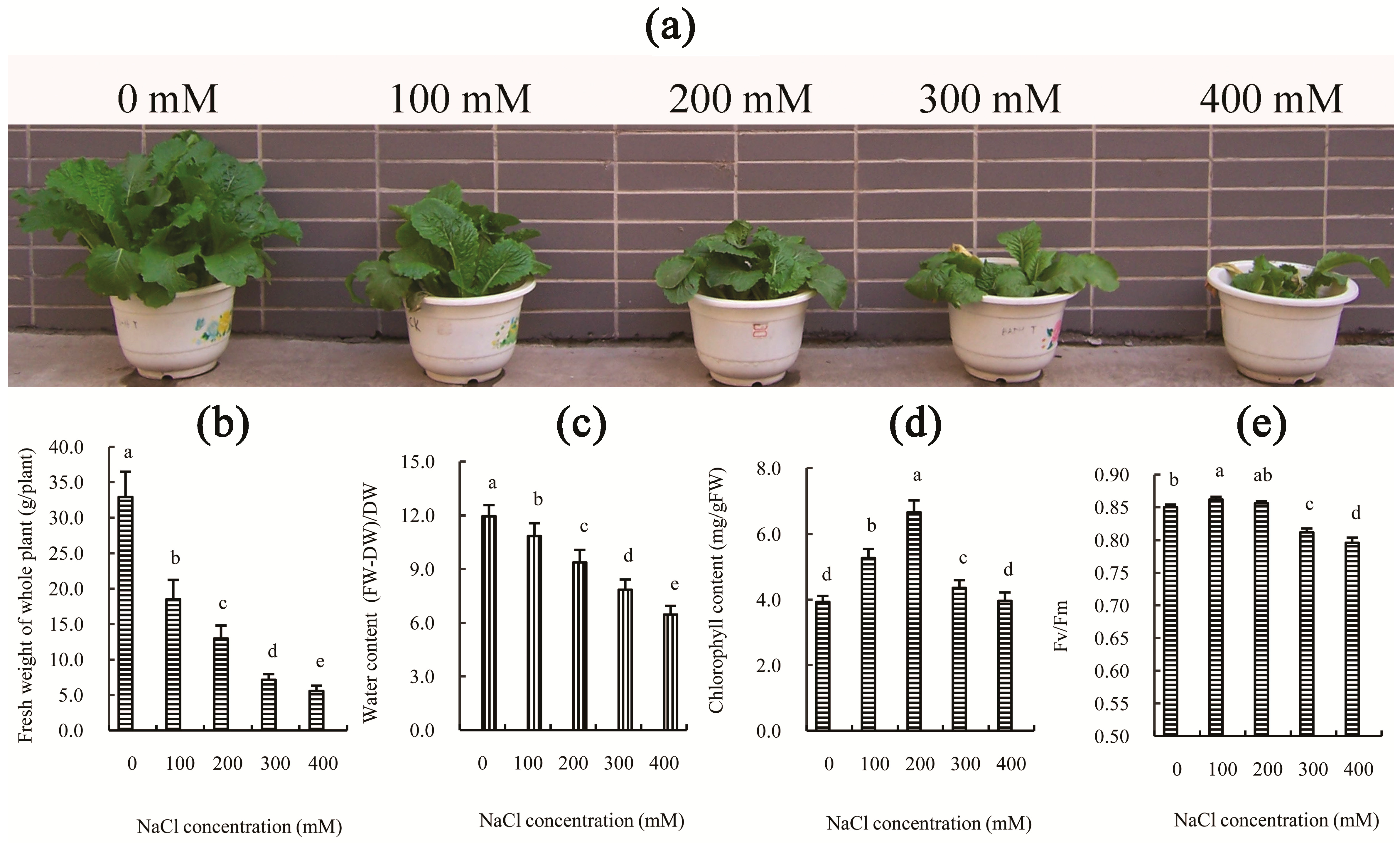
2.1. Physiological Influence of Salt Treatment on Chinese Cabbage
A series of cellular changes occur when undergoing salt stress, including water deficits, ion homeostasis, ionic toxicity, membrane alterations, and free radical production, resulting in slow growth and/or death [
2]. Previous studies have shown that treatment with different concentrations of NaCl solutions, such as 100 mM or higher, can significantly inhibit seed germination and seedling growth in Chinese cabbage [
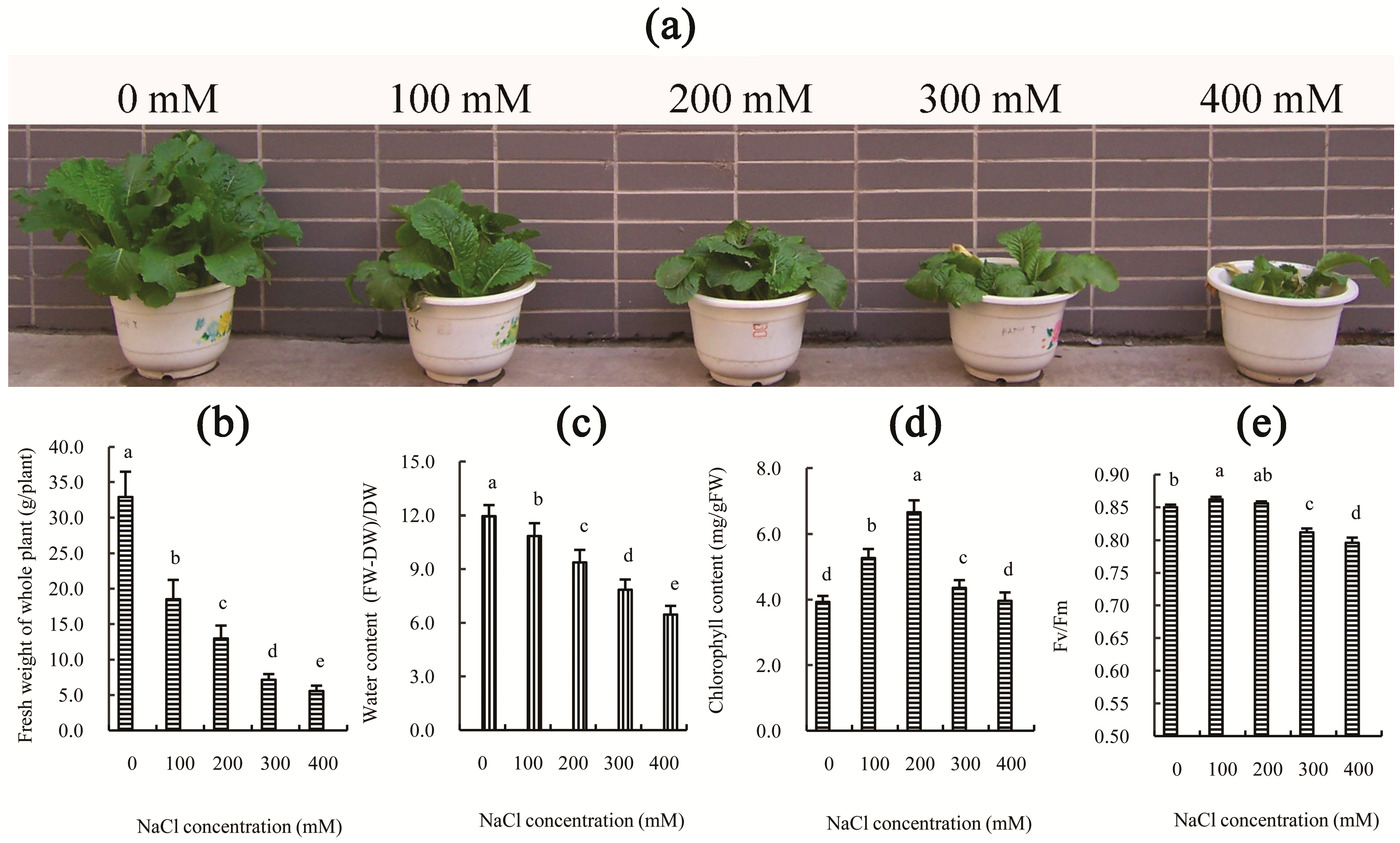
8]. In this study, the growth and the water content of Chinese cabbage were also significantly reduced by various concentrations of NaCl solution, and this effect became more pronounced following an increase in NaCl concentration at 30 days after treatment (
Figure 1a–c). However, the chlorophyll content was not reduced in the NaCl-treated leaves and even increased significantly in the 200 mM NaCl-treated leaves (
Figure 1d), although this may be partly due to inhibition of growth. Additionally, the reduction of growth of Chinese cabbage after 100 and 200 mM NaCl treatment was correlated with the decrease in photosynthetic capacity (
