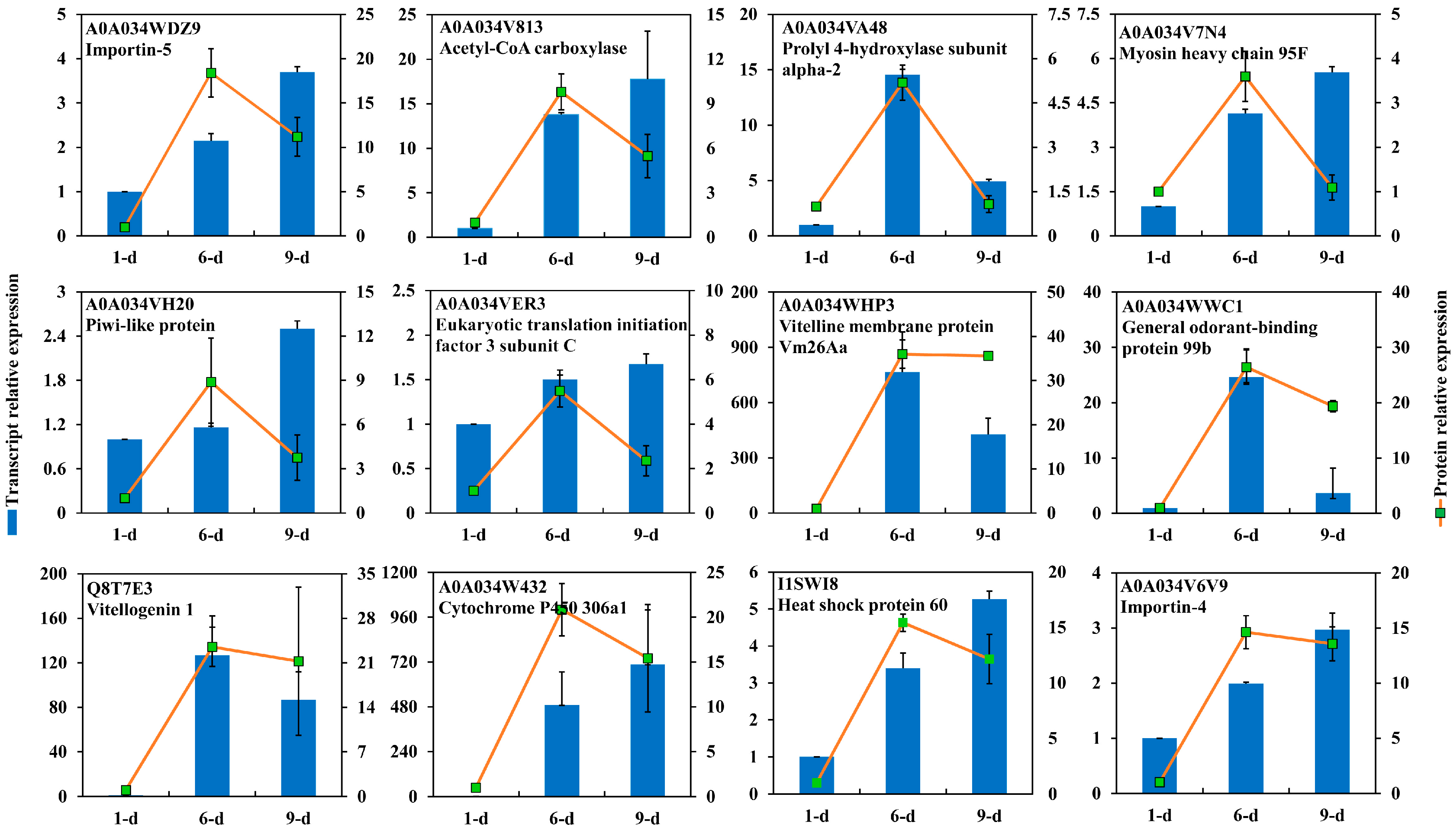
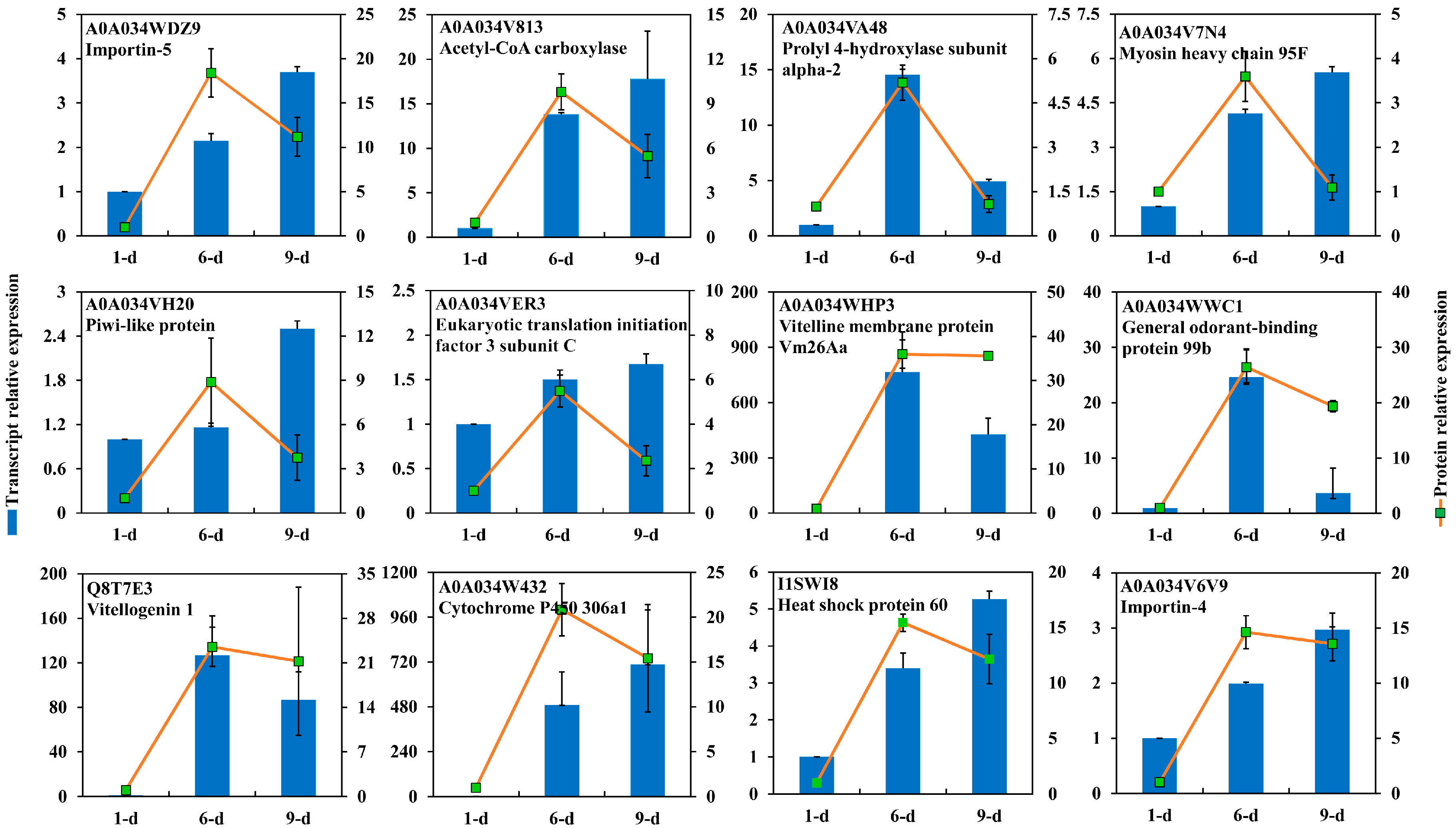
2.5. DEPs Highly Abundant in Ovary of 6-Day-Old Adult
2.6. DEPs Highly Abundant in Ovary of 9-Day-Old Adult
3. Materials and Methods
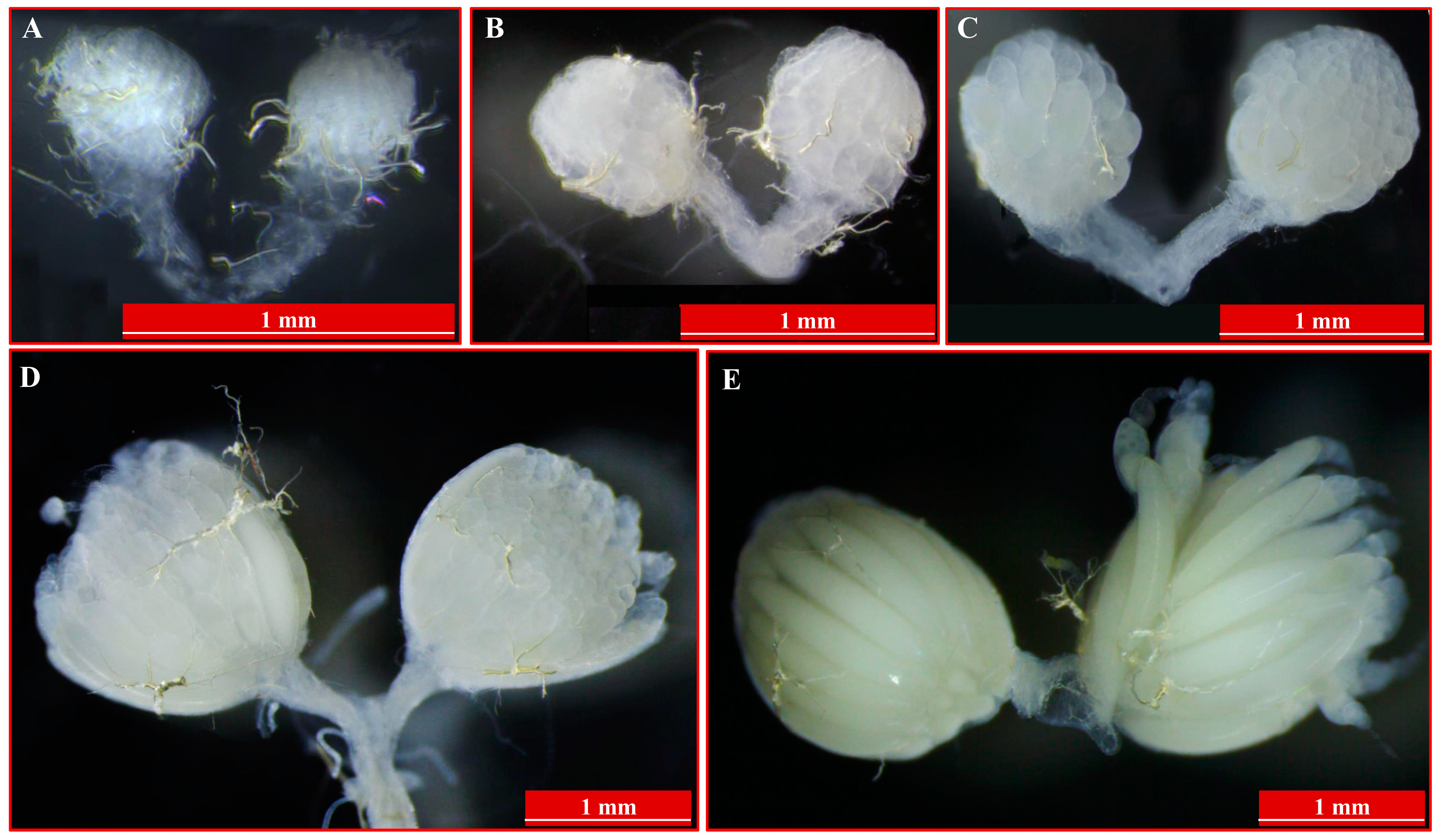
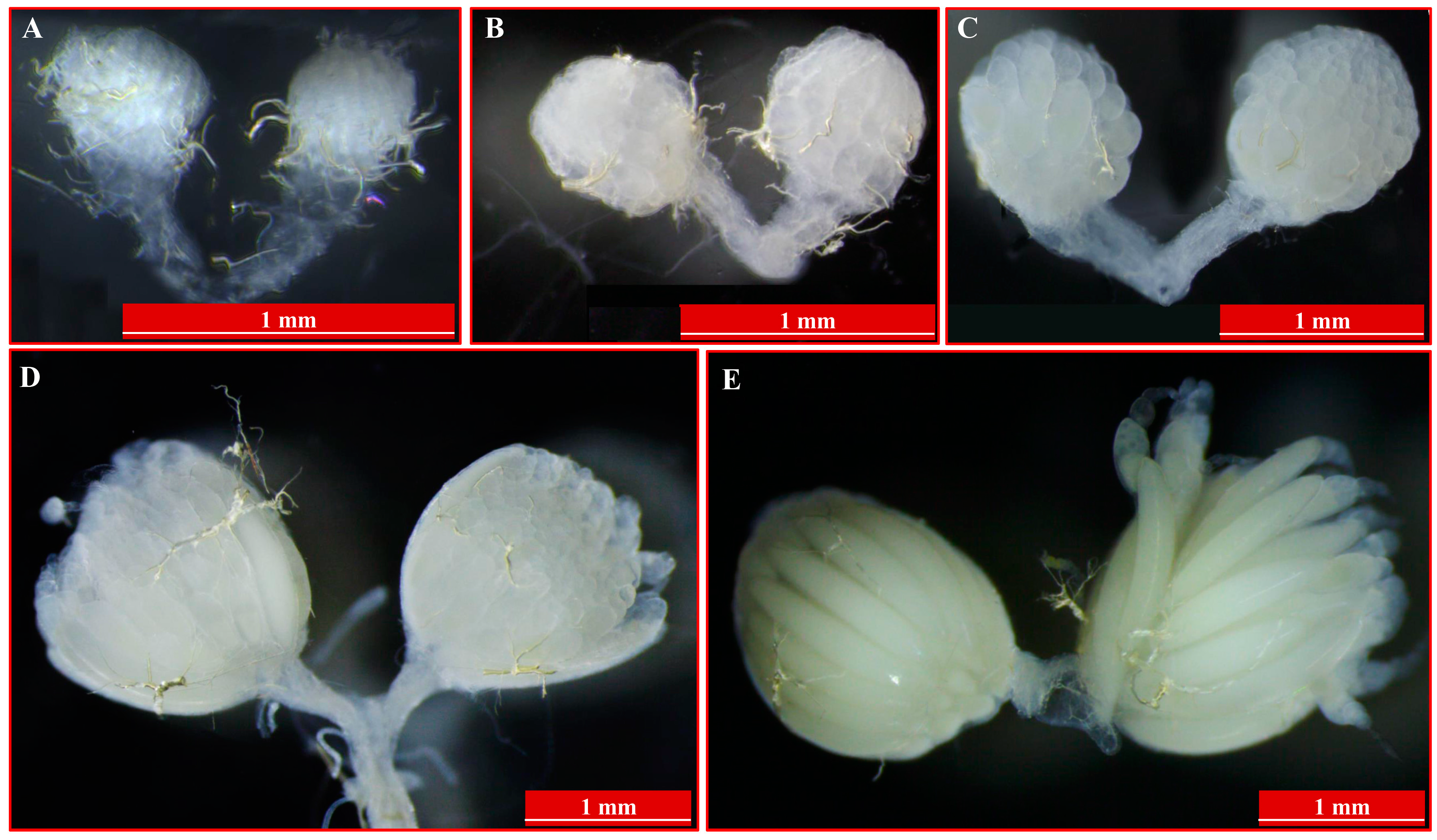
3.1. Insects Culture
3.2. Protein Extraction
3.3. Sample Digestion and Labeling
3.4. LC-MS/MS Analysis
3.5. Protein Identification and Functional Annotation
3.6. Protein Quantitative Analysis
3.7. RNA Extraction and RT-qPCR Analysis
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Shelly, T.E.; Edu, J.; Pahio, E. Influence of diet and methyl eugenol on the mating success of males of the oriental fruit fly, Bactrocera dorsalis (Diptera: Tephritidae). Fla. Entomol. 2005, 88, 307–313. [Google Scholar] [CrossRef]
- Wei, D.; Feng, Y.C.; Wei, D.D.; Yuan, G.R.; Dou, W.; Wang, J.J. Female remating inhibition and fitness of Bactrocera dorsalis (Diptera: Tephritidae) associated with male accessory glands. Fla. Entomol. 2015, 98, 52–58. [Google Scholar] [CrossRef]
- Jin, T.; Zeng, L.; Lin, Y.Y.; Lu, Y.Y.; Liang, G.W. Insecticide resistance of the oriental fruit fly, Bactrocera dorsalis (Hendel) (Diptera: Tephritidae), in mainland China. Pest Manag. Sci. 2011, 67, 370–376. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.J.; Wei, D.; Dou, W.; Hu, F.; Liu, W.F.; Wang, J.J. Toxicities and synergistic effects of several insecticides against the oriental fruit fly (Diptera: Tephritidae). J. Econ. Entomol. 2013, 106, 970–978. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.J.; Meng, Q.Q.; Li, Z.Q.; Zhang, S.Q.; Zeng, L.; Lu, Y.Y. The resistance monitoring of Bactrocera dorsalis in the field of Shenzhen. Chin. Plant Prot. 2015, 35, 63–66. [Google Scholar]
- Zheng, W.W.; Peng, T.; He, W.; Zhang, H.Y. High-throughput sequencing to reveal genes involved in reproduction and development in Bactrocera dorsalis (Diptera: Tephritidae). PLoS ONE 2012, 7, e36463. [Google Scholar] [CrossRef] [PubMed]
- Geib, S.M.; Calla, B.; Hall, B.; Hou, S.; Manoukis, N.C. Characterizing the developmental transcriptome of the oriental fruit fly, Bactrocera dorsalis (Diptera: Tephritidae) through comparative genomic analysis with Drosophila melanogaster utilizing modENCODE datasets. BMC Genom. 2014, 15, 942. [Google Scholar] [CrossRef] [PubMed]
- Wei, D.; Li, H.M.; Yang, W.J.; Wei, D.D.; Dou, W.; Huang, Y.; Wang, J.J. Transcriptome profiling of the testis reveals genes involved in spermatogenesis and marker discovery in the oriental fruit fly, Bactrocera dorsalis. Insect Mol. Biol. 2015, 24, 41–57. [Google Scholar] [CrossRef] [PubMed]
- Wei, D.; Tian, C.B.; Liu, S.H.; Wang, T.; Smagghe, G.; Jia, F.X.; Dou, W.; Wang, J.J. Transcriptome analysis to identify genes for peptides and proteins involved in immunity and reproduction from male accessory glands and ejaculatory duct of Bactrocera dorsalis. Peptides 2016, 80, 48–60. [Google Scholar] [CrossRef] [PubMed]
- Takemori, N.; Yamamoto, M.T. Proteome mapping of the Drosophila melanogaster male reproductive system. Proteomics 2009, 9, 2484–2493. [Google Scholar] [CrossRef] [PubMed]
- Velentzas, A.D.; Anagnostopoulos, A.K.; Velentzas, P.D.; Mpakou, V.E.; Sagioglou, N.E.; Tsioka, M.M.; Katarachia, S.; Manta, A.K.; Konstantakou, E.G.; Papassideri, I.S.; et al. Global proteomic profiling of Drosophila ovary: A high-resolution, unbiased, accurate and multifaceted analysis. Cancer Genom. Proteom. 2015, 12, 369–384. [Google Scholar]
- Lin, H. The stem-cell niche theory: Lessons from flies. Nat. Rev. Genet. 2002, 3, 931–940. [Google Scholar] [CrossRef] [PubMed]
- Horne-Badovinac, S.; Bilder, D. Mass transit: Epithelial morphogenesis in the Drosophila egg chamber. Dev. Dynam. 2005, 232, 559–574. [Google Scholar] [CrossRef] [PubMed]
- Dobens, L.L.; Raftery, L.A. Integration of epithelial patterning and morphogenesis in Drosophila ovarian follicle cells. Dev. Dynam. 2000, 218, 80–93. [Google Scholar] [CrossRef]
- Hook, J.; Lemckert, F.; Qin, H.; Schevzov, G.; Gunning, P. Gamma tropomyosin gene products are required for embryonic development. Mol. Cell. Biol. 2004, 24, 2318–2323. [Google Scholar] [CrossRef] [PubMed]
- Jang, W.I.; Jo, Y.J.; Kim, H.C.; Jia, J.L.; Namgoong, S.; Kim, N.H. Non-muscle tropomyosin (Tpm3) is crucial for asymmetric cell division and maintenance of cortical integrity in mouse oocytes. Cell Cycle 2014, 13, 2359–2369. [Google Scholar] [CrossRef] [PubMed]
- Clayton, L.; Johnson, M.H. Tropomyosin in preimplantation mouse development: Identification, expression, and organization during cell division and polarization. Exp. Cell Res. 1998, 238, 450–464. [Google Scholar] [CrossRef] [PubMed]
- Erdelyi, M.; Michon, A.M.; Guichet, A.; Glotzer, J.B.; Ephrussi, A. Requirement for Drosophila cytoplasmic tropomyosin in oskar mRNA localization. Nature 1995, 377, 524–527. [Google Scholar] [CrossRef] [PubMed]
