Regenerative Capacity of Endogenous Factor: Growth Differentiation Factor 11; a New Approach of the Management of Age-Related Cardiovascular Events

 ,
,
Abstract
:
{kind=link}
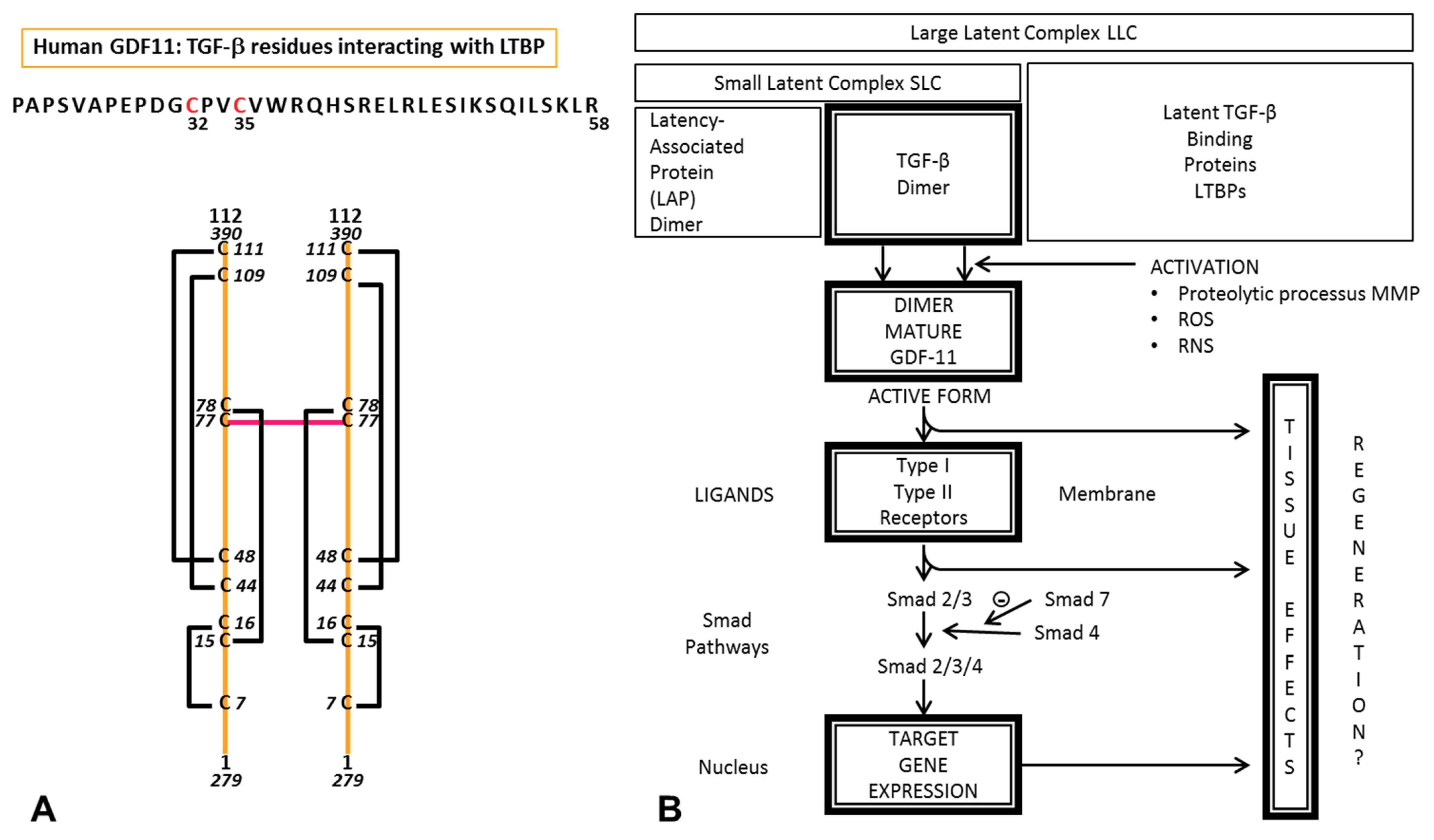{kind=link}
{kind=link}
1. Introduction
2. Mechanistic and Evolutionary Bases of Aging
3. The Surgical Technique of Parabiosis
4. Does GDF11 Reverse Age-Related Functional Impairment in Heart?
5. Does GDF11 Reverse Age-Related Functional Impairment in Skeletal Muscle?
6. Does GDF11 Reverse Age-Related Functional Impairment in Brain? Importance of the Neurovascular Unit
7. Limits and Criticism Concerning the Properties of GDF11: What is the Specificity of the Actions of GDF11?
8. Summary
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ACVRIIB | Activin receptor type IIB |
| ALK | Acting receptor-like-kinase |
| B2M | Beta-2 microglobulin |
| BMP | Bone morphogenetic proteins |
| CAD | Coronary artery disease |
| CCL11 | C-C motif chemokine 1 |
| CHF | Cardiac heart failure |
| CNS | Central nervous system |
| CVD | Cardiovascular disease |
| Foxo | Class O of forkhead box |
| GDF | Growth differentiation factor |
| HP | Heterochronic parabiotic |
| IP | Isochronic parabiotic |
| IPs | Induced pluripotent stem |
| MDSCs | Muscle derived stem cells |
| MI | Myocardial infarction |
| MMPs | Matrix metalloproteinases |
| ROS | Reactive oxygen species |
| RNS | Reactive nitrogen species |
| SIRT | Sirtuin |
| TGF | Transforming growth factor |
References
- Franceschi, C.; Bonafe, M.; Valensin, S.; Olivieri, F.; De Luca, M.; Ottaviani, E.; De Benedictis, G. Inflamm-aging. An evolutionary perspective on immunosenescence. Ann. N. Y. Acad. Sci. 2000, 908, 244–254. [Google Scholar] [CrossRef] [PubMed]
- Newgard, C.B.; Sharpless, N.E. Coming of age: Molecular drivers of aging and therapeutic opportunities. J. Clin. Investig. 2013, 123, 946–950. [Google Scholar] [CrossRef] [PubMed]
- Freije, J.M.; Lopez-Otin, C. Reprogramming aging and progeria. Curr. Opin. Cell Biol. 2012, 24, 757–764. [Google Scholar] [CrossRef] [PubMed]
- Rota, M.; Leri, A.; Anversa, P. Human heart failure: Is cell therapy a valid option? Biochem. Pharmacol. 2014, 88, 129–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leri, A.; Rota, M.; Hosoda, T.; Goichberg, P.; Anversa, P. Cardiac stem cell niches. Stem Cell Res. 2014, 13, 631–646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oudot, A.; Martin, C.; Busseuil, D.; Vergely, C.; Demaison, L.; Rochette, L. NADPH oxidases are in part responsible for increased cardiovascular superoxide production during aging. Free Radic. Biol. Med. 2006, 40, 2214–2222. [Google Scholar] [CrossRef] [PubMed]
- Rochette, L.; Lorin, J.; Zeller, M.; Guilland, J.C.; Lorgis, L.; Cottin, Y.; Vergely, C. Nitric oxide synthase inhibition and oxidative stress in cardiovascular diseases: Possible therapeutic targets? Pharmacol. Ther. 2013, 140, 239–257. [Google Scholar] [CrossRef]
- Cheitlin, M.D. Cardiovascular physiology-changes with aging. Am. J. Geriat. Cardiol. 2003, 12, 9–13. [Google Scholar] [CrossRef]