Table 1), rather than photosystem impairment (
Figure 1e). However, following an additional increase in NaCl concentration (300 or 400 mM NaCl), both the decline of photosynthetic capacity and the impairment of photosystems likely contributed to the inhibition of growth of Chinese cabbage. This conclusion is supported by the photosynthetic indexes (
Table 1), where the net photosynthetic rate (Pn), stomatal conductance (Gs), intercellular CO
2 concentration (Ci), and transpiration rate (Tr) all significantly decreased (
p-value < 0.05) as the concentration of salt increased. However, the maximal efficiency of PSII photochemistry (Fv/Fm) did not decrease after salt treatment for 30 days at concentrations below 300 mM (
Figure 1e). Furthermore, our results indicated that the decrease of P
n in the salt-treated leaves is primarily due to the decline of Gs and Ci. These results are consistent with our previous studies on Chinese cabbage cultivar “Juhong 65”, which can survive for long periods of time at 200 mM NaCl, but can only complete an entire life cycle at 0–100 mM NaCl [
8]. These results indicated that, as a glygophyte, Chinese cabbage has higher salt tolerance. Thus, based on the above results, 200 mM NaCl was selected as the optimal concentration for further physiological and transcriptomic analyses of Chinese cabbage.
2.2. Physiological Responses of Chinese Cabbage to Salt Stress
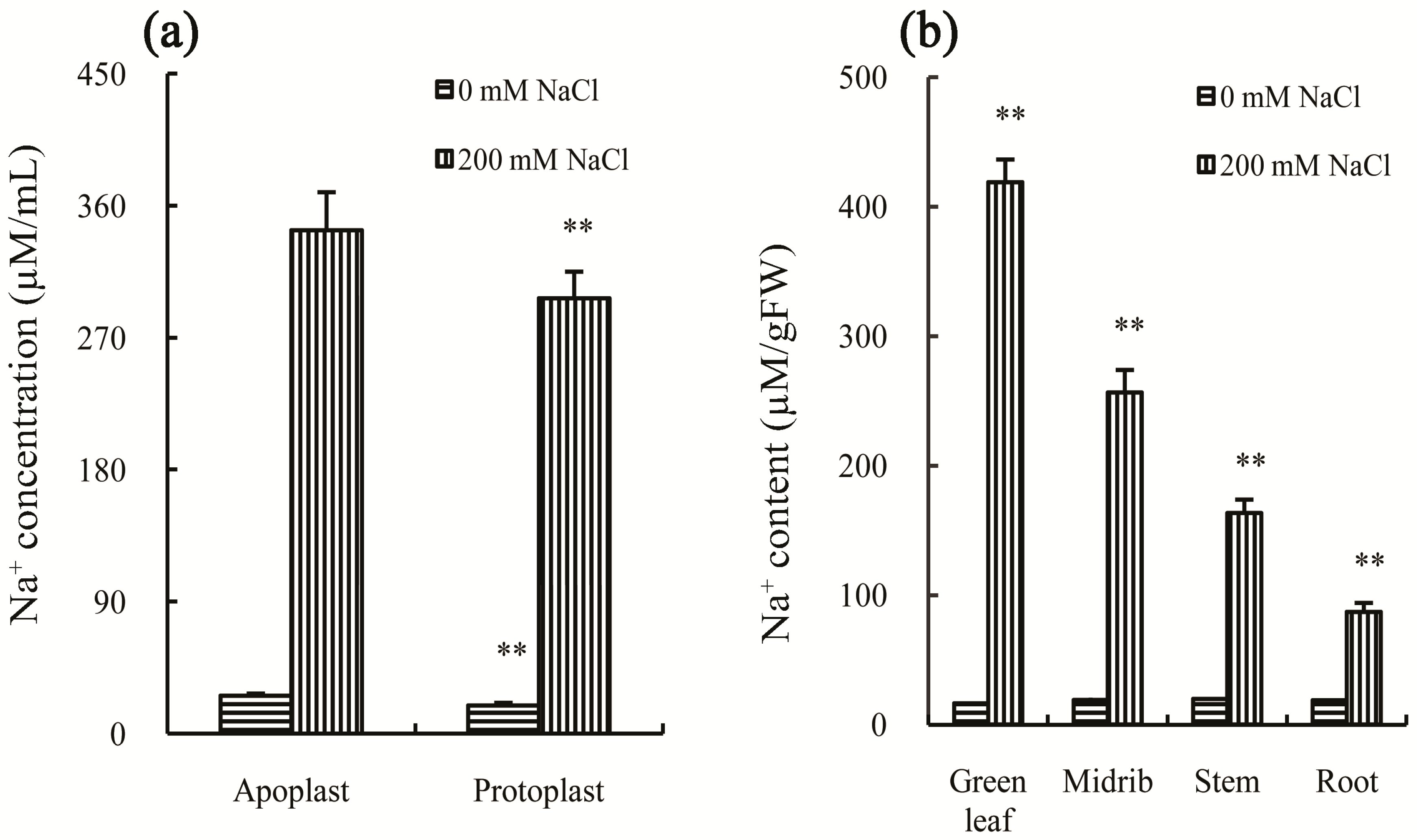
To cope with salt stress, plants have evolved many methods to reduce the damage. The common method is to quarantine the cytoplasm away from high concentrations of Na
+. There are four ways to achieve this goal, including salt excretion, salt dilution, salt accumulation, and salt exclusion [
3,
10]. However, Chinese cabbage has no salt-avoidance mechanisms with the capacity to reduce the damage caused by salt stress. Firstly, Chinese cabbage has no salt gland to secrete Na
+ out of the plant. Secondly, it cannot dilute the Na
+ by the way of more rapid growth or increased absorption of water, since its growth was significantly inhibited by salt stress, and water content decreased after salt treatment (
Figure 1a–c). Thirdly, the Na
+ content in apoplast was significantly higher (
p-value < 0.05) than that in protoplast after treatment by 200 mM NaCl for 30 days (
Figure 2a), suggesting that it cannot (unlike salt-accumulating plants) sequester the Na