- Tetzlaff, M.T.; Jackle, H.; Pankratz, M.J. Lack of Drosophila cytoskeletal tropomyosin affects head morphogenesis and the accumulation of oskar mRNA required for germ cell formation. EMBO J. 1996, 15, 1247–1254. [Google Scholar] [PubMed]
- Young, P.E.; Pesacreta, T.C.; Kiehart, D.P. Dynamic changes in the distribution of cytoplasmic myosin during Drosophila embryogenesis. Development 1991, 111, 1–14. [Google Scholar] [PubMed]
- Wheatley, S.; Kulkarni, S.; Karess, R. Drosophila nonmuscle myosin II is required for rapid cytoplasmic transport during oogenesis and for axial nuclear migration in early embryos. Development 1995, 121, 1937–1946. [Google Scholar] [PubMed]
- Edwards, K.A.; Kiehart, D.P. Drosophila nonmuscle myosin II has multiple essential roles in imaginal disc and egg chamber morphogenesis. Development 1996, 122, 1499–1511. [Google Scholar] [PubMed]
- Jordan, P.; Karess, R. Myosin light chain–activating phosphorylation sites are required for oogenesis in Drosophila. J. Cell Biol. 1997, 139, 1805–1819. [Google Scholar] [CrossRef] [PubMed]
- Krauss, J.; de Quinto, S.L.; Nüsslein-Volhard, C.; Ephrussi, A. Myosin-V regulates oskar mRNA localization in the Drosophila oocyte. Curr. Biol. 2009, 19, 1058–1063. [Google Scholar] [CrossRef] [PubMed]
- Deng, W.; Leaper, K.; Bownes, M. A targeted gene silencing technique shows that Drosophila myosin VI is required for egg chamber and imaginal disc morphogenesis. J. Cell Sci. 1999, 112, 3677–3690. [Google Scholar] [PubMed]
- Zhou, X.H.; Ban, L.P.; Iovinella, I.; Zhao, L.J.; Gao, Q.; Felicioli, A.; Sagona, S.; Pieraccini, G.; Pelosi, P.; Zhang, L. Diversity, abundance, and sex-specific expression of chemosensory proteins in the reproductive organs of the locust Locusta migratoria manilensis. Biol. Chem. 2013, 394, 43–54. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.L.; Huang, L.Q.; Pelosi, P.; Wang, C.Z. Expression in antennae and reproductive organs suggests a dual role of an odorant-binding protein in two sibling Helicoverpa species. PLoS ONE 2012, 7, e30040. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Call, G.B.; Kirilly, D.; Xie, T. Notch signaling controls germline stem cell niche formation in the Drosophila ovary. Development 2007, 134, 1071–1080. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Gridley, T. Notch signaling during oogenesis in Drosophila melanogaster. Genet. Res. Int. 2012, 2012, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Rao, A.V.S.K.; Shaha, C. Role of glutathione S-transferases in oxidative stress–induced male germ cell apoptosis. Free Radic. Bio. Med. 2000, 29, 1015–1027. [Google Scholar] [CrossRef]
- Talakhun, W.; Roytrakul, S.; Phaonakrop, N.; Kittisenachai, S.; Khamnamtong, B.; Klinbunga, S.; Menasveta, P. Identification of reproduction-related proteins and characterization of the protein disulfide isomerase A6 cDNA in ovaries of the giant tiger shrimp Penaeus monodon. Comp. Biochem. Physiol. Part D Genom. Proteom. 2012, 7, 180–190. [Google Scholar] [CrossRef] [PubMed]
- Malone, C.D.; Brennecke, J.; Dus, M.; Stark, A.; McCombie, W.R.; Sachidanandam, R.; Hannon, G.J. Specialized piRNA pathways act in germline and somatic tissues of the Drosophila ovary. Cell 2009, 137, 522–535. [Google Scholar] [CrossRef] [PubMed]
- Czech, B.; Preall, J.; Mcginn, J.; Hannon, G. A transcriptome-wide RNAi screen in the Drosophila ovary reveals factors of the germline piRNA pathway. Mol. Cell 2013, 50, 749–761. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.C.; Valouev, A.; Liu, N.; Lin, H. Piwi maintains germline stem cells and oogenesis in Drosophila through negative regulation of Polycomb Group proteins. Nat. Genet. 2016, 48, 283–291. [Google Scholar] [CrossRef] [PubMed]
- Bardsley, A.; McDonald, K.; Boswell, R.E. Distribution of tudor protein in the Drosophila embryo suggests separation of functions based on site of localization. Development 1993, 119, 207–219. [Google Scholar] [PubMed]
- Creed, T.M.; Loganathan, S.N.; Varonin, D.; Jackson, C.A.; Arkov, A.L. Novel role of specific Tudor domains in Tudor—Aubergine protein complex assembly and distribution during Drosophila oogenesis. Biochem. Bioph. Res. Commun. 2010, 402, 384–389. [Google Scholar] [CrossRef] [PubMed]
- Dufourt, J.; Dennis, C.; Boivin, A.; Gueguen, N.; Théron, E.; Goriaux, C.; Pouchin, P.; Ronsseray, S.; Brasset, E.; Vaury, C. Spatio-temporal requirements for transposable element piRNA-mediated silencing during Drosophila oogenesis. Nucleic Acids Res. 2014, 42, 2512–2524. [Google Scholar] [CrossRef] [PubMed]
- Richter, J.D.; Sonenberg, N. Regulation of cap-dependent translation by eIF4E inhibitory proteins. Nature 2005, 433, 477–480. [Google Scholar] [CrossRef] [PubMed]
- Evsikov, A.V.; Graber, J.H.; Brockman, J.M.; Hampl, A.; Holbrook, A.E.; Singh, P.; Eppig, J.J.; Solter, D.; Knowles, B.B. Cracking the egg: Molecular dynamics and evolutionary aspects of the transition from the fully grown oocyte to embryo. Genes Dev. 2006, 20, 2713–2727. [Google Scholar] [CrossRef] [PubMed]