- Demontis, F.; Piccirillo, R.; Goldberg, A.L.; Perrimon, N. The influence of skeletal muscle on systemic aging and lifespan. Aging Cell 2013, 12, 943–949. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Karpac, J.; Jasper, H. Promoting longevity by maintaining metabolic and proliferative homeostasis. J. Exp. Biol. 2014, 217, 109–118. [Google Scholar] [CrossRef]
- Pillai, V.B.; Sundaresan, N.R.; Gupta, M.P. Regulation of Akt signaling by sirtuins: Its implication in cardiac hypertrophy and aging. Circ. Res. 2014, 114, 368–378. [Google Scholar] [CrossRef] [PubMed]
- Segers, V.F.; Lee, R.T. Protein therapeutics for cardiac regeneration after myocardial infarction. J. Cardiovasc. Transl. Res. 2010, 3, 469–477. [Google Scholar] [CrossRef] [PubMed]
- Haider, H.; Lei, Y.; Ashraf, M. MyoCell, a cell-based, autologous skeletal myoblast therapy for the treatment of cardiovascular diseases. Curr. Opin. Mol. Ther. 2008, 10, 611–621. [Google Scholar] [PubMed]
- Sanganalmath, S.K.; Bolli, R. Cell therapy for heart failure: A comprehensive overview of experimental and clinical studies, current challenges, and future directions. Circ. Res. 2013, 113, 810–834. [Google Scholar] [CrossRef] [PubMed]
- Rochette, L.; Zeller, M.; Cottin, Y.; Vergely, C. Growth and differentiation factor 11 (GDF11): Functions in the regulation of erythropoiesis and cardiac regeneration. Pharmacol. Ther. 2015, 156, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Kondas, K.; Szlama, G.; Nagy, A.; Trexler, M.; Patthy, L. Biological functions of the WAP domain-containing multidomain proteins WFIKKN1 and WFIKKN2. Biochem. Soc. Trans. 2011, 39, 1416–1420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McPherron, A.C. Metabolic Functions of Myostatin and Gdf11. Immunol. Endocr. Metab. Agents Med. Chem. 2010, 10, 217–231. [Google Scholar] [CrossRef] [PubMed]
- Egerman, M.A.; Cadena, S.M.; Gilbert, J.A.; Meyer, A.; Nelson, H.N.; Swalley, S.E.; Mallozzi, C.; Jacobi, C.; Jennings, L.L.; Clay, I.; et al. GDF11 Increases with Age and Inhibits Skeletal Muscle Regeneration. Cell Metab. 2015, 22, 164–174. [Google Scholar] [CrossRef] [Green Version]
- Dschietzig, T.B. Myostatin—From the Mighty Mouse to cardiovascular disease and cachexia. Clin. Chim. Acta Int. J. Clin. Chem. 2014, 433, 216–224. [Google Scholar] [CrossRef]
- Poggioli, T.; Vujic, A.; Yang, P.; Macias-Trevino, C.; Uygur, A.; Loffredo, F.S.; Pancoast, J.R.; Cho, M.; Goldstein, J.; Tandias, R.M.; et al. Circulating Growth Differentiation Factor 11/8 Levels Decline With Age. Circ. Res. 2016, 118, 29–37. [Google Scholar] [CrossRef] [Green Version]
- Brun, C.E.; Rudnicki, M.A. GDF11 and the Mythical Fountain of Youth. Cell Metab. 2015, 22, 54–56. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Wu, H.H.; Lander, A.D.; Lyons, K.M.; Matzuk, M.M.; Calof, A.L. GDF11 controls the timing of progenitor cell competence in developing retina. Science 2005, 308, 1927–1930. [Google Scholar] [CrossRef] [PubMed]