+ in a vacuole as an osmoprotectant. Finally, Chinese cabbage has no capacity to prevent salt absorption or to transport it to the leaves as a salt-exclusion plant, since the leaves were found to have the highest Na
+ concentration after treatment by 200 mM NaCl for 30 days, followed by the midribs, stems, and roots (
Figure 2b). These results also confirm the proposal that Chinese cabbage belongs to the glycophyte class of plants [
8].
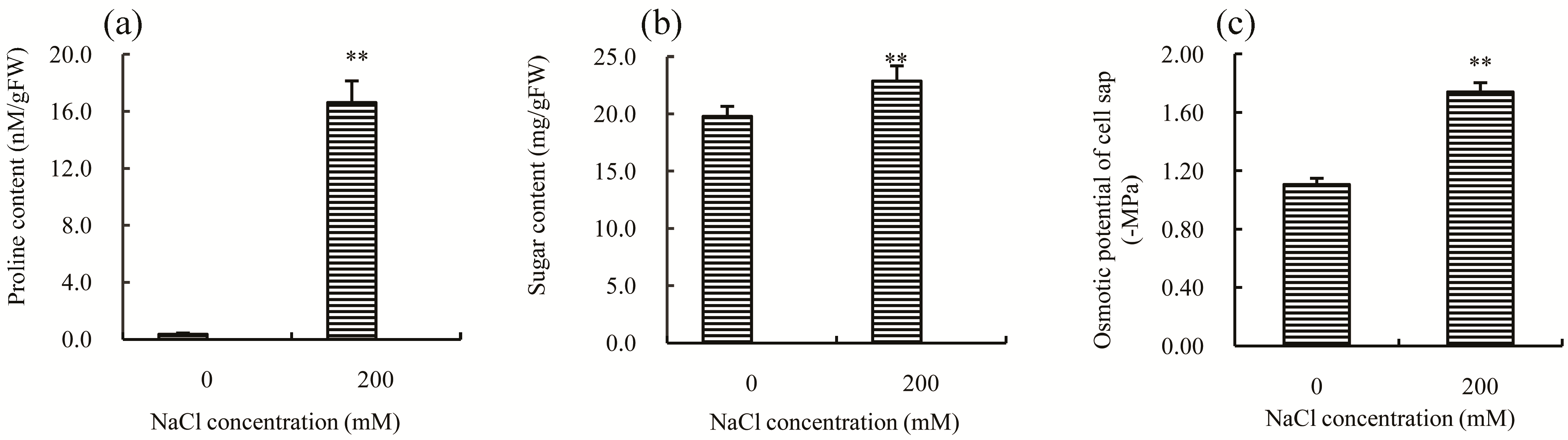
Other mechanisms that are universally employed by plants to enhance salt tolerance and to cope with salt stress include the production of osmolytes, increasing the capacity of scavenging ROS (reactive oxygen species), and maintaining the balance of Na
+/K
+ [
2,
10]. Proline and soluble sugars are two kinds of effective osmolytes that can reduce osmotic damage caused by salt stress [
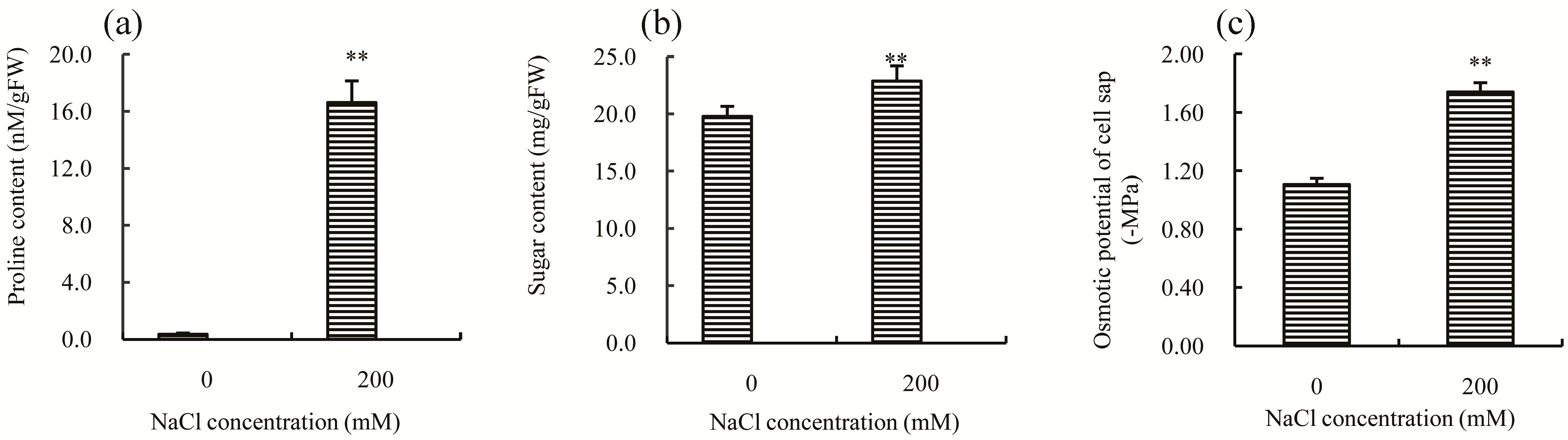
11]. After 30 days of treatment with 200 mM NaCl, both proline (
Figure 3a) and soluble sugars (
Figure 3b) significantly increased (
p-value < 0.05); notably, the concentration of proline was about 47–fold higher than the control, suggesting that this might be the principal reason for the decline observed in the osmotic potential of cell sap (
