Identification and Antithrombotic Activity of Peptides from Blue Mussel (Mytilus edulis) Protein



Abstract
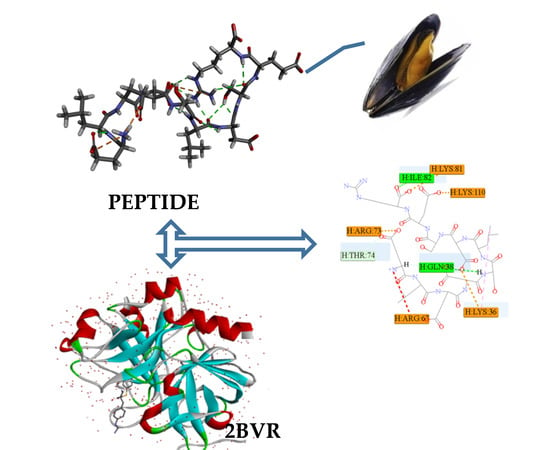
:
1. Introduction
2. Results and Discussion
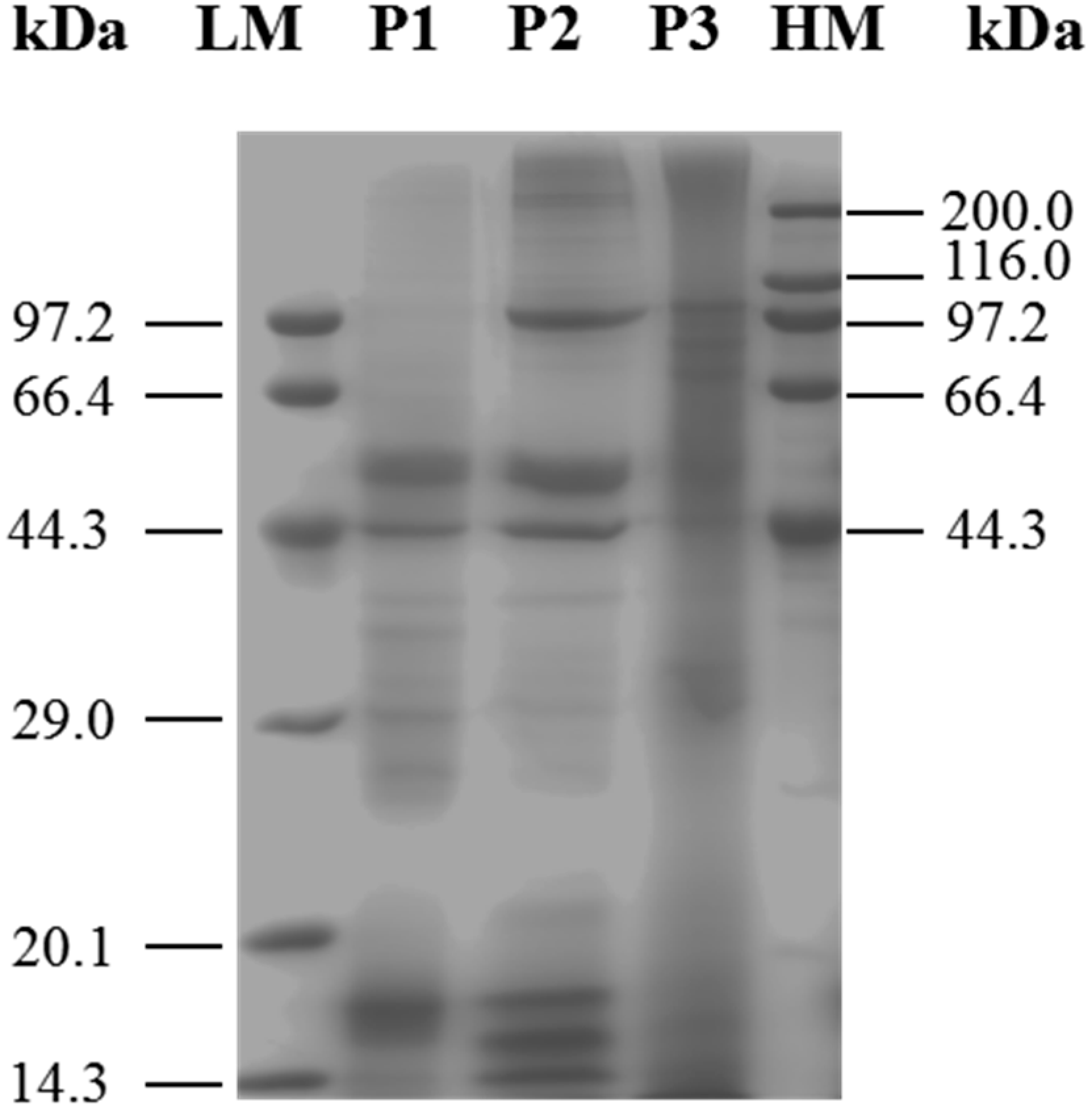
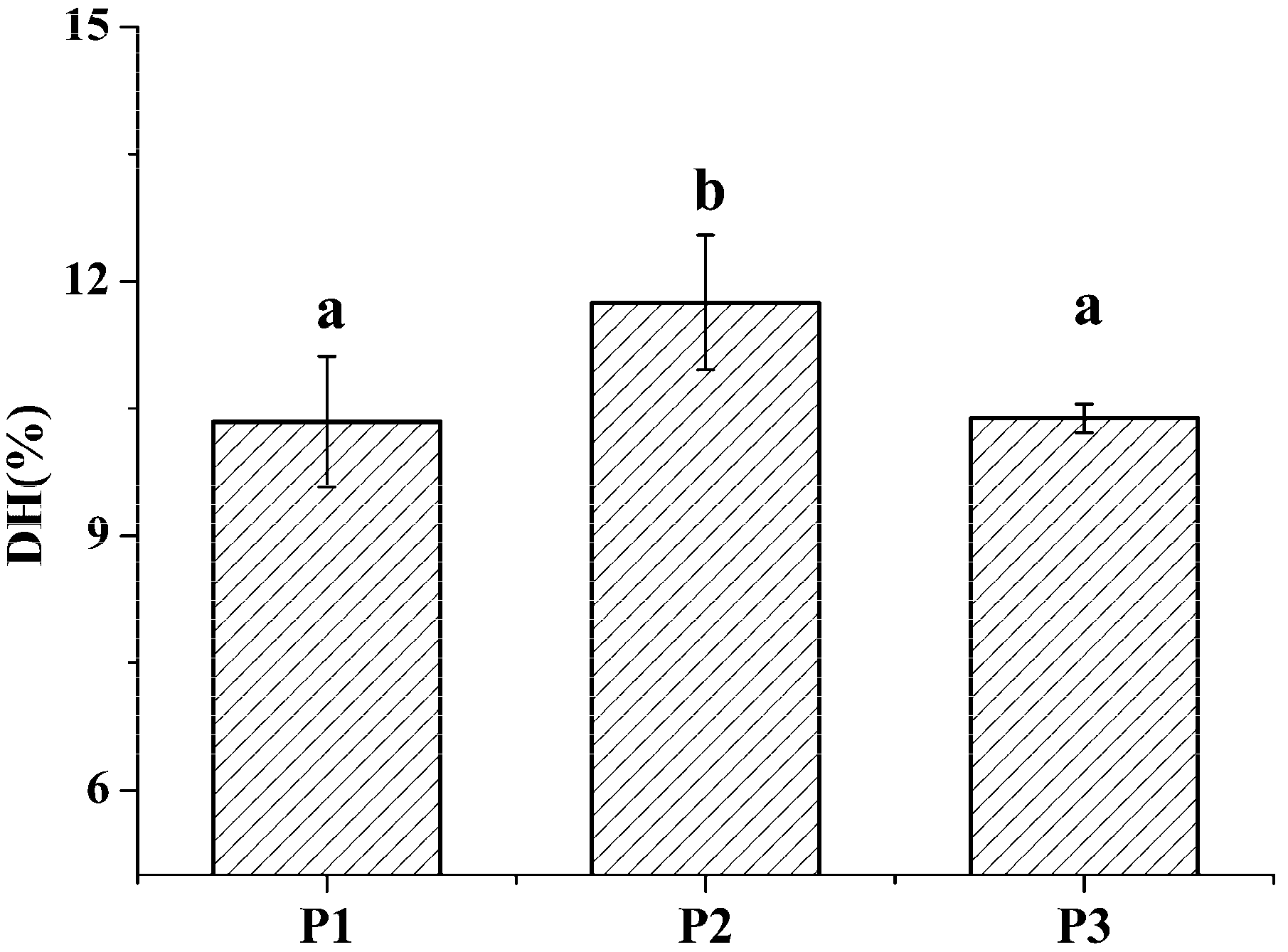
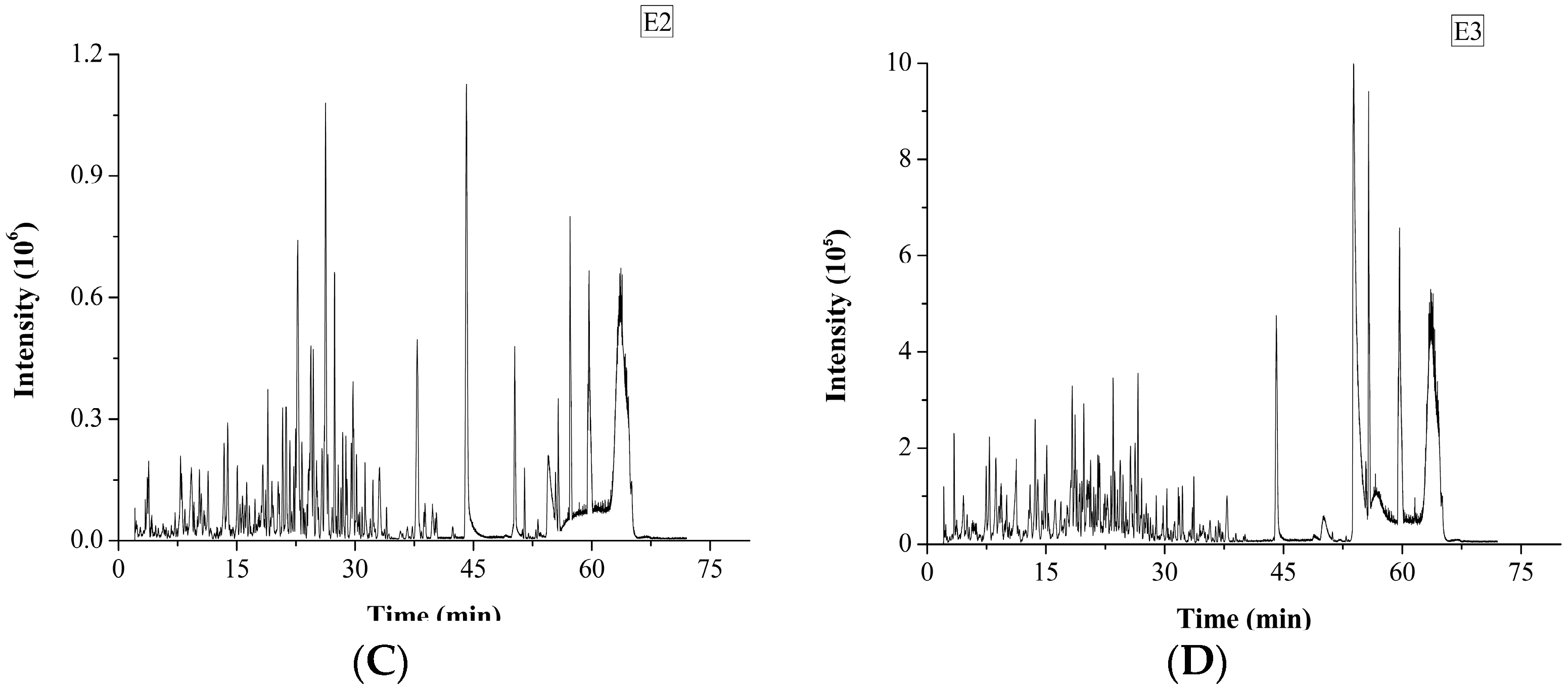
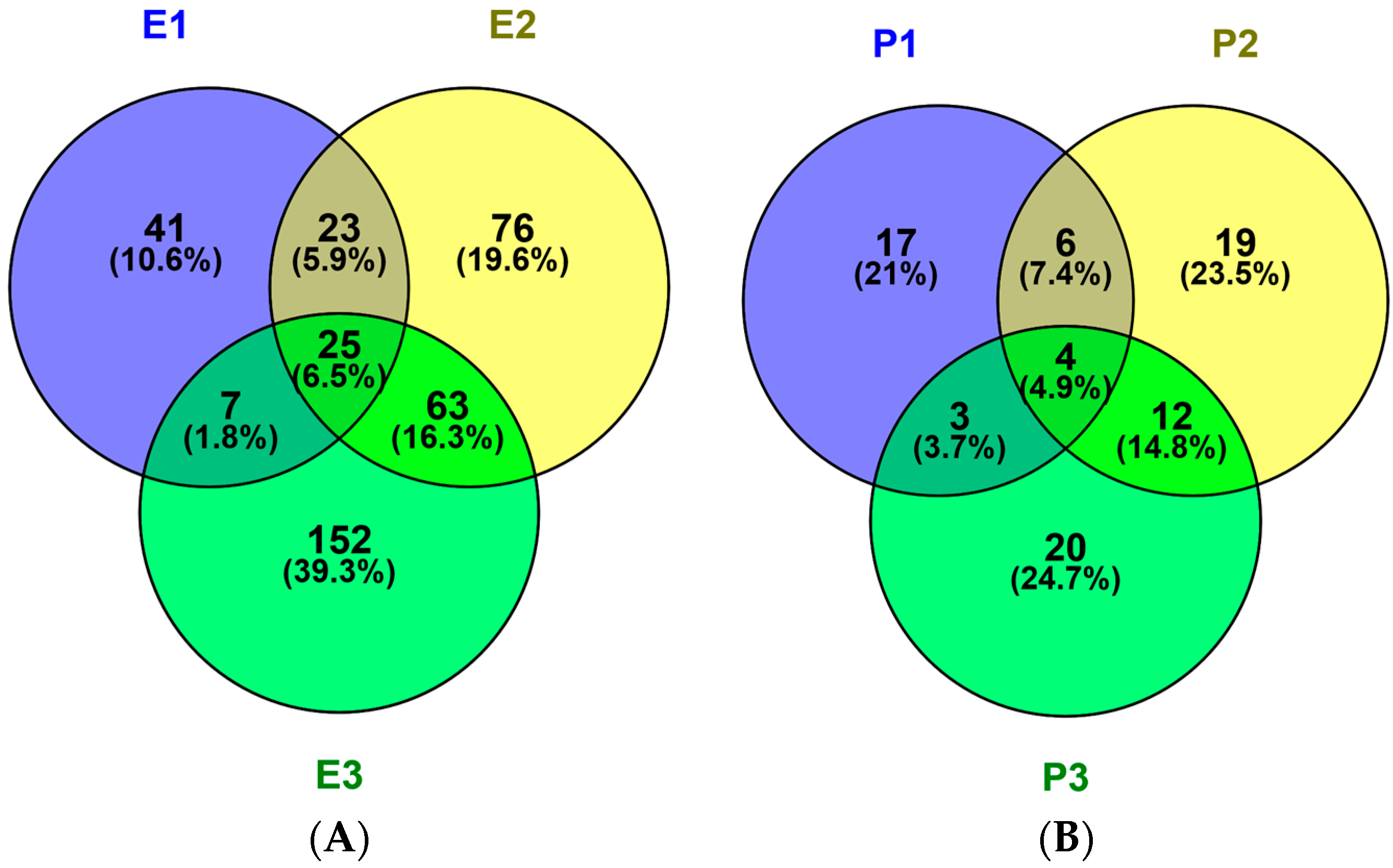
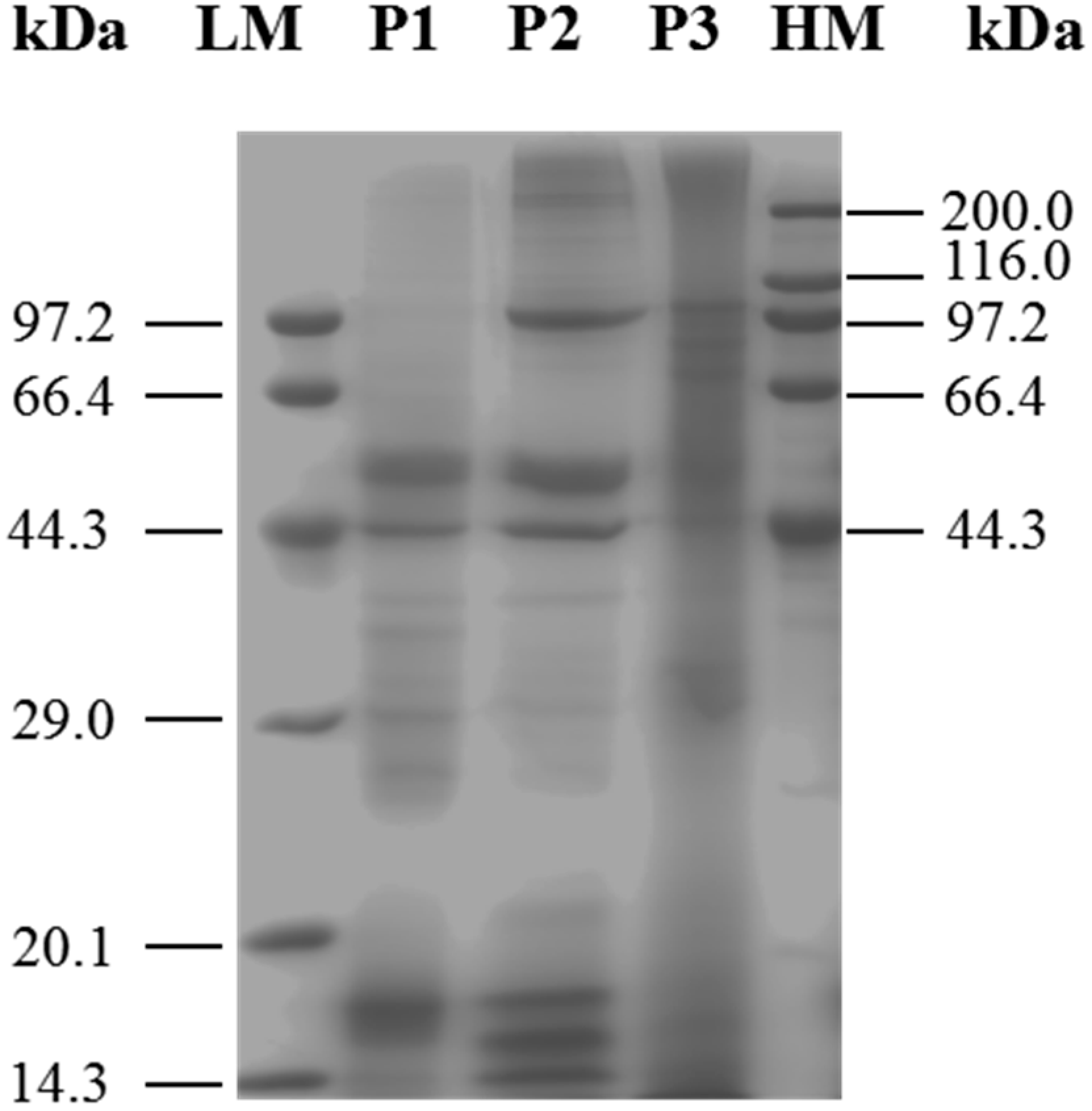
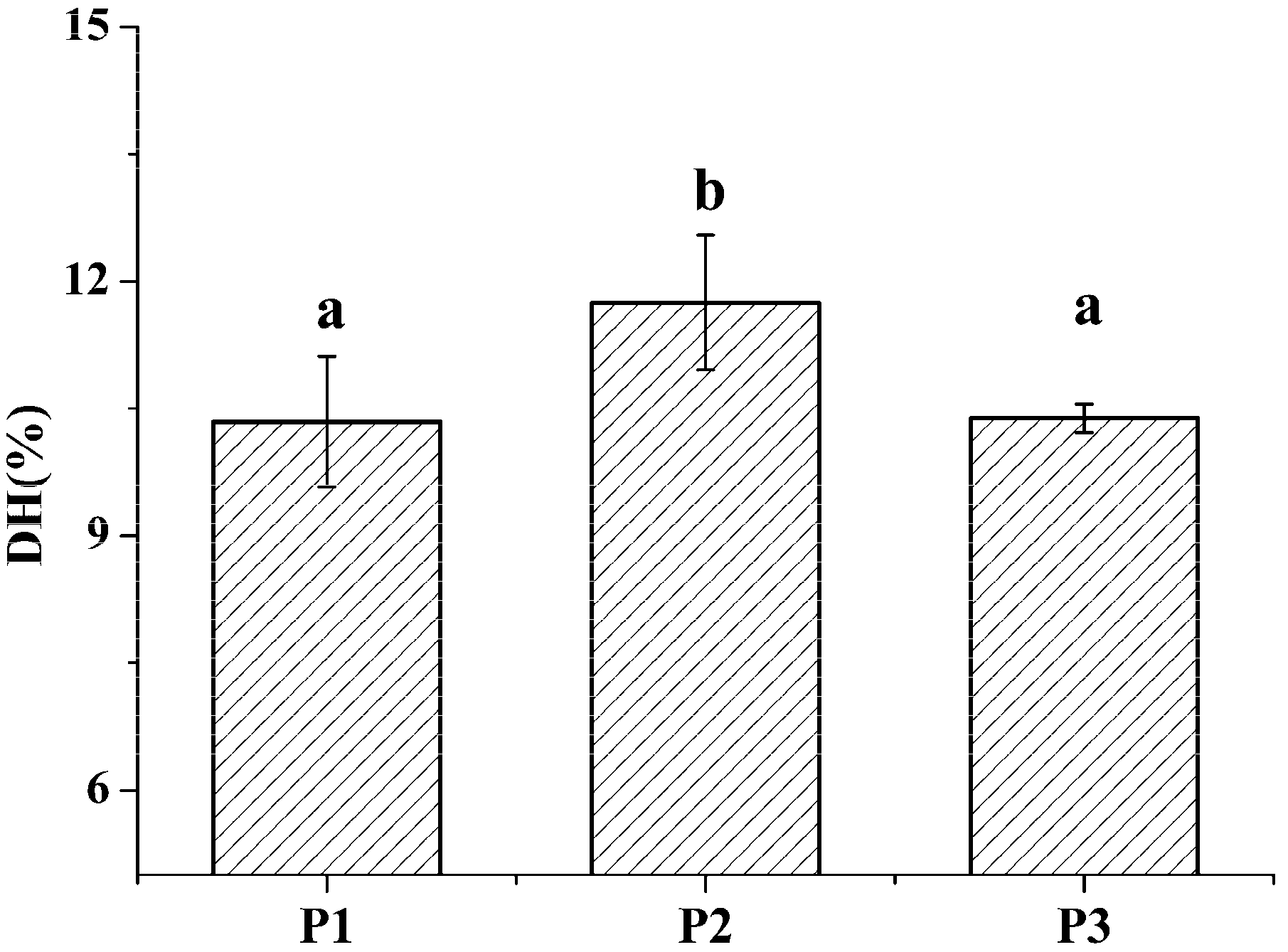
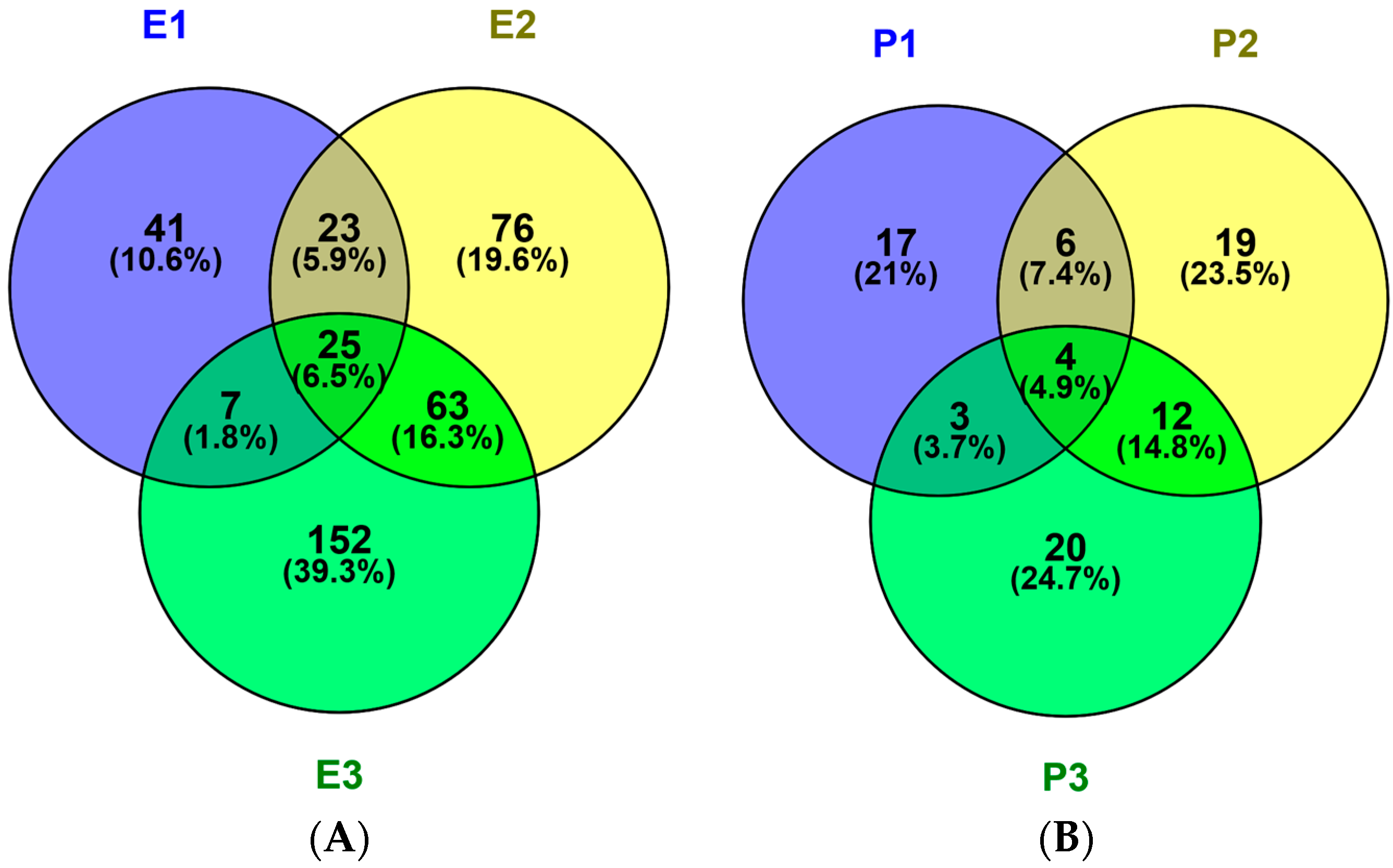
2.1. Identification and Characterization of M. edulis Proteins and Identification of Peptides by Enzymatic Hydrolysis
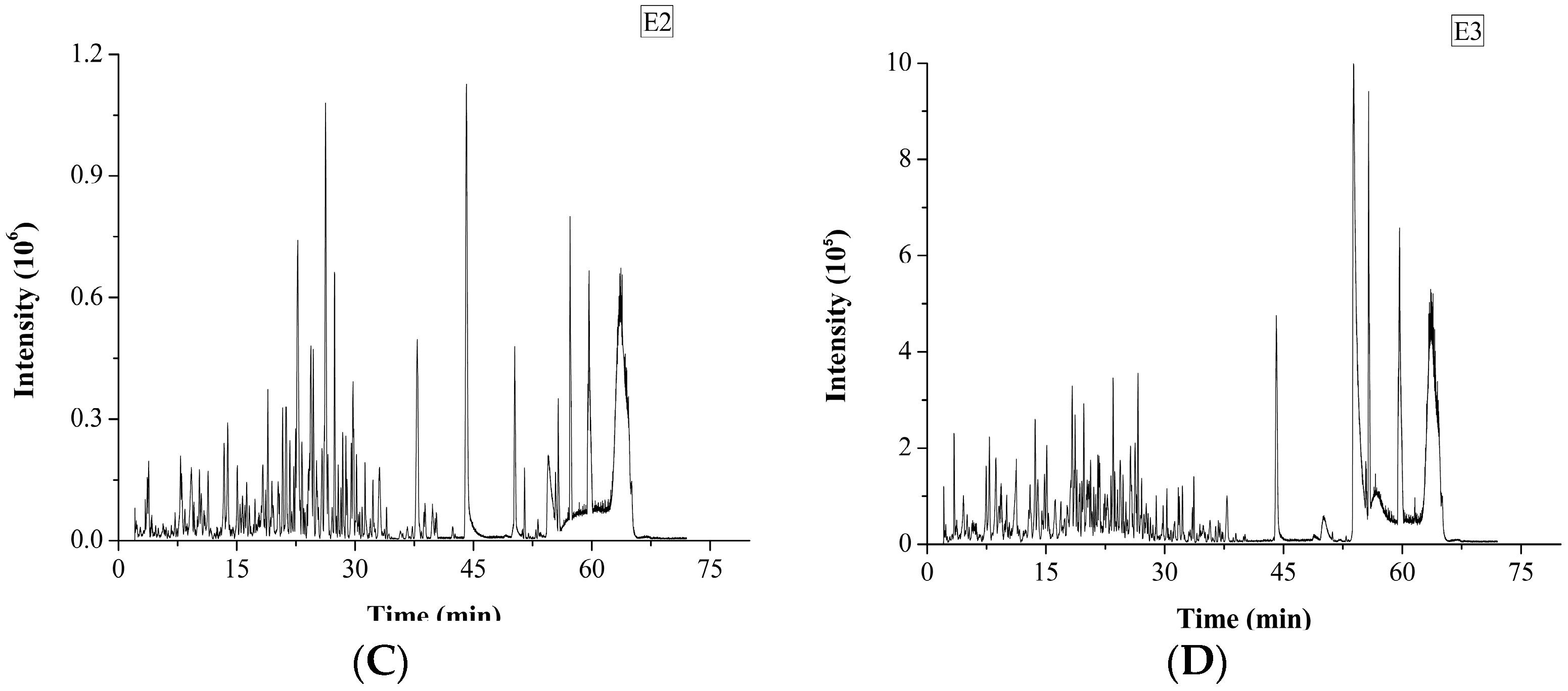
2.2. In Silico Prediction of Peptides from M. edulis
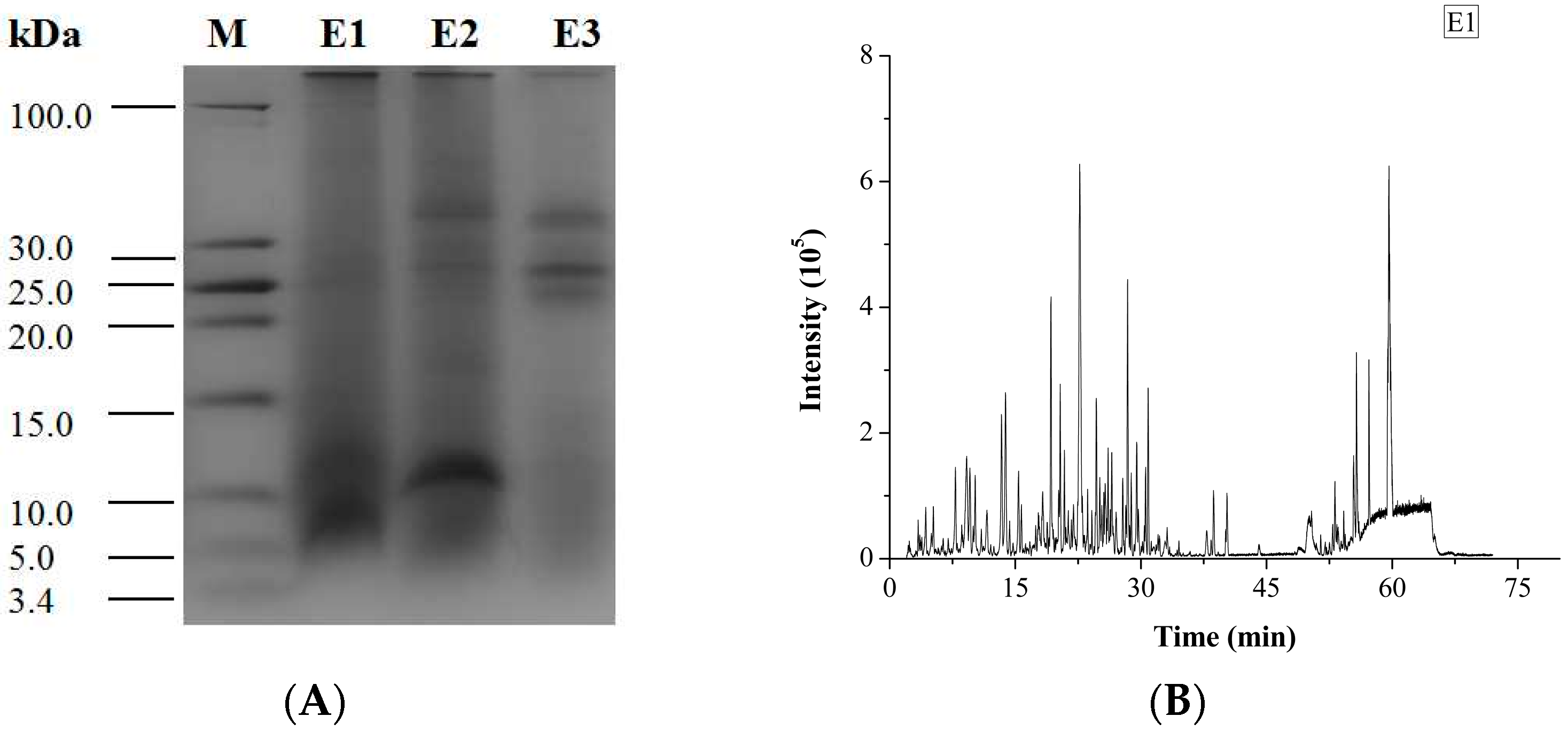
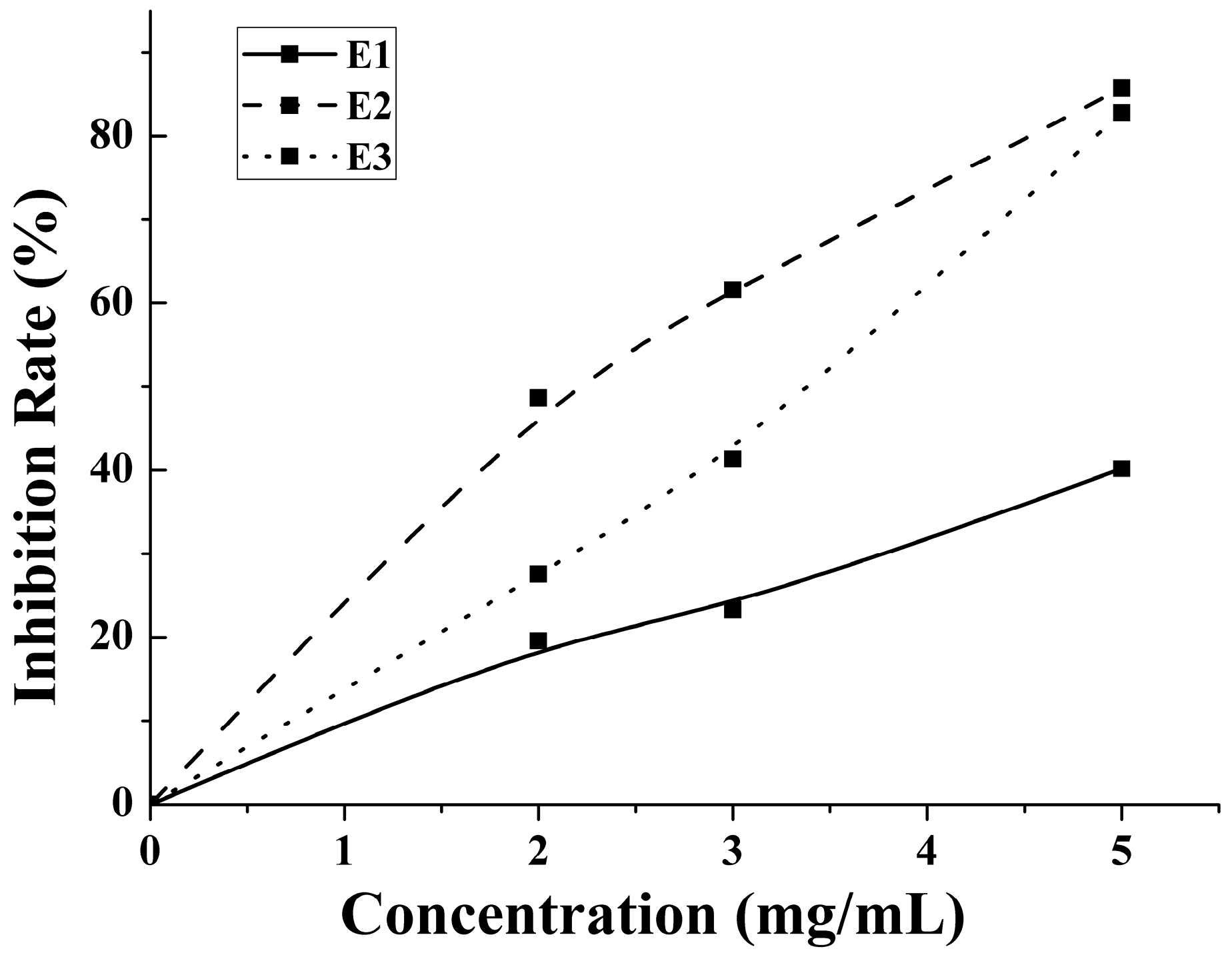
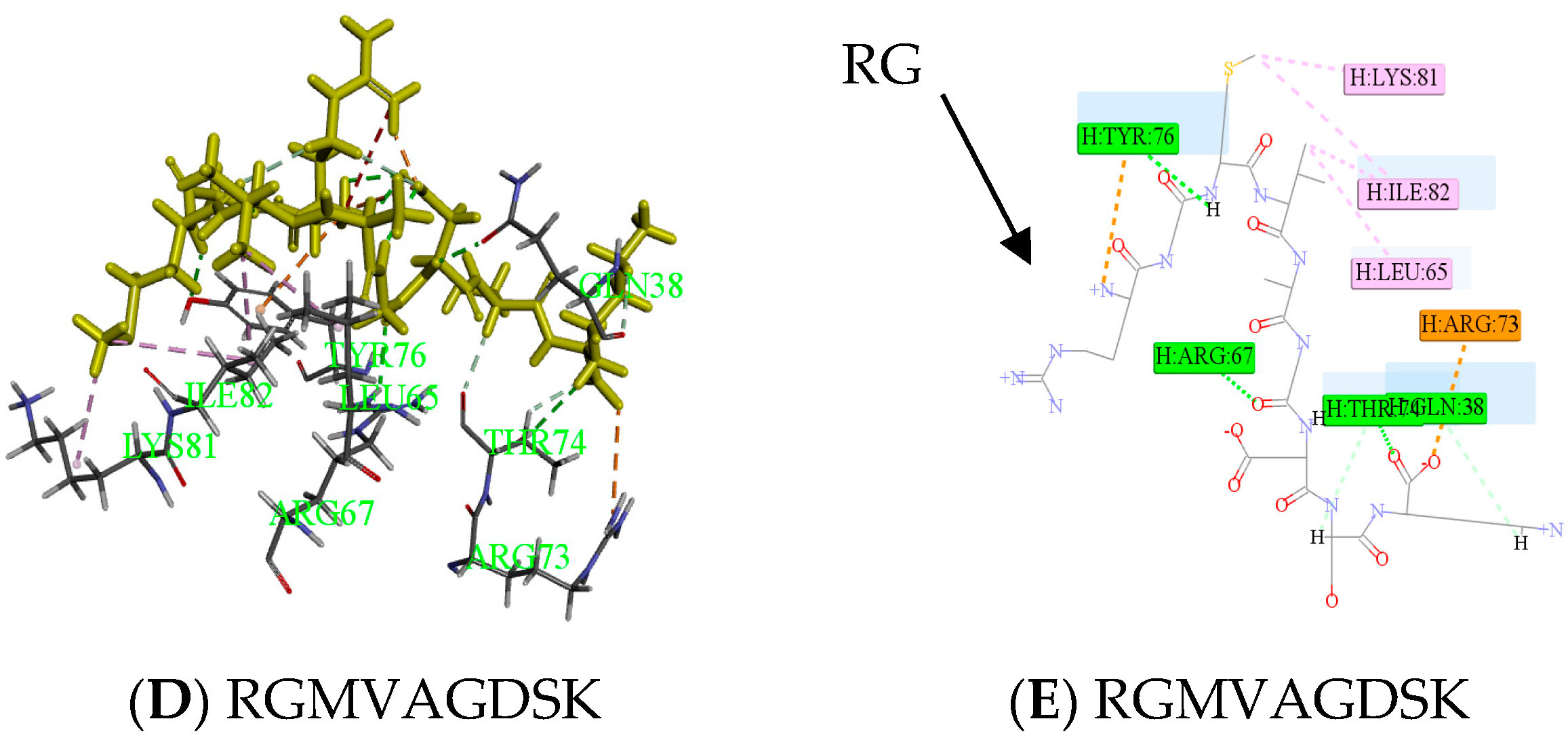
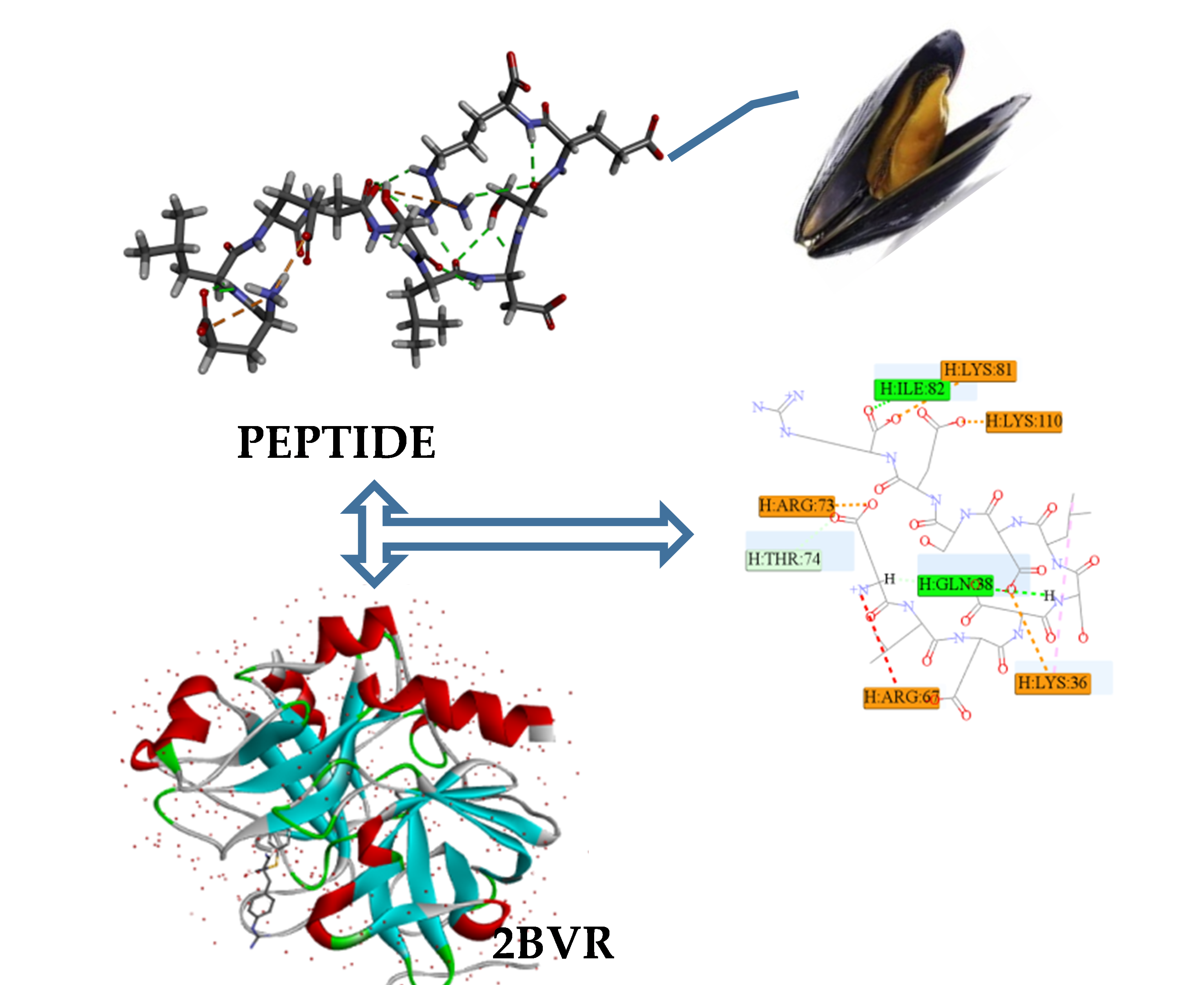
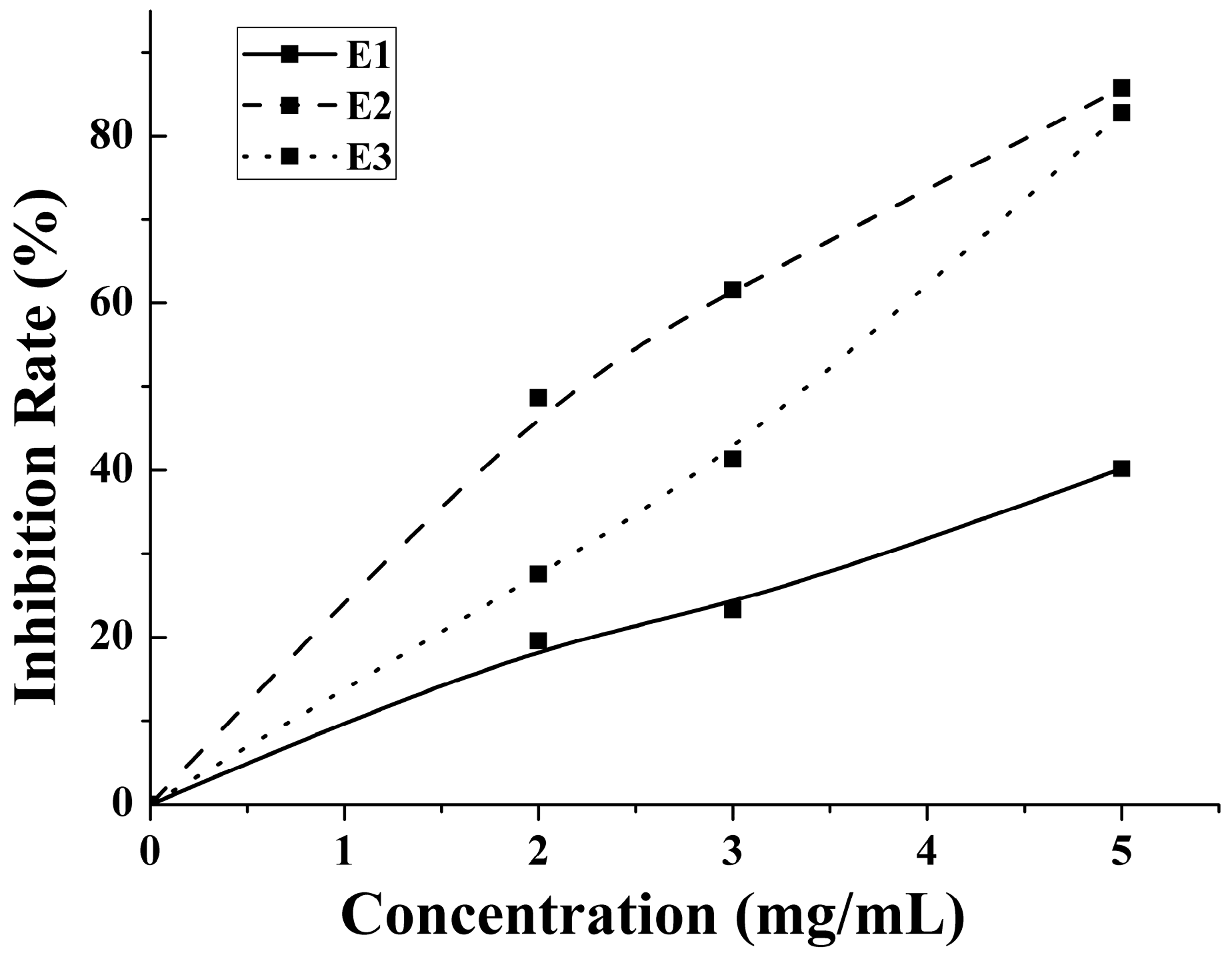
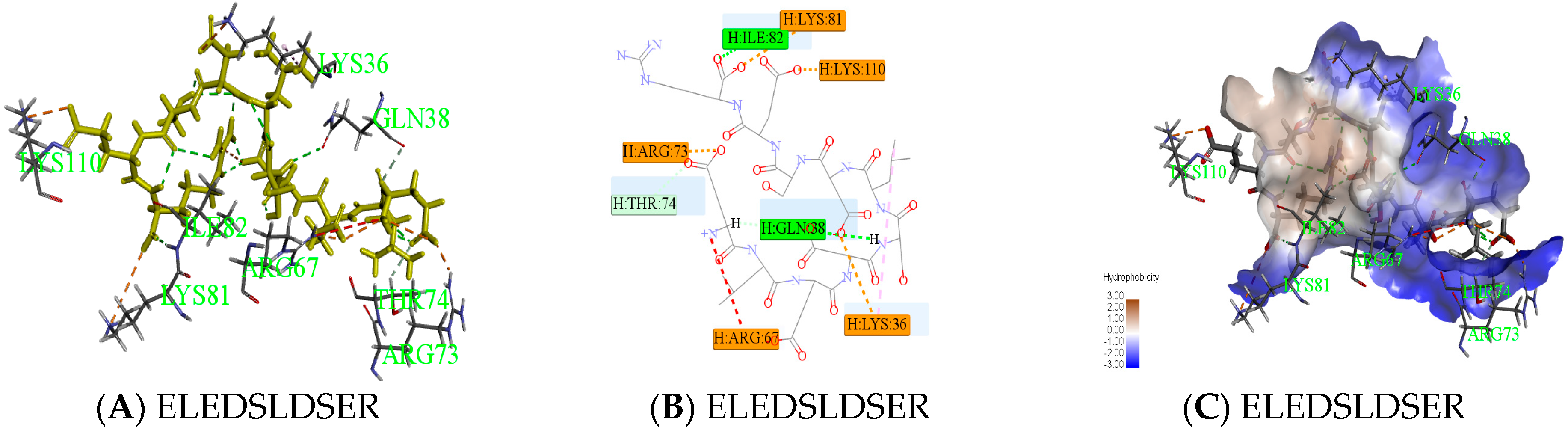
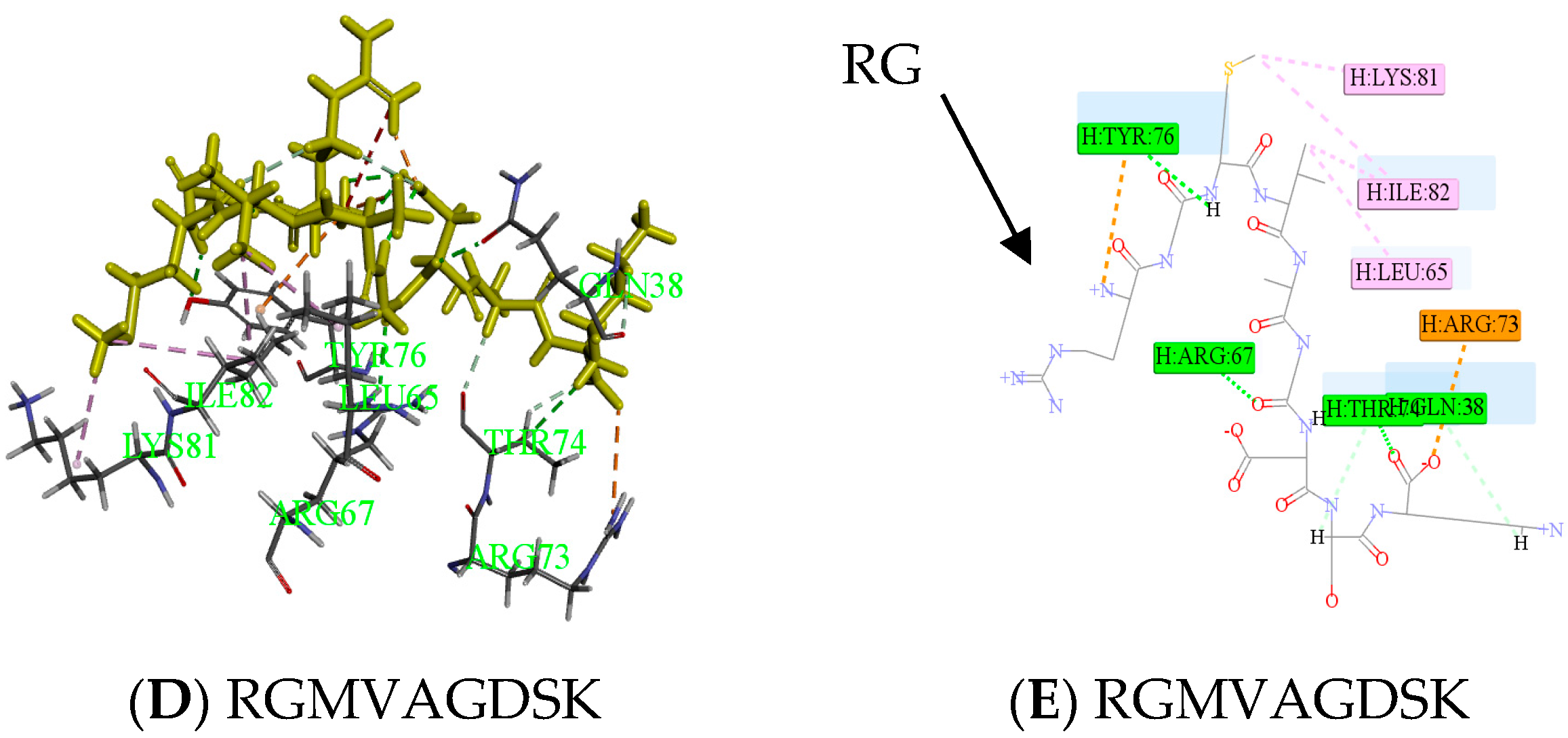
2.3. Antithrombotic Activity of E1–E3 and Active Mechanism of Antithrombotic Peptide
3. Materials and Methods
3.1. Materials and Chemicals
3.2. Preparation of M. edulis Proteins and Enzymatic Hydrolysis
3.3. Sodium Dodecyl Sulfate-Polyacryl Amide Gel Electrophoresis (SDS-PAGE)
3.4. Activation of SPE Column
3.5. Peptide Identification by UPLC-Q-TOF
3.6. In Silico Predictions of Peptide Toxicity, Allergic and Antithrombotic Activity
3.7. Determination of Thrombin Inhibitory Activity
3.8. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Field, I.A. Biology and economic value of the sea mussel Mytilus edulis. Bull. U. S. Bur. Fish. 1922, 38, 127–259. [Google Scholar]