- Piccioni, F.; Zappavigna, V.; Verrotti, A.C. Translational regulation during oogenesis and early development: The cap-poly(A) tail relationship. CR. Biol. 2005, 328, 863–881. [Google Scholar] [CrossRef] [PubMed]
- Keyes, L.N.; Spradling, A.C. The Drosophila gene fs(2)cup interacts with otu to define a cytoplasmic pathway required for the structure and function of germ-line chromosomes. Development 1997, 124, 1419–1431. [Google Scholar] [PubMed]
- Zappavigna, V.; Piccioni, F.; Villaescusa, J.C.; Verrotti, A.C. Cup is a nucleocytoplasmic shuttling protein that interacts with the eukaryotic translation initiation factor 4E to modulate Drosophila ovary development. Proc. Natl. Acad. Sci. USA 2004, 101, 14800–14805. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Shu, J.; Zhang, Q. Expression of the Tribolium castaneum (Coleoptera: Tenebrionidae) hsp83 gene and its relation to oogenesis during ovarian maturation. J. Genet. Genom. 2010, 37, 513–522. [Google Scholar] [CrossRef]
- Pisa, V.; Cozzolino, M.; Gargiulo, S.; Ottone, C.; Piccioni, F.; Monti, M.; Gigliotti, S.; Talamo, F.; Graziani, F.; Pucci, P. The molecular chaperone Hsp90 is a component of the cap-binding complex and interacts with the translational repressor Cup during Drosophila oogenesis. Gene 2009, 432, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Marin, R.; Tanguay, R.M. Stage-specific localization of the small heat shock protein Hsp27 during oogenesis in Drosophila melanogaster. Chromosoma 1996, 105, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Economou, K.; Kotsiliti, E.; Mintzas, A.C. Stage and cell-specific expression and intracellular localization of the small heat shock protein Hsp27 during oogenesis and spermatogenesis in the Mediterranean fruit fly, Ceratitis capitata. J. Insect Physiol. 2017, 96, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.W.; Li, X.; Yu, Q.Y.; Xiang, Z.H.; Kishino, H.; Zhang, Z. The small heat shock protein (sHSP) genes in the silkworm, Bombyx mori, and comparative analysis with other insect sHSP genes. BMC Evol. Biol. 2009, 9, 215. [Google Scholar] [CrossRef] [PubMed]
- Ádám, G.; Gausz, J.; Noselli, S.; Kurucz, É.; Andó, I.; Udvardy, A. Tissue- and developmental stage-specific changes in the subcellular localization of the 26S proteasome in the ovary of Drosophila melanogaster. Gene Expr. Patterns 2004, 4, 329–333. [Google Scholar] [CrossRef] [PubMed]
- Fredriksson, Å.; Johansson Krogh, E.; Hernebring, M.; Pettersson, E.; Javadi, A.; Almstedt, A.; Nyström, T. Effects of aging and reproduction on protein quality control in soma and gametes of Drosophila melanogaster. Aging Cell 2012, 11, 634–643. [Google Scholar] [CrossRef] [PubMed]
- Tufail, M.; Takeda, M. Molecular characteristics of insect vitellogenins. J. Insect Physiol. 2008, 54, 1447–1458. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.L.; Lin, C.P.; Lu, K.H. cDNA isolation, expression, and hormonal regulation of yolk protein genes in the oriental fruit fly, Bactrocera dorsalis (Hendel) (Diptera: Tephritidae). J. Insect Physiol. 2012, 58, 763–770. [Google Scholar] [CrossRef] [PubMed]
- Cingolani, G.; Petosa, C.; Weis, K.; Müller, C.W. Structure of importin-β bound to the IBB domain of importin-α. Nature 1999, 399, 221–229. [Google Scholar] [PubMed]
- Lippai, M.; Tirián, L.; Boros, I.; Mihály, J.; Erdélyi, M.; Belecz, I.; Máthé, E.; Pósfai, J.; Nagy, A.; Udvardy, A. The Ketel gene encodes a Drosophila homologue of importin-β. Genetics 2000, 156, 1889–1900. [Google Scholar] [PubMed]
- Máthé, E.; Bates, H.; Huikeshoven, H.; Deák, P.; Glover, D.M.; Cotterill, S. Importin-α3 is required at multiple stages of Drosophila development and has a role in the completion of oogenesis. Dev. Biol. 2000, 223, 307–322. [Google Scholar] [CrossRef] [PubMed]
- Gorjánácz, M.; Adám, G.; Török, I.; Mechler, B.M.; Szlanka, T.; Kiss, I. Importin-α 2 is critically required for the assembly of ring canals during Drosophila oogenesis. Dev. Biol. 2002, 251, 271–282. [Google Scholar] [CrossRef] [PubMed]
- Mason, D.A.; Fleming, R.J.; Goldfarb, D.S. Drosophila melanogaster importin α1 and α3 can replace importin α2 during spermatogenesis but not oogenesis. Genetics 2002, 161, 157–170. [Google Scholar] [PubMed]
- Ratan, R.; Mason, D.A.; Sinnot, B.; Goldfarb, D.S.; Fleming, R.J. Drosophila importin α1 performs paralog-specific functions essential for gametogenesis. Genetics 2008, 178, 839–850. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.C.; Geisbrecht, E.R. “Importin” signaling roles for import proteins: The function of Drosophila importin-7 (DIM-7) in muscle-tendon signaling. Cell Adhes. Migr. 2012, 6, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Waring, G.L. Morphogenesis of the eggshells in Drosophila. Int. Rev. Cytol. 2000, 198, 67–108. [Google Scholar] [PubMed]
- Cavaliere, V.; Bernardi, F.; Romani, P.; Duchi, S.; Gargiulo, G. Building up the Drosophila eggshell: First of all the eggshell genes must be transcribed. Dev. Dynam. 2008, 237, 2061–2072. [Google Scholar] [CrossRef] [PubMed]