Figure 3c). Thus, modulation of proline concentration significantly contributes to increased salt tolerance in Chinese cabbage.
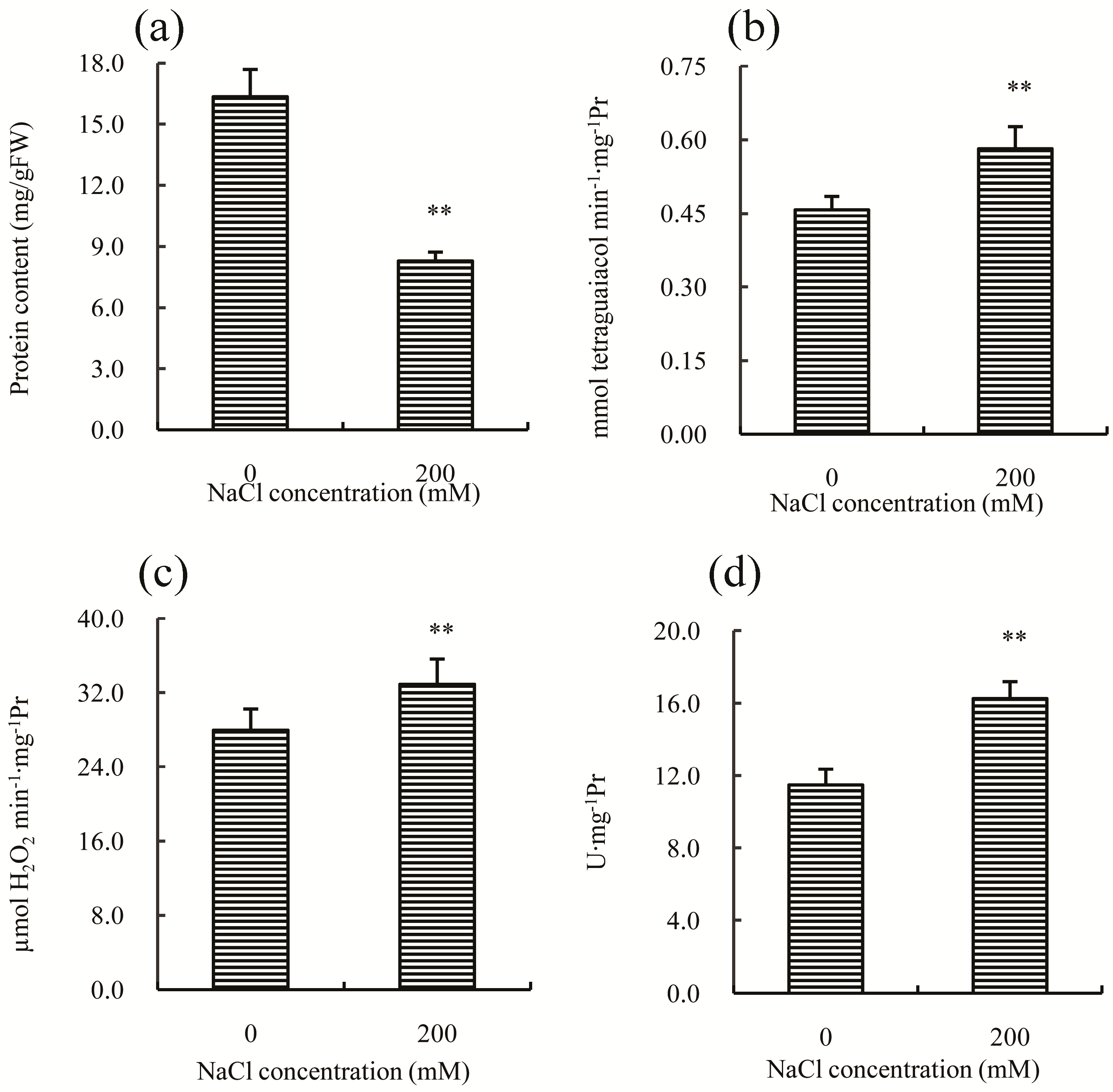
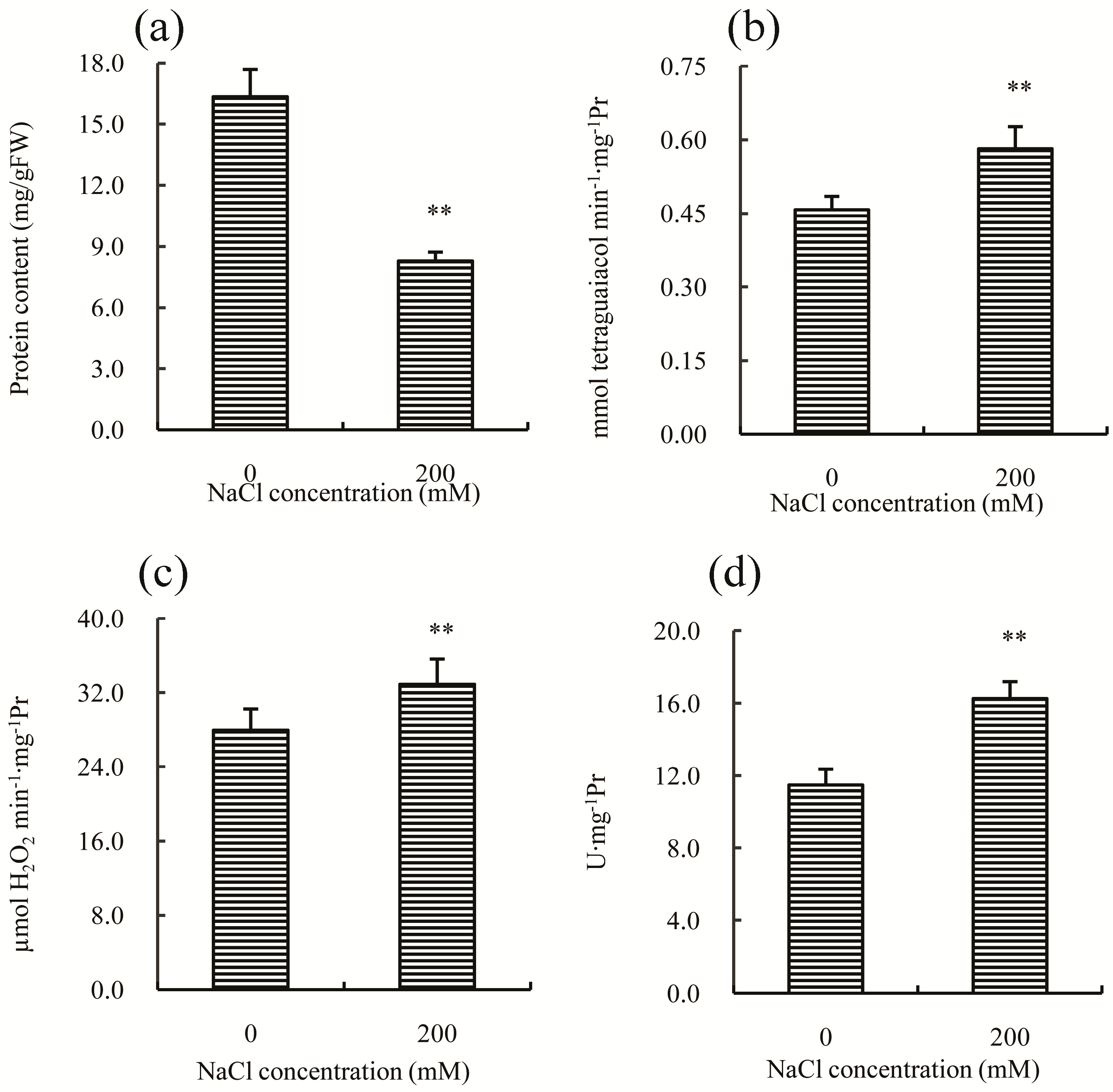
Antioxidant enzymes, such as POD (peroxidase), CAT (catalase), and SOD (superoxide dismutase), play important roles in the adaptation of plants to salt stress. In this study, the concentration of total soluble proteins was first measured, and the result indicated a significant decrease compared to the control (
Figure 4a), which may be a common response to salt stress in many plants [
12,
13,
14], due to the ability of sodium and chloride to inhibit protein synthesis [
15]. The activity of POD, CAT, and SOD increased significantly after salt treatment (
Figure 4b–d), suggesting that salt stress induced the synthesis of proteins in the antioxidant enzyme system, or that any inhibitory mechanisms in Chinese cabbage were inactivated by salt treatment. This result is consistent with previous studies on other plants, such as rice [
16], chickpea [
17] and alfalfa [
18], in which the activities of antioxidant enzymes also increase after salt treatment.