- Kim, Y.S.; Ahn, C.B.; Je, J.Y. Anti-inflammatory action of high molecular weight Mytilus edulis hydrolysates fraction in LPS-induced RAW264.7 macrophage via NF-κB and MAPK pathways. Food Chem. 2016, 202, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Darewicz, M.; Borawska, J.; Vegarud, G.E.; Minkiewica, P.; Iwaniak, A. Angiotensin I-Converting Enzyme (ACE) Inhibitory Activity and ACE Inhibitory Peptides of Salmon (Salmosalar) Protein Hydrolysates Obtained by Human and Porcine Gastrointestinal Enzymes. Int. J. Mol. Sci. 2014, 15, 14077–14101. [Google Scholar] [CrossRef] [PubMed]
- Al-Obaidi, J.R.; Saidi, N.B.; Rokhiyah, S.; Nahadatul, S.; Yusoff, N.M.; Idris, A.S. Comparison of Different Protein Extraction Methods for Gel-Based Proteomic Analysis of Ganoderma spp. Protein J. 2016, 35, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Alu’datt, M.H.; Rababah, T.; Alhamad, M.N.; Gammoh, S.; Ereifej, K.; Alodat, M.; Hussein, N.M.; Kubow, S.; Torley, P.J. Antioxidant and antihypertensive properties of phenolic–protein complexes in extracted protein fractions from Nigella damascena and Nigella arvensis. Food Hydrocoll. 2016, 56, 84–92. [Google Scholar] [CrossRef]
- Dziuba, B.; Dziuba, M. New Milk Protein-Derived Peptides with Potential Antimicrobial Activity: An Approach Based on Bioinformatic Studies. Int. J. Mol. Sci. 2014, 15, 14531–14545. [Google Scholar] [CrossRef] [PubMed]
- Skripnikov, A.Y.; Kazakov, N.A.; Dolgovx, S.V.; Ziganshin, R.K.; Govorum, V.M.; Ivanov, V.T. The search for and identification of peptides from the moss Physcomitrella patens. Russ. J. Biol. Chem. 2011, 37, 95–104. [Google Scholar] [CrossRef]
- Stefanska, I.; PiaseckaJóźwiak, K.; Kotyrba, D.; Kolenda, M.; Stecka, K.M. Selection of lactic acid bacteria strains for the hydrolysis of allergenic proteins of wheat flour. J. Sci. Food Agric. 2016, 96, 3897–3905. [Google Scholar] [CrossRef] [PubMed]