- Chen, A.; Gao, P.; Zhao, Q.; Tang, S.; Shen, X.; Zhang, G.; Qiu, Z.; Xia, D.; Huang, Y.; Xu, Y.; et al. Mutation of a vitelline membrane protein, BmEP80, is responsible for the silkworm “Ming” lethal egg mutant. Gene 2013, 515, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Sdralia, N.; Swevers, L.; Iatrou, K. BmVMP90, a large vitelline membrane protein of the domesticated silkmoth Bombyx mori, is an essential component of the developing ovarian follicle. Insect Biochem. Mol. Biol. 2012, 42, 717–727. [Google Scholar] [CrossRef] [PubMed]
- Kendirgi, F.; Swevers, L.; Iatrou, K. An ovarian follicular epithelium protein of the silkworm (Bombyx mori) that associates with the vitelline membrane and contributes to the structural integrity of the follicle. FEBS Lett. 2002, 524, 59–68. [Google Scholar] [CrossRef]
- Velentzas, A.D.; Velentzas, P.D.; Sagioglou, N.E.; Konstantakou, E.G.; Anagnostopoulos, A.K.; Tsioka, M.M.; Mpakou, V.E.; Kollia, Z.; Consoulas, C.; Margaritis, L.H. Targeted downregulation of s36 protein unearths its cardinal role in chorion biogenesis and architecture during Drosophila melanogaster oogenesis. Sci. Rep. 2016, 6, 35511. [Google Scholar] [CrossRef] [PubMed]
- Konstandi, O.A.; Papassideri, I.S.; Stravopodis, D.J.; Kenoutis, C.A.; Hasan, Z.; Katsorchis, T.; Wever, R.; Margaritis, L.H. The enzymatic component of Drosophila melanogaster chorion is the Pxd peroxidase. Insect Biochem. Mol. Biol. 2005, 35, 1043–1057. [Google Scholar] [CrossRef] [PubMed]
- Konstandi, O.A.; Papassideri, I.S.; Stravopodis, D.J.; Antonelou, M.H.; Kenoutis, C.A.; Stefanidou, D.C.; Margaritis, L.H. The dual role of chorion peroxidase in Bactrocera oleae chorion assembly. Int. J. Dev. Biol. 2006, 50, 543–552. [Google Scholar] [CrossRef] [PubMed]
- Buszczak, M.; Freeman, M.R.; Carlson, J.R.; Bender, M.; Cooley, L.; Segraves, W.A. Ecdysone response genes govern egg chamber development during mid-oogenesis in Drosophila. Development 1999, 126, 4581–4589. [Google Scholar] [PubMed]
- Bernardi, F.; Romani, P.; Tzertzinis, G.; Gargiulo, G.; Cavaliere, V. EcR-B1 and Usp nuclear hormone receptors regulate expression of the VM32E eggshell gene during Drosophila oogenesis. Dev. Biol. 2009, 328, 541–551. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.K.; Yang, W.J.; Tian, Y.; Wu, Y.B.; Wang, J.J. Insulin signaling pathway in the oriental fruit fly: The role of insulin receptor substrate in ovarian development. Gen. Comp. Endocr. 2015, 126, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Abrisqueta, M.; Sueren-Castillo, S.; Maestro, J.L. Insulin receptor-mediated nutritional signalling regulates juvenile hormone biosynthesis and vitellogenin production in the German cockroach. Insect Biochem. Mol. Biol. 2014, 49, 14–23. [Google Scholar] [CrossRef] [PubMed]
- Vanorny, D.A.; Prasasya, R.D.; Chalpe, A.J.; Kilen, S.M.; Mayo, K.E. Notch signaling regulates ovarian follicle formation and coordinates follicular growth. Mol. Endocrinol. 2014, 28, 499–511. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Shilov, I.V.; Seymour, S.L.; Patel, A.A.; Loboda, A.; Tang, W.H.; Keating, S.P.; Hunter, C.L.; Nuwaysir, L.M.; Schaeffer, D.A. The paragon algorithm, a next generation search engine that uses sequence temperature values and feature probabilities to identify peptides from tandem mass spectra. Mol. Cell. Proteom. 2007, 6, 1638–1655. [Google Scholar] [CrossRef] [PubMed]
- Conesa, A.; Götz, S.; García-Gómez, J.M.; Terol, J.; Talón, M.; Robles, M. Blast2GO: A universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 2005, 21, 3674–3676. [Google Scholar] [CrossRef] [PubMed]
- Shen, G.M.; Jiang, H.B.; Wang, X.N.; Wang, J.J. Evaluation of endogenous references for gene expression profiling in different tissues of the oriental fruit fly Bactrocera dorsalis (Diptera: Tephritidae). BMC Mol. Biol. 2010, 11, 76. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein ID | Annotation | Length | Coverage | Peptides | Ov-6/Ov-1 | Ov-9/Ov-1 | Ov-9/Ov-6 a,b |
|---|---|---|---|---|---|---|---|
| A0A034WJQ4 | Fat-body protein 1 | 511 | 48.53 | 31 | ↓ 0.02 ± 0 | ↓ 0.02 ± 0 | ↑ 50.24 ± 28.2 a |
| A0A034V3B3 | Arylphorin subunit A4 | 1224 | 45.92 | 55 | ↓ 0.02 ± 0 | ↓ 0.04 ± 0.01 | ↑ 4.95 ± 2.14 a |
| A0A034VYM4 | Larval serum protein 1 gamma chain | 773 | 31.31 | 27 | ↓ 0.02 ± 0 | ↓ 0.03 ± 0 | ↑ 2.95 ± 1.03 a |
| A0A034V6N3 | High mobility group protein Z | 112 | 33.04 | 5 | ↓ 0.02 ± 0 | ↓ 0.02 ± 0 | ↑ 1.87 ± 0.47 a |
| A0A034WL03 | Development-specific 25 kDa protein | 259 | 69.5 | 19 | ↓ 0.03 ± 0 | ↓ 0.04 ± 0 | ↑ 3.04 ± 1 a |
| A0A034W585 | Larval cuticle protein A3A | 138 | 75.36 | 17 | ↓ 0.03 ± 0 | ↓ 0.04 ± 0 | ↑ 2.3 ± 0.55 a |
| A0A034WSW4 | Larval cuticle protein 8 | 104 | 22.12 | 2 | ↓ 0.03 ± 0 | ↓ 0.04 ± 0.01 | ↑ 1.98 ± 0.23 a |
| A0A034WXX0 | Histone H1 | 240 | 30.83 | 11 | ↓ 0.03 ± 0.01 | ↓ 0.03 ± 0 | ↑ 8.18 ± 4.64 a |