- Conboy, M.J.; Conboy, I.M.; Rando, T.A. Heterochronic parabiosis: Historical perspective and methodological considerations for studies of aging and longevity. Aging Cell 2013, 12, 525–530. [Google Scholar] [CrossRef] [PubMed]
- Song, G.; Nguyen, D.T.; Pietramaggiori, G.; Scherer, S.; Chen, B.; Zhan, Q.; Ogawa, R.; Yannas, I.V.; Wagers, A.J.; Orgill, D.P.; et al. Use of the parabiotic model in studies of cutaneous wound healing to define the participation of circulating cells. Wound Repair Regen. 2010, 18, 426–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamran, P.; Sereti, K.I.; Zhao, P.; Ali, S.R.; Weissman, I.L.; Ardehali, R. Parabiosis in mice: A detailed protocol. J. Vis. Exp. JoVE 2013. [Google Scholar] [CrossRef] [PubMed]
- Loffredo, F.S.; Steinhauser, M.L.; Jay, S.M.; Gannon, J.; Pancoast, J.R.; Yalamanchi, P.; Sinha, M.; Dall'Osso, C.; Khong, D.; Shadrach, J.L.; et al. Growth differentiation factor 11 is a circulating factor that reverses age-related cardiac hypertrophy. Cell 2013, 153, 828–839. [Google Scholar] [CrossRef] [PubMed]
- Mendelsohn, A.R.; Larrick, J.W. Rejuvenation of aging hearts. Rejuv. Res. 2013, 16, 330–332. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.C.; Zhang, X.; Zhang, X.; Gross, P.; Starosta, T.; Mohsin, S.; Franti, M.; Gupta, P.; Hayes, D.; Myzithras, M.; et al. GDF11 does not rescue aging-related pathological hypertrophy. Circ. Res. 2015, 117, 926–932. [Google Scholar] [CrossRef]
- Olson, K.A.; Beatty, A.L.; Heidecker, B.; Regan, M.C.; Brody, E.N.; Foreman, T.; Kato, S.; Mehler, R.E.; Singer, B.S.; Hveem, K.; et al. Association of growth differentiation factor 11/8, putative anti-ageing factor, with cardiovascular outcomes and overall mortality in humans: Analysis of the Heart and Soul and HUNT3 cohorts. Eur. Heart J. 2015, 36, 3426–3434. [Google Scholar] [CrossRef]
- Villeda, S.A.; Plambeck, K.E.; Middeldorp, J.; Castellano, J.M.; Mosher, K.I.; Luo, J.; Smith, L.K.; Bieri, G.; Lin, K.; Berdnik, D.; et al. Young blood reverses age-related impairments in cognitive function and synaptic plasticity in mice. Nat. Med. 2014, 20, 659–663. [Google Scholar] [CrossRef] [Green Version]
- Leung, R.; Proitsi, P.; Simmons, A.; Lunnon, K.; Guntert, A.; Kronenberg, D.; Pritchard, M.; Tsolaki, M.; Mecocci, P.; Kloszewska, I.; et al. Inflammatory proteins in plasma are associated with severity of Alzheimer's disease. PLoS ONE 2013, 8, e64971. [Google Scholar] [CrossRef] [PubMed]
- Suragani, R.N.; Cadena, S.M.; Cawley, S.M.; Sako, D.; Mitchell, D.; Li, R.; Davies, M.V.; Alexander, M.J.; Devine, M.; Loveday, K.S.; et al. Transforming growth factor-beta superfamily ligand trap ACE-536 corrects anemia by promoting late-stage erythropoiesis. Nat. Med. 2014, 20, 408–414. [Google Scholar] [CrossRef] [PubMed]
- Dussiot, M.; Maciel, T.T.; Fricot, A.; Chartier, C.; Negre, O.; Veiga, J.; Grapton, D.; Paubelle, E.; Payen, E.; Beuzard, Y.; et al. An activin receptor IIA ligand trap corrects ineffective erythropoiesis in beta-thalassemia. Nat. Med. 2014, 20, 398–407. [Google Scholar] [CrossRef] [PubMed]