- Gomaa, A.; Boye, J. Simultaneous detection of multi-allergens in an incurred food matrix using ELISA, multiplex flow cytometry and liquid chromatography mass spectrometry (LC–MS). Food Chem. 2015, 175, 585–592. [Google Scholar] [CrossRef] [PubMed]
- Zellal, D.; Kaddouri, H.; Grar, H.; Belarbi, H.; Kheroua, O.; Saidi, D. Allergenic changes in β-lactoglobulin induced by microwave irradiation under different pH conditions. Food. Agric. Immunol. 2007, 22, 355–363. [Google Scholar] [CrossRef]
- Feng, X.L.; Liu, Q.T.; Cao, R.B.; Zhou, B.; Zhang, Y.P.; Liu, K.; Liu, X.D.; Wei, J.C.; Li, X.F.; Chen, P.Y. Characterization and immunomodulatory function comparison of various bursal-derived peptides iolated from the humoral central immune Organ. Peptides 2012, 33, 258–264. [Google Scholar] [CrossRef] [PubMed]
- Liao, Z.; Wang, X.; Liu, H.; Fan, M.H.; Sun, J.J.; Shen, W. Molecular characterization of a novel antimicrobial peptide from Mytilus coruscus. Fish Shellish Immunol. 2013, 34, 610–616. [Google Scholar] [CrossRef] [PubMed]
- Qu, W.; Ma, H.; Jia, J.; He, R.; Luo, L.; Pan, Z. Enzymolysis kinetics and activities of ACE inhibitory peptides from wheat germ protein prepared with SFP Ultrasound-assisted processing. Ultrason. Sonochem. 2012, 19, 1021–1026. [Google Scholar] [CrossRef] [PubMed]
- Kleekayai, T.; Harnedy, P.A.; O’Keeffe, M.B.; Poyarkov, A.A.; Cunhaneves, A.; Suntornsuk, A.X.; FitzGerald, R.J. Extraction of antioxidant and ACE inhibitory peptides from pthai traditional fermented shrimp pastes. Food Chem. 2015, 176, 441–447. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.X.; Wu, H.; Ling, Y.F.; Lu, R.R. Isolation and identification of antioxidant peptides derived from whey protein enzymatic hydrolysate by consecutive chromatography and Q-TOF MS. J. Dairy Res. 2013, 80, 367–373. [Google Scholar] [CrossRef] [PubMed]