| A0A034WP07 | Attacin-A | 239 | 72.8 | 14 | ↓ 0.03 ± 0.01 | ↓ 0.13 ± 0.03 | ↑ 6.37 ± 1.87 a |
| A0A034V3D6 | J domain-containing protein | 177 | 70.62 | 15 | ↓ 0.04 ± 0.01 | ↓ 0.03 ± 0 | 0.84 ± 0.25 b |
| A0A034W4Q0 | Larval cuticle protein A2B | 180 | 73.89 | 17 | ↓ 0.05 ± 0 | ↓ 0.06 ± 0 | ↑ 1.61 ± 0.32 a |
| A0A034VT34 | Angiopoietin-4 | 454 | 20.04 | 7 | ↓ 0.05 ± 0 | ↓ 0.06 ± 0.01 | 1.39 ± 0.36 b |
| A0A034W9Y1 | Larval cuticle protein A2B | 250 | 92.4 | 19 | ↓ 0.05 ± 0.01 | ↓ 0.06 ± 0 | ↑ 1.73 ± 0.6 a |
| A0A034VU29 | Regucalcin | 351 | 64.96 | 26 | ↓ 0.05 ± 0.01 | ↓ 0.05 ± 0.01 | 1.49 ± 0.42 b |
| I1T1H2 | Heat shock protein 20 | 170 | 67.65 | 10 | ↓ 0.05 ± 0.02 | ↓ 0.13 ± 0.03 | ↑ 4.75 ± 1.43 a |
| A0A034VSB5 | Fasciclin-1 | 542 | 35.42 | 17 | ↓ 0.07 ± 0 | ↓ 0.56 ± 0.04 | ↑ 8.35 ± 0.24 |
| A0A034WRI9 | Sarcocystatin-A | 125 | 47.2 | 6 | ↓ 0.07 ± 0.02 | ↓ 0.15 ± 0.04 | ↑ 3.48 ± 1.58 a |
| A0A034WF27 | TPPP family protein CG4893 | 201 | 40.3 | 7 | ↓ 0.08 ± 0.01 | ↓ 0.09 ± 0.02 | 1.12 ± 0.28 b |
| A0A034VUV3 | Lamin Dm0 | 617 | 71.64 | 46 | ↓ 0.08 ± 0.02 | ↓ 0.06 ± 0 | 0.8 ± 0.28 b |
| A0A034WSV2 | Lamin-C | 622 | 56.27 | 37 | ↓ 0.09 ± 0.01 | ↓ 0.06 ± 0 | ↓ 0.63 ± 0.13 a |
| A0A034V8W7 | Cytochrome P450 4g15 | 484 | 23.76 | 10 | ↓ 0.09 ± 0.01 | ↓ 0.11 ± 0.03 | 1.27 ± 0.29 b |
| A0A034VPE5 | Tropomyosin-1, isoforms 9A/A/B | 188 | 69.68 | 19 | ↓ 0.09 ± 0.01 | ↓ 0.1 ± 0.02 | 0.92 ± 0.09 b |
| A0A034V2U0 | Protein 4.1-like protein | 1703 | 57.37 | 82 | ↓ 0.09 ± 0.02 | ↓ 0.07 ± 0 | 0.96 ± 0.24 b |
| A0A034VW09 | Protein Skeletor | 741 | 29.82 | 14 | ↓ 0.1 ± 0.01 | ↓ 0.21 ± 0.05 | ↑ 2.25 ± 0.46 a |
| A0A034W3W2 | Regucalcin | 296 | 42.91 | 12 | ↓ 0.1 ± 0.02 | ↓ 0.08 ± 0.01 | 0.87 ± 0.25 b |
| A0A034V7Q2 | Protein yellow | 261 | 31.42 | 6 | ↓ 0.1 ± 0.03 | ↓ 0.13 ± 0.01 | 1.48 ± 0.41 b |
| A0A034VI82 | Tropomodulin | 386 | 34.97 | 10 | ↓ 0.11 ± 0 | ↓ 0.08 ± 0 | ↓ 0.62 ± 0.07 a |
| A0A034VUS5 | Integrin-linked protein kinase | 448 | 32.37 | 14 | ↓ 0.12 ± 0.01 | ↓ 0.13 ± 0.02 | 1.15 ± 0.3 b |
| A0A034VKK9 | Alcohol dehydrogenase | 324 | 38.89 | 13 | ↓ 0.12 ± 0.01 | ↓ 0.13 ± 0.02 | 0.94 ± 0.2 b |
| A0A034VYW9 | Contactin | 1393 | 25.2 | 27 | ↓ 0.12 ± 0.02 | ↓ 0.21 ± 0.04 | ↑ 2.08 ± 0.42 a |
| A0A034WVT6 | Membrane metallo-endopeptidase-like 1 | 714 | 22.13 | 14 | ↓ 0.12 ± 0.02 | ↓ 0.15 ± 0.01 | 1.18 ± 0.19 b |
| A0A034V5Q1 | Ras suppressor protein 1 | 138 | 39.86 | 5 | ↓ 0.13 ± 0.02 | ↓ 0.19 ± 0.03 | ↑ 1.67 ± 0.41 a |
| A0A034W0B4 | Chitinase-like protein CG5210 | 279 | 51.25 | 14 | ↓ 0.13 ± 0.04 | ↓ 0.37 ± 0.04 | ↑ 4.97 ± 1.76 a |
| A0A034WBA7 | Putative G-protein coupled receptor 158 | 809 | 29.05 | 20 | ↓ 0.14 ± 0.02 | ↓ 0.28 ± 0.04 | ↑ 2.24 ± 0.56 a |
| A0A034V5R2 | Talin-1 | 1928 | 48.86 | 69 | ↓ 0.15 ± 0.01 | ↓ 0.45 ± 0.02 | ↑ 3.15 ± 0.09 |
| A0A034VQZ0 | Zinc finger protein 512B | 427 | 26.23 | 7 | ↓ 0.15 ± 0.01 | ↓ 0.13 ± 0.02 | 0.84 ± 0.16 b |
| A0A034VDB5 | Protein lap4 | 915 | 20.11 | 14 | ↓ 0.16 ± 0 | ↓ 0.11 ± 0.01 | ↓ 0.6 ± 0.09 a |
| A0A034VTK3 | Chitinase-like protein Idgf5 | 432 | 27.31 | 9 | ↓ 0.16 ± 0.05 | ↓ 0.16 ± 0.01 | 1.27 ± 0.41 b |
| A0A034W8P6 | Protein E(Sev)2B | 211 | 43.6 | 9 | ↓ 0.17 ± 0.03 | ↓ 0.29 ± 0.02 | ↑ 2.05 ± 0.44 a |
| A0A034VVP8 | Serpin B9 | 504 | 29.76 | 14 | ↓ 0.18 ± 0.01 | ↓ 0.27 ± 0.02 | 1.49 ± 0.12 b |
| A0A034W4R7 | α,α-trehalose-phosphate synthase A | 813 | 33.46 | 24 | ↓ 0.18 ± 0.02 | ↓ 0.09 ± 0.01 | ↓ 0.41 ± 0.06 a |
| A0A034WI29 | Protein lin-7-like protein B | 195 | 28.72 | 5 | ↓ 0.18 ± 0.03 | ↓ 0.19 ± 0.02 | 1.1 ± 0.18 b |
| A0A034VY88 | Protein hu-li tai shao | 698 | 69.63 | 35 | ↓ 0.19 ± 0.01 | ↓ 0.2 ± 0.02 | 1.07 ± 0.09 b |
| A0A034W1X2 | Alaserpin | 398 | 27.39 | 10 | ↓ 0.19 ± 0.02 | ↓ 0.42 ± 0.03 | ↑ 2.43 ± 0.28 a |
| A0A034W8L1 | Flotillin-2 | 425 | 43.53 | 17 | ↓ 0.19 ± 0.02 | ↓ 0.13 ± 0.02 | ↓ 0.63 ± 0.08 a |
| A0A034VQR8 | Troponin T, skeletal muscle | 384 | 39.32 | 22 | ↓ 0.19 ± 0.02 | ↓ 0.07 ± 0.01 | ↓ 0.31 ± 0.07 a |
| A0A034WRQ2 | α-parvin | 366 | 33.06 | 10 | ↓ 0.19 ± 0.05 | ↓ 0.26 ± 0.05 | ↑ 1.58 ± 0.39 a |
| A0A034VP83 | Poly(U)-specific endoribonuclease-like protein | 675 | 24.74 | 13 | ↓ 0.2 ± 0.01 | ↓ 0.35 ± 0.03 | ↑ 1.79 ± 0.16 a |
| A0A034WSM6 | Guanine nucleotide-binding protein G(S) subunit α | 382 | 32.2 | 10 | ↓ 0.2 ± 0.02 | ↓ 0.36 ± 0.02 | ↑ 1.88 ± 0.18 a |
| Q86QQ0 | Glutathione S-transferase | 209 | 37.32 | 6 | ↓ 0.21 ± 0.01 | ↓ 0.17 ± 0.01 | 0.81 ± 0.07 b |
| A0A034VND6 | Tropomyosin-1, isoforms 33/34 | 282 | 48.58 | 20 | ↓ 0.21 ± 0.04 | ↓ 0.13 ± 0.02 | ↓ 0.61 ± 0.13 a |
| A0A034VSX0 | Phosphate-regulating neutral endopeptidase | 683 | 22.69 | 13 | ↓ 0.21 ± 0.04 | ↓ 0.16 ± 0.02 | 0.85 ± 0.16 b |
| A0A034V6M3 | 60S ribosomal protein L23a | 266 | 26.32 | 10 | ↓ 0.21 ± 0.05 | ↓ 0.26 ± 0.07 | ↑ 1.62 ± 0.78 a |
| A0A034UZS9 | PDZ and LIM domain protein | 495 | 22.22 | 9 | ↓ 0.22 ± 0.01 | ↓ 0.19 ± 0.04 | 0.87 ± 0.25 b |
| A0A034VR92 | Tropomyosin-2 | 284 | 57.04 | 17 | ↓ 0.22 ± 0.02 | ↓ 0.06 ± 0 | ↓ 0.22 ± 0.02 |