- Rochette, L.; Gudjoncik, A.; Guenancia, C.; Zeller, M.; Cottin, Y.; Vergely, C. The iron-regulatory hormone hepcidin: A possible therapeutic target? Pharmacol. Ther. 2015, 146, 35–52. [Google Scholar] [CrossRef] [PubMed]
- Murrant, C.L.; Sarelius, I.H. Coupling of muscle metabolism and muscle blood flow in capillary units during contraction. Acta Physiol. Scand. 2000, 168, 531–541. [Google Scholar] [CrossRef] [PubMed]
- Brack, A.S.; Conboy, M.J.; Roy, S.; Lee, M.; Kuo, C.J.; Keller, C.; Rando, T.A. Increased Wnt signaling during aging alters muscle stem cell fate and increases fibrosis. Science 2007, 317, 807–810. [Google Scholar] [CrossRef]
- Motohashi, N.; Asakura, A. Muscle satellite cell heterogeneity and self-renewal. Front. Cell Dev. Biol. 2014, 2, 1. [Google Scholar] [CrossRef] [PubMed]
- Burks, T.N.; Cohn, R.D. Role of TGF-beta signaling in inherited and acquired myopathies. Skeletal Muscle 2011, 1, 19. [Google Scholar] [CrossRef]
- Brooks, N.E.; Myburgh, K.H. Skeletal muscle wasting with disuse atrophy is multi-dimensional: The response and interaction of myonuclei, satellite cells and signaling pathways. Front. Physiol. 2014, 5, 99. [Google Scholar] [CrossRef]
- Conboy, I.M.; Conboy, M.J.; Wagers, A.J.; Girma, E.R.; Weissman, I.L.; Rando, T.A. Rejuvenation of aged progenitor cells by exposure to a young systemic environment. Nature 2005, 433, 760–764. [Google Scholar] [CrossRef]
- Sinha, M.; Jang, Y.C.; Oh, J.; Khong, D.; Wu, E.Y.; Manohar, R.; Miller, C.; Regalado, S.G.; Loffredo, F.S.; Pancoast, J.R.; et al. Restoring systemic GDF11 levels reverses age-related dysfunction in mouse skeletal muscle. Science 2014, 344, 649–652. [Google Scholar] [CrossRef] [PubMed]
- Tarantini, S.; Tran, C.H.T.; Gordon, G.R.; Ungvari, Z.; Csiszar, A. Impaired neurovascular coupling in aging and Alzheimer's disease: Contribution of astrocyte dysfunction and endothelial impairment to cognitive decline. Exp. Gerontol. 2017, 94, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Hillary, F.G.; Biswal, B. The influence of neuropathology on the FMRI signal: A measurement of brain or vein? Clin. Neuropsychol. 2007, 21, 58–72. [Google Scholar] [CrossRef] [PubMed]
- Grady, C.L. Cognitive neuroscience of aging. Ann. N. Y. Acad. Sci. 2008, 1124, 127–144. [Google Scholar] [CrossRef] [PubMed]
- Flores, I.; Canela, A.; Vera, E.; Tejera, A.; Cotsarelis, G.; Blasco, M.A. The longest telomeres: A general signature of adult stem cell compartments. Genes Dev. 2008, 22, 654–667. [Google Scholar] [CrossRef] [PubMed]
- Mimeault, M.; Batra, S.K. Recent insights into the molecular mechanisms involved in aging and the malignant transformation of adult stem/progenitor cells and their therapeutic implications. Ageing Res. Rev. 2009, 8, 94–112. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.S.; Iijima-Ando, K.; Iijima, K.; Lee, W.J.; Lee, J.H.; Yu, K.; Lee, D.S. JNK/FOXO-mediated neuronal expression of fly homologue of peroxiredoxin II reduces oxidative stress and extends life span. J. Biol. Chem. 2009, 284, 29454–29461. [Google Scholar] [CrossRef]