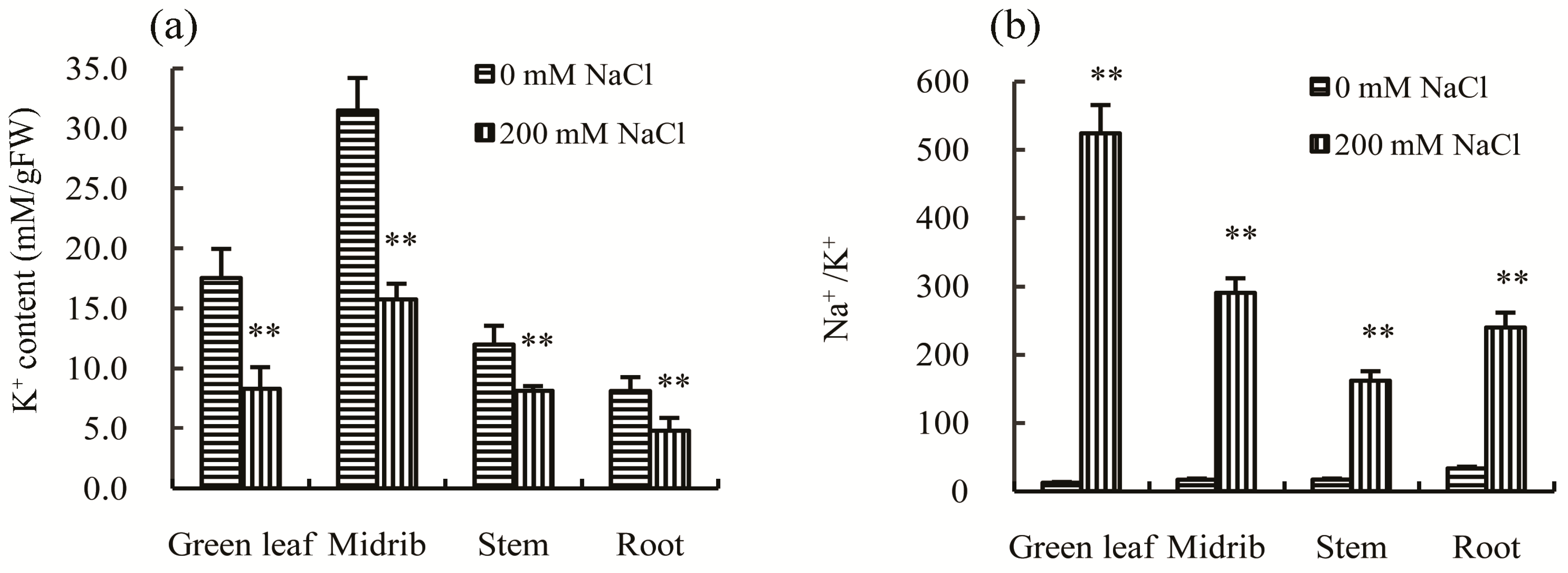
Maintaining a lower Na
+/K
+ ratio in the cytoplasm is necessary for cell function in plants. Therefore, the ability to maintain a lower Na
+/K
+ ratio in the cytoplasm of plants when under salt stress is critical. For example, the concentration of K
+ in the halophyte
Suaeda maritime increases with increased NaCl concentration, maintaining a relatively stable Na
+/K
+ ratio in the cytoplasm [
19]. However, excessive Na
+ absorbed into the cytoplasm eventually leads to competitive inhibition of K
+ uptake, resulting in a decrease in the concentration of K
+ for most plants under salt stress [
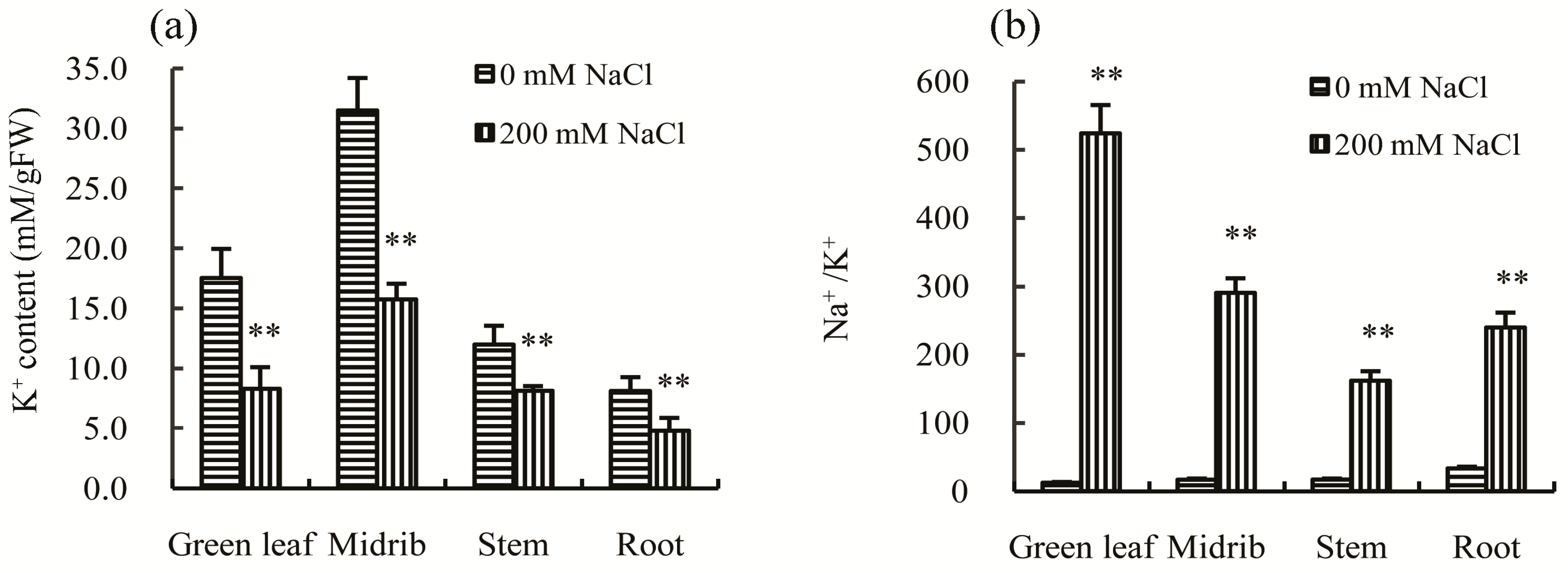
20]. In this study, the concentration of K
+ was found to be significantly lower in green leaves, midribs, stems, and roots (
Figure 5a), while the Na
+/K
+ ratios in the above organs were all significantly increased after salt treatment (
Figure 5b). These results suggest that Chinese cabbage does not have the capacity to maintain a balanced Na
+/K
+ ratio to enhance tolerance to salt stress.