- Koyama, M.; Naramoto, K.; Nakajima, T.; Aoyama, T.; Watanabe, M.; Nakamura, K. Purification and identification of antihypertensive peptides from fermented buckwheat sprouts. J. Agric. Food Chem. 2013, 61, 3013–3021. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Wu, H.; Lai, F.R.; Yang, M.Y.; Li, X.F.; Tang, Y.Q. Isolation and identification of a novel anticoagulant peptide from enzymatic hydrolysates of scorpion (Buthusmartensii Karsch) protein. Food Res. Int. 2014, 64, 931–938. [Google Scholar] [CrossRef]
- Ren, Y.; Yang, Y.J.; Wu, W.J.; Zhang, M.M.; Wu, H.; Li, X.F. Identification and characterization of novel anticoagulant peptide with thrombolytic effect and nutrient oligopeptides with high branched chain amino acid from Whitmaniapigra protein. Amino Acids 2016, 48, 2657–2670. [Google Scholar] [CrossRef] [PubMed]
- Neumann, T.; Junker, H.D.; Keil, O.; Burkert, K.; Ottleben, H.; Gamer, J.; Sekul, R.; Deppr, H.; Feurer, A.; Tomandl, D.; et al. Discovery of thrombin inhibitor fragments from chemical microarray screening. Lett. Drug Des. Discov. 2005, 2, 590–594. [Google Scholar] [CrossRef]
- Bode, W. Structure and interaction modes of thrombin. Blood Cells Mol. Dis. 2006, 36, 122–130. [Google Scholar] [CrossRef] [PubMed]
- Kjeldahl, J. Neuemethodezurbestimmung des stickstoffs in organischenKörpern (New method for the determination of nitrogen in organic substances). Z. Anal. Chem. 1883, 22, 366–383. [Google Scholar] [CrossRef]