- Villeda, S.A.; Luo, J.; Mosher, K.I.; Zou, B.; Britschgi, M.; Bieri, G.; Stan, T.M.; Fainberg, N.; Ding, Z.; Eggel, A.; et al. The ageing systemic milieu negatively regulates neurogenesis and cognitive function. Nature 2011, 477, 90–94. [Google Scholar] [CrossRef] [Green Version]
- Ruckh, J.M.; Zhao, J.W.; Shadrach, J.L.; van Wijngaarden, P.; Rao, T.N.; Wagers, A.J.; Franklin, R.J. Rejuvenation of regeneration in the aging central nervous system. Cell Stem Cell 2012, 10, 96–103. [Google Scholar] [CrossRef]
- Katsimpardi, L.; Litterman, N.K.; Schein, P.A.; Miller, C.M.; Loffredo, F.S.; Wojtkiewicz, G.R.; Chen, J.W.; Lee, R.T.; Wagers, A.J.; Rubin, L.L. Vascular and neurogenic rejuvenation of the aging mouse brain by young systemic factors. Science 2014, 344, 630–634. [Google Scholar] [CrossRef]
- Tsuchida, K.; Nakatani, M.; Uezumi, A.; Murakami, T.; Cui, X. Signal transduction pathway through activin receptors as a therapeutic target of musculoskeletal diseases and cancer. Endocr. J. 2008, 55, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Xiang, B.; Xiao, C.; Shen, T.; Li, X. Anti-inflammatory effects of anisalcohol on lipopolysaccharide-stimulated BV2 microglia via selective modulation of microglia polarization and down-regulation of NF-kappaB p65 and JNK activation. Mol. Immunol. 2018, 95, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.; Brown, C.W.; Matzuk, M.M. Genetic analysis of the mammalian transforming growth factor-beta superfamily. Endocr. Rev. 2002, 23, 787–823. [Google Scholar] [CrossRef] [PubMed]
- McNally, E.M. Questions and Answers About Myostatin, GDF11, and the Aging Heart. Circ. Res. 2016, 118, 6–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McPherron, A.C.; Lawler, A.M.; Lee, S.J. Regulation of anterior/posterior patterning of the axial skeleton by growth/differentiation factor 11. Nat. Genet. 1999, 22, 260–264. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; McMahon, A.P.; Valerius, M.T. Recent genetic studies of mouse kidney development. Curr. Opin. Genet. Dev. 2004, 14, 550–557. [Google Scholar] [CrossRef] [PubMed]
- Girgenrath, S.; Song, K.; Whittemore, L.A. Loss of myostatin expression alters fiber-type distribution and expression of myosin heavy chain isoforms in slow- and fast-type skeletal muscle. Muscle Nerve 2005, 31, 34–40. [Google Scholar] [CrossRef] [PubMed]
- McPherron, A.C.; Huynh, T.V.; Lee, S.J. Redundancy of myostatin and growth/differentiation factor 11 function. BMC Dev. biol. 2009, 9, 24. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, J.; Vernus, B.; Chelh, I.; Cassar-Malek, I.; Gabillard, J.C.; Hadj Sassi, A.; Seiliez, I.; Picard, B.; Bonnieu, A. Myostatin and the skeletal muscle atrophy and hypertrophy signaling pathways. Cell. Mol. Life Sci. CMLS 2014, 71, 4361–4371. [Google Scholar] [CrossRef] [PubMed]