Additionally, it is possible that salinity, as an important environmental stressor, could promote leaf senescence and thus affect crop yield. However, previous studies have shown that nutrients accumulated in senescent leaves can be exported to actively growing organs such as young leaves, thus allowing for fruit development during senescence [
21]. In
Figure 6a, the old leaves of a Chinese cabbage plant can be seen to be yellow and senesced, while the young leaves are still green after 30 days of treatment by 200 mM NaCl (
Figure 6a). Additionally, the senescing leaves accumulated more NaCl than the young leaves (
Figure 6b). Therefore, we propose that the course of leaf senescence is a mechanism by which the plant can resist high salinity, since this method would remove excess Na
+ from the plant by loss of the senesced leaves, allowing the plant as a whole to complete its life cycle. For example, although the Chinese cabbage is significantly inhibited by different concentrations of salt, it can still complete an entire life cycle under 100 mM NaCl treatment [
8].
2.3. Transcriptomic Responses of Chinese Cabbage to Salt Stress
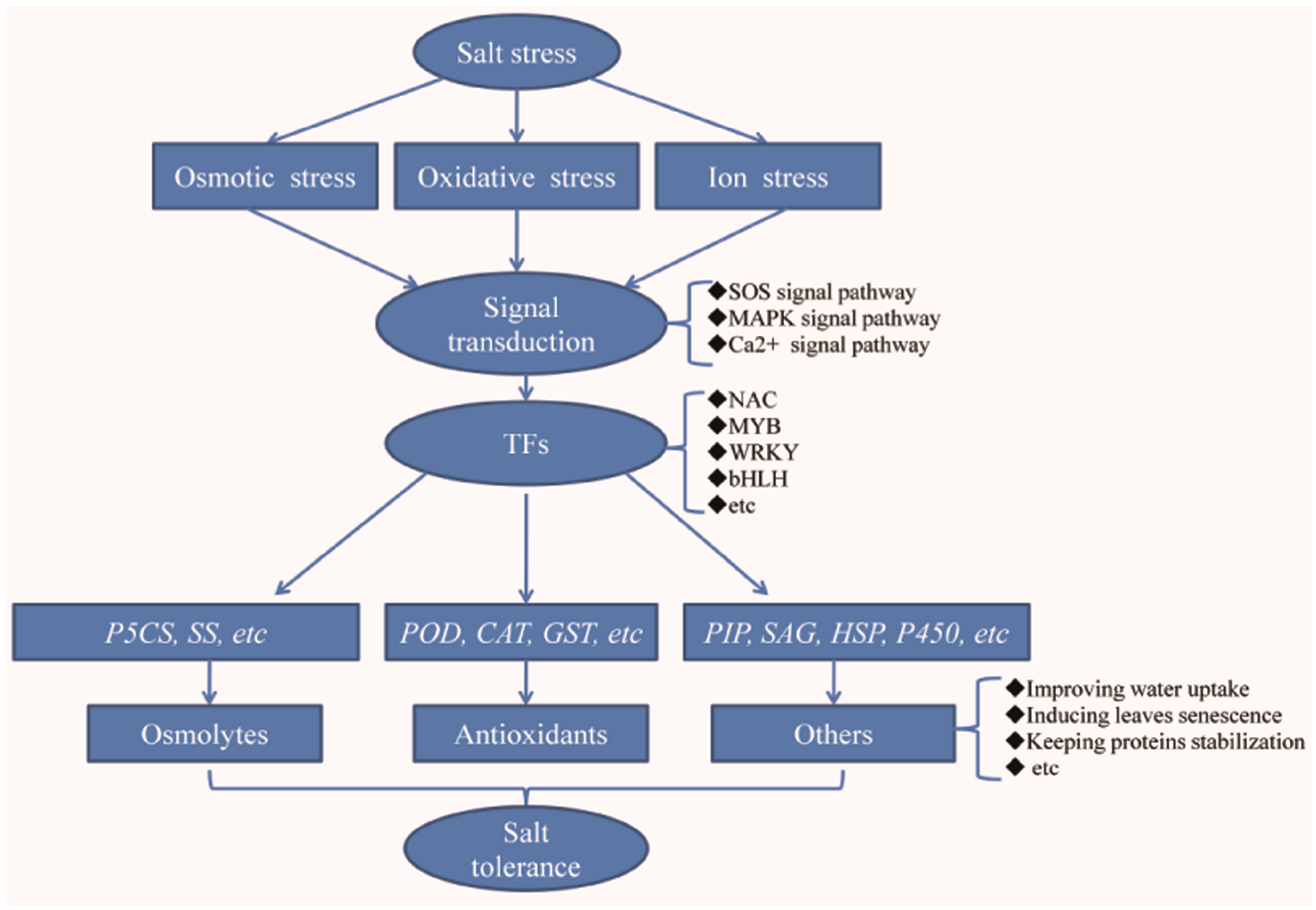
When suffering from salt stress, first the signal transduction pathways will be activated, and then a series of defense response-related genes will be induced, including those encoding TFs, proteins involved in osmolyte synthesis, antioxidant proteins, and other proteins that can confer salt tolerance on plants [
6,
7]. To explore the molecular mechanism of salt tolerance in Chinese cabbage, the RNA expression profile of both a salt-treated and a control Chinese cabbage were detected using DGE method. Additionally, to reduce false positive rates, two biological replicates each from salt-treated and control plants were sequenced, and the gene expression correlations between the two biological replicates were analyzed. The Pearson
r values were 0.987 and 0.995 for control (
Figure S1a) and salt treatment (
Figure S1b), respectively, suggesting that the data obtained from these two biological replicates were reliable. In this study, based on a threshold of divergent probability ≥0.8 and an absolute value of log2 Ratio ≥1, a total of 1235 significantly differentially expressed genes (DEGs), including 740 up-regulated and 495 down-regulated, were identified after 12 h of treatment by 200 mM NaCl (
Table S1). Additionally, to validate the DGE results, an RT-qPCR analysis was performed, and the results showed that the expression patterns of the selected 38 genes were uniform between the DGE and the RT-qPCR databases (
Table S2). Furthermore, based on their functions, the up-regulated DEGs that related to the plant defense response were classified into five subgroups, including those homologous to signal transduction proteins, TFs, proteins related to osmolyte synthesis, antioxidant proteins, and other proteins that could confer salt tolerance on plants. Of course, some genes involved in negative regulating salt tolerance were identified from the down-regulated DEGs. Additionally, the down-regulated genes that involved in salt stress response were also analyzed.