- Lynch, J.M.; Barbano, D.M. Kjeldahlnitrogen analysis as a reference method for protein determination in dairy products. J. AOAC Int. 1999, 82, 1389–1398. [Google Scholar] [PubMed]
- Khaket, T.P.; Singh, J. Potential of plant’s dipeptidyl peptidase I & II homologs in generation of ace inhibitory peptides. Int. J. Pept. Res. Ther. 2016, 1, 81–90. [Google Scholar]
- Hörnaeus, K.; Guillemant, J.; Mi, J.; Hernroth, B.; Bergquist, J.; Bergstrom, L. Mass spectrometry data from a quantitative analysis of protein expression in gills of immuno-challenged blue mussels (Mytilus edulis). Data Brief 2016, 8, 470–473. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.M.; Zhu, B.W.; Wu, H.T.; Yu, L.; Zhou, D.Y.; Dong, X.; Yang, J.-F.; Li, D.-M.; Ye, W.-X.; Murata, Y. Purification and characterization of cathepsin B from the gut of the sea cucumber (Stichopus japonicas). Food Sci. Biotechnol. 2011, 20, 919–925. [Google Scholar] [CrossRef]
- Marbaix, H.; Budinger, D.; Dieu, M.; Fumiere, O.; Gillard, N.; Delahaut, P.; Mauro, S.; Raes, M. Identification of proteins and peptide biomarkers for detecting banned processed animal proteins (PAPs) in meat and bone meal by mass spectrometry. J. Agric. Food Chem. 2016, 64, 2405–2414. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, J.; Li, W.; Ullah, K.; Hasan, M.; Linna, G.; Awan, U.; Zhang, Y.Q.; Batool, S.; Qing, H.; Deng, Y.L. Study of rat hypothalamic proteome by HPLC/ESI ion trap and HPLC/ESI-Q-TOF MS. Proteomics 2013, 13, 2455–2468. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Zheng, W.; Li, J.; Wang, L.C.; Wu, H.; Wang, X.Z.; Shi, L. Rapid identification of bioactive peptides with antioxidant activity from the enzymatic hydrolysate of mactraveneriformis by UHPLC-Q-TOF mass spectrometry. Food Chem. 2015, 167, 484–499. [Google Scholar] [CrossRef] [PubMed]