- Gilson, H.; Schakman, O.; Kalista, S.; Lause, P.; Tsuchida, K.; Thissen, J.P. Follistatin induces muscle hypertrophy through satellite cell proliferation and inhibition of both myostatin and activin. Am. J. Physiol. Endocrinol. Metab. 2009, 297, E157–E164. [Google Scholar] [CrossRef] [PubMed]
- Bose, B.; Shenoy, P.S. Pluripotent Conversion of Muscle Stem Cells Without Reprogramming Factors or Small Molecules. Stem Cell Rev. 2016, 12, 73–89. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Su, X.; Ashraf, M.; Kim, I.M.; Weintraub, N.L.; Jiang, M.; Tang, Y. Regenerative Therapy for Cardiomyopathies. J. Cardiovasc. Transl. Res. 2018. [Google Scholar] [CrossRef] [PubMed]
- Yousef, H.; Conboy, M.J.; Morgenthaler, A.; Schlesinger, C.; Bugaj, L.; Paliwal, P.; Greer, C.; Conboy, I.M.; Schaffer, D. Systemic attenuation of the TGF-beta pathway by a single drug simultaneously rejuvenates hippocampal neurogenesis and myogenesis in the same old mammal. Oncotarget 2015, 6, 11959–11978. [Google Scholar] [CrossRef] [PubMed]
- Elabd, C.; Cousin, W.; Upadhyayula, P.; Chen, R.Y.; Chooljian, M.S.; Li, J.; Kung, S.; Jiang, K.P.; Conboy, I.M. Oxytocin is an age-specific circulating hormone that is necessary for muscle maintenance and regeneration. Nat. Commun. 2014, 5, 4082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, L.K.; He, Y.; Park, J.S.; Bieri, G.; Snethlage, C.E.; Lin, K.; Gontier, G.; Wabl, R.; Plambeck, K.E.; Udeochu, J.; et al. beta2-microglobulin is a systemic pro-aging factor that impairs cognitive function and neurogenesis. Nat. Med. 2015, 21, 932–937. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Li, F.; Long, T.; Li, F.; Peng, L.; Xia, K.; Jing, R.; Xie, Q.; Yang, T. Beta 2-Microglobulin and the Severity of Coronary Stenosis in Patients With Acute Coronary Syndrome. Heart Lung Circ. 2018. [Google Scholar] [CrossRef] [PubMed]
- Bouchard, J.; Villeda, S.A. Aging and brain rejuvenation as systemic events. J. Neurochem. 2015, 132, 5–19. [Google Scholar] [CrossRef] [PubMed]


© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rochette, L.; Meloux, A.; Rigal, E.; Zeller, M.; Cottin, Y.; Malka, G.; Vergely, C. Regenerative Capacity of Endogenous Factor: Growth Differentiation Factor 11; a New Approach of the Management of Age-Related Cardiovascular Events. Int. J. Mol. Sci. 2018, 19, 3998. https://doi.org/10.3390/ijms19123998
Rochette L, Meloux A, Rigal E, Zeller M, Cottin Y, Malka G, Vergely C. Regenerative Capacity of Endogenous Factor: Growth Differentiation Factor 11; a New Approach of the Management of Age-Related Cardiovascular Events. International Journal of Molecular Sciences. 2018; 19(12):3998. https://doi.org/10.3390/ijms19123998
Chicago/Turabian StyleRochette, Luc, Alexandre Meloux, Eve Rigal, Marianne Zeller, Yves Cottin, Gabriel Malka, and Catherine Vergely. 2018. "Regenerative Capacity of Endogenous Factor: Growth Differentiation Factor 11; a New Approach of the Management of Age-Related Cardiovascular Events" International Journal of Molecular Sciences 19, no. 12: 3998. https://doi.org/10.3390/ijms19123998