| A0A034V2K7 | Spectrin β chain, non-erythrocytic 5 | 4200 | 26.48 | 94 | ↓ 0.22 ± 0.02 | ↓ 0.29 ± 0.03 | 1.32 ± 0.04 b |
| A0A034VDP0 | Obscurin | 631 | 30.74 | 13 | ↓ 0.22 ± 0.03 | ↓ 0.15 ± 0.02 | 0.71 ± 0.19 b |
| A0A034UX56 | Trehalose 6-phosphate phosphatase | 274 | 81.75 | 25 | ↓ 0.23 ± 0.01 | ↓ 0.03 ± 0 | ↓ 0.08 ± 0.01 |
| A0A034VE41 | Twitchin | 4978 | 23.54 | 86 | ↓ 0.23 ± 0.02 | ↓ 0.14 ± 0.01 | ↓ 0.57 ± 0.05 a |
| A0A034WWI9 | Ejaculatory bulb-specific protein 3 | 127 | 22.05 | 3 | ↓ 0.23 ± 0.02 | ↓ 0.18 ± 0.04 | 0.77 ± 0.14 b |
| A0A034VPA5 | Heterogeneous nuclear ribonucleoprotein Q | 417 | 21.34 | 9 | ↓ 0.23 ± 0.05 | ↓ 0.15 ± 0.01 | 0.69 ± 0.15 b |
| A0A034VWH3 | Myosin regulatory light chain 2 | 222 | 52.7 | 10 | ↓ 0.25 ± 0.02 | ↓ 0.12 ± 0.02 | ↓ 0.41 ± 0.06 a |
| A0A034VAC6 | Protein elav | 446 | 35.65 | 12 | ↓ 0.26 ± 0.02 | ↓ 0.29 ± 0.04 | 1.08 ± 0.17 b |
| A0A034V4T6 | Fasciclin-2 | 520 | 45.19 | 17 | ↓ 0.26 ± 0.03 | ↓ 0.08 ± 0.01 | ↓ 0.27 ± 0.02 a |
| A0A034VRQ6 | Protein held out wings | 356 | 37.64 | 11 | ↓ 0.28 ± 0.01 | ↓ 0.15 ± 0.01 | ↓ 0.48 ± 0.02 a |
| A0A034WML8 | Protein takeout | 269 | 20.07 | 4 | ↓ 0.29 ± 0.02 | ↓ 0.14 ± 0.01 | ↓ 0.46 ± 0.05 a |
| A0A034WF58 | Glutathione peroxidase | 278 | 48.92 | 13 | ↓ 0.29 ± 0.05 | ↓ 0.26 ± 0.03 | 0.94 ± 0.18 b |
| A0A034VTK4 | DNA topoisomerase 2 | 1481 | 42.13 | 59 | ↓ 0.3 ± 0.02 | ↓ 0.24 ± 0.01 | 0.76 ± 0.06 b |
| A0A034WS42 | Histone H4 | 103 | 65.05 | 15 | ↓ 0.3 ± 0.06 | ↓ 0.08 ± 0.01 | ↓ 0.24 ± 0.05 |
| A0A034WN98 | 17-β-hydroxysteroid dehydrogenase 13 | 326 | 25.15 | 7 | ↓ 0.33 ± 0.05 | ↓ 0.3 ± 0.07 | 0.98 ± 0.27 b |
| A0A034V7N7 | Protein lethal(2) giant larvae | 1176 | 24.23 | 21 | ↓ 0.33 ± 0.05 | ↓ 0.24 ± 0.04 | 0.67 ± 0.11 b |
| M1F3Z9 | Glutathione S-transferase ω-1 | 255 | 46.67 | 11 | ↓ 0.34 ± 0.02 | ↓ 0.26 ± 0.02 | 0.76 ± 0.06 b |
| A0A034VDX0 | L-2-hydroxyglutarate dehydrogenase, mitochondrial | 455 | 31.21 | 10 | ↓ 0.35 ± 0.03 | ↓ 0.33 ± 0.07 | 0.95 ± 0.23 b |
| A0A034VM61 | Clavesin-2 | 320 | 33.44 | 9 | ↓ 0.35 ± 0.04 | ↓ 0.45 ± 0.07 | 1.24 ± 0.16 b |
| A0A034VPR7 | Putative peptidyl-prolyl cis-trans isomerase dodo | 160 | 57.5 | 11 | ↓ 0.35 ± 0.06 | ↓ 0.32 ± 0 | 0.99 ± 0.18 b |
| A0A034WSZ7 | Heterochromatin protein 1 | 212 | 23.11 | 5 | ↓ 0.37 ± 0.02 | ↓ 0.24 ± 0.01 | ↓ 0.6 ± 0.01 a |
| A0A034WQQ0 | Calcyphosin-like protein | 219 | 27.4 | 6 | ↓ 0.37 ± 0.1 | ↓ 0.35 ± 0.06 | 1.04 ± 0.21 b |
| A0A034WBM1 | Acetyl-CoA acetyltransferase, cytosolic | 394 | 41.62 | 11 | ↓ 0.41 ± 0.08 | ↓ 0.45 ± 0.05 | 1.24 ± 0.3 b |
| A0A034VYI5 | Non-specific protein-tyrosine kinase | 510 | 27.25 | 12 | ↓ 0.45 ± 0.07 | ↓ 0.34 ± 0.01 | 0.79 ± 0.12 b |
| A0A034VW38 | SUN domain-containing protein 1 | 594 | 21.55 | 13 | ↓ 0.45 ± 0.1 | ↓ 0.31 ± 0.09 | 0.7 ± 0.14 b |
| A0A034VQG5 | Nucleoprotein TPR | 2410 | 26.22 | 53 | ↓ 0.47 ± 0.03 | ↓ 0.26 ± 0.01 | ↓ 0.49 ± 0.05 a |
| A0A034VKE6 | Bifunctional methylenetetrahydrofolate dehydrogenase/cyclohydrolase, mitochondrial | 316 | 32.59 | 8 | ↓ 0.5 ± 0.01 | ↓ 0.21 ± 0.05 | ↓ 0.43 ± 0.08 a |
| A0A034VPM4 | Vinculin | 960 | 50.63 | 38 | ↓ 0.54 ± 0.04 | ↓ 0.54 ± 0.04 | 0.98 ± 0.07 b |
| A0A034VRI7 | α-actinin, sarcomeric | 895 | 62.35 | 53 | ↓ 0.55 ± 0.04 | ↓ 0.42 ± 0.02 | 0.75 ± 0.04 b |
| Protein ID | Annotation | Length | Coverage | Peptides | Ov-6/Ov-1 | Ov-9/Ov-1 a,b | Ov-9/Ov-6 |
|---|---|---|---|---|---|---|---|
| A0A034V9A3 | CUGBP Elav-like family member 2 | 587 | 25.55 | 13 | ↑ 29.06 ± 4.88 | ↑ 13.14 ± 6.66 a | ↓ 0.37 ± 0.06 |
| A0A034WDZ9 | Importin-5 | 1106 | 53.35 | 47 | ↑ 18.4 ± 2.73 | ↑ 11.19 ± 4.34 | ↓ 0.53 ± 0.04 |
| A0A034VSG4 | Maternal protein exuperantia | 495 | 51.11 | 26 | ↑ 17 ± 1.02 | ↑ 5.37 ± 1.74 a | ↓ 0.27 ± 0.04 |
| A0A034WW15 | 40S ribosomal protein S3 | 244 | 80.33 | 26 | ↑ 16.58 ± 0.53 | ↑ 10.31 ± 1.82 | ↓ 0.54 ± 0.05 |
| A0A034VN85 | Eukaryotic translation initiation factor 3 subunit E | 434 | 38.71 | 15 | ↑ 11.54 ± 0.68 | ↑ 5.3 ± 2.25 a | ↓ 0.42 ± 0.08 |
| A0A034WLA5 | ATP-dependent RNA helicase vasa, isoform A | 621 | 57.49 | 35 | ↑ 11.45 ± 3.76 | ↑ 6.59 ± 4.91 a | ↓ 0.51 ± 0.04 |
| A0A034WFM3 | 60S ribosomal protein L3 | 413 | 50.36 | 26 | ↑ 10.66 ± 1.06 | ↑ 5.58 ± 2.23 | ↓ 0.51 ± 0.07 |
| A0A034WMJ8 | 40S ribosomal protein S4 | 280 | 59.29 | 23 | ↑ 10.35 ± 1.61 | ↑ 4.44 ± 2.48 a | ↓ 0.41 ± 0.1 |
| A0A034VGJ1 | Clustered mitochondria protein | 1362 | 30.54 | 35 | ↑ 9.85 ± 2.74 | ↑ 5.7 ± 3.47 | ↓ 0.53 ± 0.02 |
| A0A034V813 | Acetyl-CoA carboxylase | 2391 | 34.84 | 58 | ↑ 9.8 ± 1.2 | ↑ 5.48 ± 1.47 | ↓ 0.51 ± 0.03 |
| A0A034WGD2 | Ribosomal L1 domain-containing protein | 608 | 32.07 | 18 | ↑ 9.37 ± 0.71 | ↑ 2.86 ± 1.34 a | ↓ 0.29 ± 0.06 |
| A0A034VH20 | Piwi-like protein | 900 | 49.56 | 37 | ↑ 8.88 ± 2.99 | ↑ 3.75 ± 3.08 a | ↓ 0.35 ± 0.05 |
| A0A034VGU8 | Eukaryotic translation initiation factor 2 subunit 3 | 475 | 45.68 | 16 | ↑ 8.09 ± 1.03 | ↑ 4.25 ± 1.63 a | ↓ 0.49 ± 0.05 |