- Bougatef, A.; Balti, R.; Nedjar-Arroume, N.; Ravallec, R.; Adje, E.Y.; Souissi, N.; Lassoued, I.; Guillochon, D.; Nasri, M. Evaluation of Angiotensin I-converting enzyme (ACE) inhibitory activities of smooth hound (Mustelusmustelus) muscle protein hydrolysates generated by gastrointestinal proteases: Identification of the most potent active peptide. Eur. Food Res. Technol. 2010, 231, 127–135. [Google Scholar] [CrossRef]
- Aivaliotis, M.; Gevaert, K.; Falb, M.; Tebbe, A.; Konstantinidis, K.; Bisle, B.; Klein, C.; Martens, L.; Staes, A.; Timmerman, E.; et al. Large-scale identification of N-terminal peptides in the Halophilic Archaea Halobacterium salinarum and Natronomonas pharaonis. J. Proteome Res. 2007, 6, 2195–2204. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Kapoor, P.; Chaudhary, K.; Gautam, A.; Kumar, R. In Silico Approach for predicting toxicity of peptides and proteins. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [PubMed]
- Lafarga, T.; Wilm, M.; Wynne, K.; Hayes, W. Bioactive hydrolysates from bovine blood globulins: Generation, characterisation, and in silico prediction of toxicity and allergenicity. J. Funct. Foods 2016, 24, 142–155. [Google Scholar] [CrossRef]
- Tu, M.L.; Feng, T.L.; Wang, Z.Y.; Qiao, M.L.; Shahidi, F.; Lu, W.H.; Du, M. Sequence analysis and molecular docking of antithrombotic peptides from casein hydrolysate by trypsin digestion. J. Funct. Foods 2017, 32, 313–323. [Google Scholar] [CrossRef]
- Lin, K.; Zhang, L.W.; Han, X.; Cheng, D.Y. Novel angiotensin I-converting enzyme inhibitory peptides from protease hydrolysates of Qula casein: Quantitative structure-activity relationship modeling and molecular docking study. J. Funct. Foods 2017, 32, 266–277. [Google Scholar] [CrossRef]