2.4. Up-Regulated Genes Involved in Signal Transduction
One important role of the SOS pathway is to activate the
SOS1 (salt overly sensitive 1) gene, which encodes a Na
+/H
+ antiporter with the function of expelling excess Na
+ from the cell, thereby maintaining cellular ion homeostasis [
22].
SOS2 encodes a CBL-interacting protein kinase that can interact with SOS3 (calcineurin B-like protein), thus activating SOS1 in the SOS pathway [
3]. In this study, five genes homologous to
SOS2 were significantly up-regulated after salt treatment (
Table 2), suggesting that they play a specific role in transducing the salt stress signal.
Calcium, a ubiquitous secondary messenger, plays an important role in the cellular response to various environmental stresses [
23,
24]. In plants, four groups of Ca
2+ sensors, including calmodulins (CaM), calmodulin-like proteins (CMLs), calcium-dependent protein kinases (CDPKs) and calcineurin B-like proteins (CBLs) are responsible for the recognition of the Ca
2+ signal [
25,
26,
27]. In this study, two
CMLs were found to be significantly up-regulated at 12 h after salt treatment (
Table 2), suggesting that they could play important roles in the salt stress response in plants. Previous studies also support this theory, as overexpression of the rice
MSR2 gene, which encodes a CML protein, enhances salt tolerance in
Arabidopsis [
28], while knockout of the
AtCML gene results in a salt-sensitive phenotype [
29].
Many studies have shown that the MAPK cascades are significant salt-stress signal transducers [
30]. The MAPK cascades are modulated by three classes of protein kinases, including MAPK kinase kinases (MAPKKKs), MAPK kinases (MAPKKs) and MAPKs [
31] Previous studies indicate that the
MAPK genes can be induced by salt treatment in a range of plant species [
32,
33,
34], and overexpression of the
MAPK genes can increase the salt tolerance of plants [
35]. Additionally, as a protein involved in the first steps of the MAPK cascade, MAPKKK can also be induced by salt stress, and plays an important role in increasing the salt tolerance of plants [
36]. Based on our data, one
MAPKKK gene was significantly induced by salt treatment (
Table 2), suggesting that MAPKKK is vital to the salt response in Chinese cabbage. However, genes homologous to
MAPK or
MAPKK were not significantly up-regulated at 12 h after salt treatment. The functions of the MAPK cascades in salt stress are subjected to further investigation.
2.7. Up-Regulated Genes Encoding Antioxidant Proteins
Salt stress leads to oxidative stress via rapid and excessive production of reactive oxygen species (ROS), such as H
2O
2, OH
● and O₂
●−, which can cause oxidative damage [
3]. As a survival mechanism, plants have evolved a set of complex antioxidant defense systems, including antioxidant enzymes, such as SOD, POD, CAT, and GST. Many studies have shown that the activity of antioxidant enzymes can be induced by salt stress [
50,
51], and overexpression of antioxidant enzymes can mitigate the damage caused by salt treatment [
52,
53,
54]. In the current study, genes homologous to POD (five genes), GST (five genes), CAT (three genes), thioredoxin superfamily protein (one gene) and ferritin 1 (one gene) were observed to be significantly up-regulated after salt treatment (
Table 5). This result was similar to previous findings in radish, where 34 genes encoding antioxidant enzymes, such as POD, SOD, and GST are significantly up-regulated by salt stress [
46]. Therefore, it can be reasonably inferred that antioxidant defense systems play a critical role in the salt stress response in Chinese cabbage.
2.8. Other Genes Induced by Salt Treatment
LEA proteins are a group of proteins that accumulate in the last stages of seed development, and play important roles in response to various stresses. For example, overexpression of
LEA genes can improve drought, salt, and freezing stress tolerance in transgenic plants [
55,
56]. LEA proteins function to improve enzyme activity and to stabilize membranes under various stress conditions [
57,
58,
59]. In this study, the expression of 19
LEA genes, including six dehydrin genes that belong to the second subgroup of
LEA genes [
60], were strongly induced by salt stress (
Table 6), suggesting that they may be involved in the salt stress response in Chinese cabbage. This result is consistent with previous findings in radish where five
LEA genes were significantly up-regulated after salt treatment [
46].
Improving the capacity of water absorption is an efficient strategy that can be used to resist osmotic stress. In plants, the transmembrane transport of water is primarily carried out by water channel proteins. Aquaporins, a class of water channel proteins, play an important role in the uptake of groundwater during stress-induced water deficits in plants [
61,
62]. The transcription level of aquaporin mRNA can be influenced by various stresses, including drought, salt, and low temperature [
63]. It has been reported that PIP (plasma membrane intrinsic protein) expression can contribute to water uptake in roots, and promote growth recovery in salinized plants [
64]. In the current study, two
PIP genes were significantly induced by salt treatment (
Table 6), suggesting that this protein may promote the transmembrane transport of water during osmotic adjustment.
Leaf senescence is a mechanism that allows Chinese cabbage to complete its life cycle under salt stress. However, the molecular mechanism is still unknown in Chinese cabbage. Previous studies have reported that the senescence-associated gene (
SAG) is involved in the regulation of leaf senescence [
65,
66]. In our study, three genes homologous to senescence-associated gene 29 were significantly induced by salt stress. These results are also consistent with the hypothesis that leaf senescence is important for salt tolerance in Chinese cabbage, and suggest that these
SAG genes may be involved in this regulatory process.
Additionally, several genes that are homologous to HSP, P450, ERD were also significantly up-regulated by salt treatment (
Table 6). These results are consistent with previous findings in other plants, including radish [
46], barley [
67], and cotton [
68]. Furthermore, several studies using protein overexpression or mutational analysis have reported that the above genes play essential roles in salt stress response [
69,
70,
71]. Taken together, these results suggest that the mechanism of salt tolerance in Chinese cabbage is very complex and is likely to be modulated by multiple genes.






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}