| A0A034VGS8 | Tubulin β-3 chain | 454 | 60.79 | 27 | ↑ 7.51 ± 0.22 | ↑ 3.32 ± 1.16 a | ↓ 0.42 ± 0.08 |
| A0A034V5U3 | Staphylococcal nuclease domain-containing protein 1 | 928 | 64.12 | 47 | ↑ 7.34 ± 0.89 | ↑ 2.91 ± 1.09 a | ↓ 0.35 ± 0.04 |
| A0A034VX01 | DNA-binding protein modulo | 578 | 37.89 | 22 | ↑ 7.2 ± 1.01 | ↑ 2.62 ± 0.83 a | ↓ 0.35 ± 0.01 |
| A0A034VMS2 | Peptide methionine sulfoxide reductase | 243 | 29.63 | 6 | ↑ 6.68 ± 0.34 | ↑ 3.19 ± 0.6 a | ↓ 0.45 ± 0.04 |
| A0A034VUY6 | Methenyltetrahydrofolate synthase domain-containing protein | 538 | 56.51 | 32 | ↑ 5.91 ± 0.39 | 1.09 ± 0.49 b | ↓ 0.16 ± 0.03 |
| A0A034WNF6 | Importin subunit α | 519 | 29.09 | 12 | ↑ 5.51 ± 0.44 | 1 ± 0.32 b | ↓ 0.19 ± 0.02 |
| A0A034VER3 | Eukaryotic translation initiation factor 3 subunit C | 923 | 37.92 | 32 | ↑ 5.49 ± 0.72 | ↑ 2.35 ± 1.36 a | ↓ 0.42 ± 0.09 |
| A0A034W2L3 | Citrate synthase | 257 | 63.42 | 15 | ↑ 5.37 ± 0.42 | ↑ 1.75 ± 1.24 a | ↓ 0.31 ± 0.09 |
| A0A034VH70 | La-related protein | 1206 | 24.71 | 20 | ↑ 5.31 ± 0.28 | ↑ 2.77 ± 0.4 a | ↓ 0.5 ± 0.03 |
| A0A034VA48 | Prolyl 4-hydroxylase subunit α2 | 561 | 23.53 | 9 | ↑ 5.19 ± 0.6 | 1.08 ± 0.28 b | ↓ 0.21 ± 0.01 |
| A0A034W6X9 | GTP-binding protein 128up | 368 | 52.45 | 16 | ↑ 5.12 ± 0.22 | ↑ 2.39 ± 0.93 a | ↓ 0.48 ± 0.08 |
| A0A034WIR4 | DnaJ-like protein subfamily C member 2 | 618 | 28.48 | 14 | ↑ 4.9 ± 0.62 | ↑ 2.81 ± 0.79 a | ↓ 0.56 ± 0.03 |
| A0A034WLF6 | FK506-binding protein 59 | 437 | 59.27 | 29 | ↑ 4.84 ± 0.23 | ↑ 2.14 ± 1.3 a | ↓ 0.41 ± 0.11 |
| A0A034V6Q5 | mRNA turnover protein 4-like protein | 257 | 28.79 | 8 | ↑ 4.61 ± 0.36 | ↑ 2.13 ± 0.13 a | ↓ 0.47 ± 0.04 |
| A0A034W8Z3 | Putative ATP-dependent RNA helicase DDX43 | 662 | 47.28 | 27 | ↑ 4.6 ± 0.28 | ↑ 2.5 ± 0.55 a | ↓ 0.51 ± 0.04 |
| A0A034VYR3 | Nucleolar protein 58 | 728 | 43.82 | 30 | ↑ 4.56 ± 0.83 | ↑ 1.98 ± 1.14 a | ↓ 0.41 ± 0.07 |
| A0A034WSY7 | Nucleolar GTP-binding protein | 651 | 23.04 | 13 | ↑ 3.91 ± 0.06 | 1.14 ± 0.41 b | ↓ 0.28 ± 0.05 |
| A0A034V087 | Guanine nucleotide-binding-like protein 3-like protein | 610 | 29.02 | 16 | ↑ 3.67 ± 0.47 | 0.76 ± 0.24 b | ↓ 0.2 ± 0.01 |
| A0A034V7N4 | Myosin heavy chain 95F | 1240 | 28.63 | 32 | ↑ 3.6 ± 0.57 | 1.09 ± 0.56 b | ↓ 0.3 ± 0.06 |
| A0A034VCY4 | Tudor domain-containing protein 7 | 956 | 25.63 | 18 | ↑ 3.41 ± 0.76 | 0.85 ± 0.49 b | ↓ 0.25 ± 0.05 |
| A0A034WCL6 | Peptidyl-prolyl cis-trans isomerase FKBP6 | 478 | 43.93 | 16 | ↑ 3.22 ± 0.32 | ↑ 1.53 ± 0.39 a | ↓ 0.46 ± 0.04 |
| A0A034VFT5 | tRNA (Cytosine(34)-C(5))-methyltransferase | 728 | 33.52 | 16 | ↑ 3.05 ± 0.54 | ↑ 1.78 ± 0.68 a | ↓ 0.57 ± 0.03 |
| A0A034VFV1 | Cytosolic 10-formyltetrahydrofolate dehydrogenase | 687 | 47.02 | 23 | ↑ 3.04 ± 0.23 | 1.42 ± 0.26 b | ↓ 0.48 ± 0.04 |
| A0A034VJX4 | Very long-chain fatty-acid--CoA ligase bubblegum | 666 | 30.33 | 15 | ↑ 2.95 ± 0.41 | 1.3 ± 0.44 b | ↓ 0.43 ± 0.04 |
| Protein ID | Annotation | Length | Coverage | Peptides | Ov-6/Ov-1 a,b | Ov-9/Ov-1 | Ov-9/Ov-6 |
|---|---|---|---|---|---|---|---|
| A0A034WSR6 | Defective chorion-1 protein | 723 | 28.49 | 25 | ↑ 27.31 ± 1.25 | ↑ 45.64 ± 1.03 | ↑ 2.8 ± 0.2 |
| A0A034W812 | Importin subunit α | 520 | 63.65 | 22 | ↑ 10.77 ± 0.98 | ↑ 26.6 ± 1.41 | ↑ 3.62 ± 0.43 |
| A0A034VZV8 | Chorion peroxidase | 836 | 40.31 | 29 | ↑ 12 ± 1.27 | ↑ 20.11 ± 2.27 | ↑ 2.56 ± 0.39 |
| A0A034UZ33 | Importin subunit β | 885 | 37.51 | 26 | ↑ 10.39 ± 1.97 | ↑ 16.2 ± 2.62 | ↑ 1.94 ± 0.02 |
| A0A034V3F4 | Tripeptidyl-peptidase 2 | 1353 | 47.97 | 54 | ↑ 4.9 ± 0.24 | ↑ 11.08 ± 0.67 | ↑ 2.61 ± 0.16 |
| A0A034V085 | Proteasome-associated protein ECM29-like protein | 1889 | 26.95 | 41 | ↑ 7.64 ± 2.22 | ↑ 11.06 ± 3.02 | ↑ 1.6 ± 0.04 |
| A0A034VPR3 | Elongation factor Tu GTP-binding domain-containing protein 1 | 1043 | 31.06 | 28 | ↑ 7.36 ± 0.75 | ↑ 10.71 ± 1 | ↑ 1.62 ± 0.04 |
| A0A034WID4 | Putative cation-transporting ATPase 13A1 | 1216 | 18.67 | 19 | ↑ 6.44 ± 0.93 | ↑ 10.57 ± 1.35 | ↑ 1.79 ± 0.07 |
| A0A034VC08 | Xaa-Pro dipeptidase | 480 | 42.29 | 18 | ↑ 5.88 ± 0.25 | ↑ 10.51 ± 0.34 | ↑ 2.09 ± 0.08 |
| A0A034VG52 | Adenosylhomocysteinase | 509 | 53.24 | 23 | ↑ 6.14 ± 0.98 | ↑ 9.11 ± 1.26 | ↑ 1.66 ± 0.04 |
| A0A034VV21 | Nuclear pore complex protein Nup205 | 1088 | 22.79 | 20 | ↑ 4.06 ± 0.49 | ↑ 7.84 ± 0.87 | ↑ 2.11 ± 0.04 |
| A0A034VMD8 | Cytoskeleton-associated protein 5 | 1227 | 26.57 | 26 | ↑ 3.15 ± 0.56 | ↑ 6.72 ± 1.11 | ↑ 2.26 ± 0.11 |
| A0A034WDU4 | Endoplasmin | 797 | 56.21 | 46 | ↑ 3.9 ± 0.12 | ↑ 6.47 ± 0.19 | ↑ 1.74 ± 0.03 |
| A0A034W4A7 | Glutathione S-transferase 1-1 | 207 | 49.76 | 11 | ↑ 2.74 ± 0.06 | ↑ 5.86 ± 0.51 | ↑ 2.26 ± 0.2 |
| A0A034WP50 | Chorion protein S36 | 233 | 54.08 | 10 | ↑ 8.94 ± 1.36 a | ↑ 36.76 ± 2.19 | ↑ 15.69 ± 1.6 |
| A0A034VUE5 | Myotubularin-related protein 14 | 765 | 30.59 | 18 | ↑ 8.44 ± 0.67 a | ↑ 29.22 ± 1.46 | ↑ 6.21 ± 0.44 |
| A0A034W787 | Juvenile hormone epoxide hydrolase 1 | 459 | 32.46 | 18 | ↑ 16.8 ± 1.38 a | ↑ 26.88 ± 1.96 | ↑ 2.64 ± 0.29 |