- Yang, W.G.; Wang, Z.; Xu, S.Y. A new method for determination of antithrombotic activity of egg white protein hydrolysate by microplate reader. Chin. Chem. Lett. 2007, 18, 449–451. [Google Scholar] [CrossRef]
- Zhang, S.B.; Wang, Z.; Xu, S.Y. Antioxidant and antithrombotic activities of rapeseed peptides. J. Am. Oil Chem. Soc. 2008, 85, 521–527. [Google Scholar] [CrossRef]
- Charles, R.S.T.; Padmanabhan, K.; Arni, R.V. Structure of tick anticoagulant peptide at 1.6 Å resolution complexed with bovine pancreatic trypsin inhibitor. Protein Sci. 2000, 9, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Cong, W.; Xu, H.; Liu, Y.; Li, Q.Z.; Li, W.; Zhou, X.W. Production and functional characterization of a novel fungal immunomodulatory protein FIP-SN15 shuffled from two genes of ganoderma species. Appl. Microbiol. Biol. 2014, 98, 5967–5975. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sequence | Source | –CDOCKER ENERGY | Toxin | Allergic |
|---|---|---|---|---|
| ELEDSLDSER | P2, P3 | 191.202 | non | non |
| ITIQQELEDAR | P2 | 189.481 | non | non |
| STELLIR | P2, P3 | 188.957 | non | non |
| EKDEEIDSIR | P3 | 182.45 | non | non |
| GDEELDSLIK | P2, P3 | 174.548 | non | non |
| EQEAKAELQR | P3 | 173.992 | non | non |
| ISMLEEDIMK | P1, P2, P3 | 169.018 | non | non |
| LAITEVDLER | P3 | 168.65 | non | non |
| QEELEEIRR | P2, P3 | 164.775 | non | non |
| IEDDYNSLQK | P1, P2, P3 | 164.759 | non | non |
| LEDELLTEK | P1 | 163.193 | non | non |
| LEEAEAQALK | P2 | 163.148 | non | non |
| KLAITEVDLER | P3 | 162.253 | non | non |
| QEELEEIR | P2, P3 | 160.569 | non | non |
| ELEDLSER | P1, P2, P3 | 160.178 | non | non |
| ALADEGSDIK | P1 | 160.159 | non | non |
| NSFVNDIFER | P3 | 158.605 | non | non |
| EDSYEETIR | P2 | 157.024 | non | non |
| DLDSDVSSTR | P2, P3 | 155.361 | non | non |
| VSDLAEDMKR | P3 | 154.278 | non | non |
| EIAEILYDR | P1, P2 | 153.187 | non | non |
| VEFLDDSNR | P1, P2 | 153.163 | non | non |
| WIAEEADK | P1, P2, P3 | 152.732 | non | non |
| VSDLAEDMK | P3 | 152.661 | non | non |
| EIVDLVLDR | P1, P2 | 152.039 | non | non |
| LQSEVTEINR | P3 | 151.634 | non | non |
| VISLTDDTSK | P1, P2 | 151.11 | non | non |
| IDALEGSNSR | P2, P3 | 150.169 | non | non |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qiao, M.; Tu, M.; Wang, Z.; Mao, F.; Chen, H.; Qin, L.; Du, M. Identification and Antithrombotic Activity of Peptides from Blue Mussel (Mytilus edulis) Protein. Int. J. Mol. Sci. 2018, 19, 138. https://doi.org/10.3390/ijms19010138
Qiao M, Tu M, Wang Z, Mao F, Chen H, Qin L, Du M. Identification and Antithrombotic Activity of Peptides from Blue Mussel (Mytilus edulis) Protein. International Journal of Molecular Sciences. 2018; 19(1):138. https://doi.org/10.3390/ijms19010138
Chicago/Turabian StyleQiao, Meiling, Maolin Tu, Zhenyu Wang, Fengjiao Mao, Hui Chen, Lei Qin, and Ming Du. 2018. "Identification and Antithrombotic Activity of Peptides from Blue Mussel (Mytilus edulis) Protein" International Journal of Molecular Sciences 19, no. 1: 138. https://doi.org/10.3390/ijms19010138
APA StyleQiao, M., Tu, M., Wang, Z., Mao, F., Chen, H., Qin, L., & Du, M. (2018). Identification and Antithrombotic Activity of Peptides from Blue Mussel (Mytilus edulis) Protein. International Journal of Molecular Sciences, 19(1), 138. https://doi.org/10.3390/ijms19010138