| A0A034VVU8 | Muskelin | 890 | 38.2 | 32 | ↑ 6.51 ± 0.55 a | ↑ 17.65 ± 3.35 | ↑ 4.04 ± 0.9 |
| A0A034W030 | MAP kinase-activating death domain protein | 2175 | 7.724 | 13 | ↑ 5.65 ± 0.65 a | ↑ 17.31 ± 1.03 | ↑ 3.84 ± 0.3 |
| A0A034W1D2 | UDP-glucuronosyltransferase 1-1 | 640 | 8.594 | 5 | ↑ 4.31 ± 0.54 a | ↑ 12.79 ± 1.88 | ↑ 3.44 ± 0.53 |
| A0A034W397 | Dual specificity mitogen-activated protein kinase kinase dSOR1 | 397 | 44.58 | 18 | ↑ 5.39 ± 0.1 a | ↑ 11.14 ± 0.87 | ↑ 2.67 ± 0.14 |
| A0A034WCE7 | Insulin receptor | 963 | 30.43 | 21 | ↑ 3.01 ± 0.76 a | ↑ 10.2 ± 1.62 | ↑ 4.24 ± 0.39 |
| A0A034W9D5 | Glutaminyl-peptide cyclotransferase-like protein | 330 | 15.45 | 5 | ↑ 2.57 ± 0.71 a | ↑ 9.6 ± 1.15 | ↑ 4.79 ± 1 |
| A0A034WBX7 | Ribonucleoside-diphosphate reductase | 804 | 46.27 | 34 | ↑ 2.49 ± 0.45 a | ↑ 9.34 ± 1.45 | ↑ 3.83 ± 0.28 |
| A0A034V8K5 | Aminopeptidase N | 1035 | 19.81 | 16 | ↑ 2.64 ± 0.68 a | ↑ 7.97 ± 1.66 | ↑ 3.55 ± 0.26 |
| A0A034VKU6 | Myotubularin-related protein 3 | 1279 | 17.98 | 18 | ↑ 1.86 ± 0.21 a | ↑ 7.79 ± 0.76 | ↑ 4.51 ± 0.12 |
| A0A034VIB7 | Protein phosphatase 1B | 370 | 40.54 | 13 | ↑ 3± 0.31 a | ↑ 7.73 ± 0.75 | ↑ 2.73 ± 0.23 |
| A0A034WC12 | Cytosolic endo-β-N-acetylglucosaminidase | 604 | 30.3 | 15 | ↑ 2.21 ± 0.39 a | ↑ 7.67 ± 1.22 | ↑ 3.88 ± 0.72 |
| A0A034WUY1 | Ubiquitin carboxyl-terminal hydrolase | 227 | 74.89 | 11 | ↑ 4.46 ± 0.45 a | ↑ 7.42 ± 0.68 | ↑ 1.9 ± 0.07 |
| A0A034VK02 | Dihydrolipoyl dehydrogenase | 504 | 58.33 | 26 | ↑ 3.79 ± 0.22 a | ↑ 6.88 ± 0.35 | ↑ 1.98 ± 0.15 |
| A0A034VAA7 | Centrosomin | 992 | 10.58 | 8 | ↑ 1.57 ± 0.21 a | ↑ 6.77 ± 0.68 | ↑ 4.8 ± 0.35 |
| A0A034W647 | Peptidyl-prolyl cis-trans isomerase D | 442 | 34.62 | 15 | ↑ 2.29 ± 0.07 a | ↑ 6.55 ± 0.29 | ↑ 2.98 ± 0.2 |
| G9F9Y5 | Chitinase | 483 | 24.43 | 8 | 1.18 ± 0.17 b | ↑ 6.26 ± 0.61 | ↑ 5.86 ± 0.67 |
| A0A034WDM9 | 26S protease regulatory subunit 6A | 428 | 63.55 | 25 | ↑ 3.65 ± 0.06 a | ↑ 5.63 ± 0.08 | ↑ 1.68 ± 0.06 |
| A0A034WB53 | Cullin-5 | 850 | 16.24 | 12 | 0.95 ± 0.23 b | ↑ 5.13 ± 0.53 | ↑ 5.75 ± 1.07 |
| A0A034W865 | Gamma-tubulin complex component 3-like protein | 951 | 19.77 | 15 | ↑ 1.86 ± 0.13 a | ↑ 4.98 ± 0.65 | ↑ 2.88 ± 0.36 |
| A0A034WNA4 | GMP synthase (Glutamine-hydrolyzing) | 683 | 53.44 | 30 | ↑ 1.94 ± 0.1 a | ↑ 4.92 ± 0.18 | ↑ 2.68 ± 0.09 |
| A0A034W2K6 | Major royal jelly protein 1 | 425 | 40.24 | 14 | 0.67 ± 0.05 b | ↑ 4.68 ± 0.62 | ↑ 6.5 ± 0.67 |
| A0A034WHL8 | Venom carboxylesterase-6 | 549 | 25.14 | 12 | 1.31 ± 0.07 b | ↑ 4.68 ± 0.27 | ↑ 3.84 ± 0.3 |
| A0A034WMG0 | Ubiquitin carboxyl-terminal hydrolase | 1110 | 31.08 | 29 | ↑ 2.24 ± 0.47 a | ↑ 4.28 ± 0.91 | ↑ 2.08 ± 0.18 |
| A0A034VRJ4 | Neutral α-glucosidase AB | 449 | 45.88 | 18 | 1.1 ± 0.18 b | ↑ 3.91 ± 0.57 | ↑ 3.5 ± 0.24 |
| A0A034VK16 | Cytoskeleton-associated protein 5 | 734 | 32.7 | 20 | ↑ 2.41 ± 0.18 a | ↑ 3.9 ± 0.22 | ↑ 1.69 ± 0.06 |
| A0A034W7H6 | Cyclin-dependent kinase 5-like protein | 293 | 14.68 | 4 | 1.22 ± 0.17 b | ↑ 3.82 ± 0.33 | ↑ 3.46 ± 0.48 |
| A0A034V2C6 | Serine/threonine-protein kinase Warts | 610 | 16.23 | 7 | 1.41 ± 0.06 b | ↑ 3.62 ± 0.07 | ↑ 2.66 ± 0.14 |
| A0A034W7N0 | Protein NASP-like protein | 427 | 41.22 | 15 | ↑ 1.67 ± 0.1 a | ↑ 3.4 ± 0.19 | ↑ 2.11 ± 0.12 |
| A0A034WQ10 | Cullin-4A | 841 | 27.59 | 20 | ↑ 1.79 ± 0.14 a | ↑ 3.35 ± 0.17 | ↑ 1.92 ± 0.12 |
| A0A034VQ97 | CD109 antigen | 1430 | 30.49 | 39 | 0.87 ± 0.14 b | ↑ 3.11 ± 0.21 | ↑ 3.78 ± 0.54 |
| A0A034VKF8 | Cysteine and histidine-rich protein 1-like protein | 447 | 30.2 | 10 | 1.26 ± 0.03 b | ↑ 2.88 ± 0.13 | ↑ 2.34 ± 0.12 |
| A0A034WBQ5 | α-1,4 glucan phosphorylase | 845 | 61.07 | 50 | 1.33 ± 0.11 b | ↑ 2.84 ± 0.33 | ↑ 2.2 ± 0.3 |
| A0A034VJU9 | 85/88 kDa calcium-independent phospholipase A2 | 872 | 23.28 | 19 | 1.36 ± 0.11 b | ↑ 2.8 ± 0.08 | ↑ 2.17 ± 0.2 |
| A0A034WAP1 | β-ureidopropionase | 385 | 41.3 | 11 | 0.8 ± 0.03 b | ↑ 1.98 ± 0.09 | ↑ 2.41 ± 0.12 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, D.; Li, R.; Zhang, M.-Y.; Liu, Y.-W.; Zhang, Z.; Smagghe, G.; Wang, J.-J. Comparative Proteomic Profiling Reveals Molecular Characteristics Associated with Oogenesis and Oocyte Maturation during Ovarian Development of Bactrocera dorsalis (Hendel). Int. J. Mol. Sci. 2017, 18, 1379. https://doi.org/10.3390/ijms18071379
Wei D, Li R, Zhang M-Y, Liu Y-W, Zhang Z, Smagghe G, Wang J-J. Comparative Proteomic Profiling Reveals Molecular Characteristics Associated with Oogenesis and Oocyte Maturation during Ovarian Development of Bactrocera dorsalis (Hendel). International Journal of Molecular Sciences. 2017; 18(7):1379. https://doi.org/10.3390/ijms18071379
Chicago/Turabian StyleWei, Dong, Ran Li, Meng-Yi Zhang, Yu-Wei Liu, Zheng Zhang, Guy Smagghe, and Jin-Jun Wang. 2017. "Comparative Proteomic Profiling Reveals Molecular Characteristics Associated with Oogenesis and Oocyte Maturation during Ovarian Development of Bactrocera dorsalis (Hendel)" International Journal of Molecular Sciences 18, no. 7: 1379. https://doi.org/10.3390/ijms18071379