Inflammatory Alteration of Human T Cells Exposed Continuously to Asbestos


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
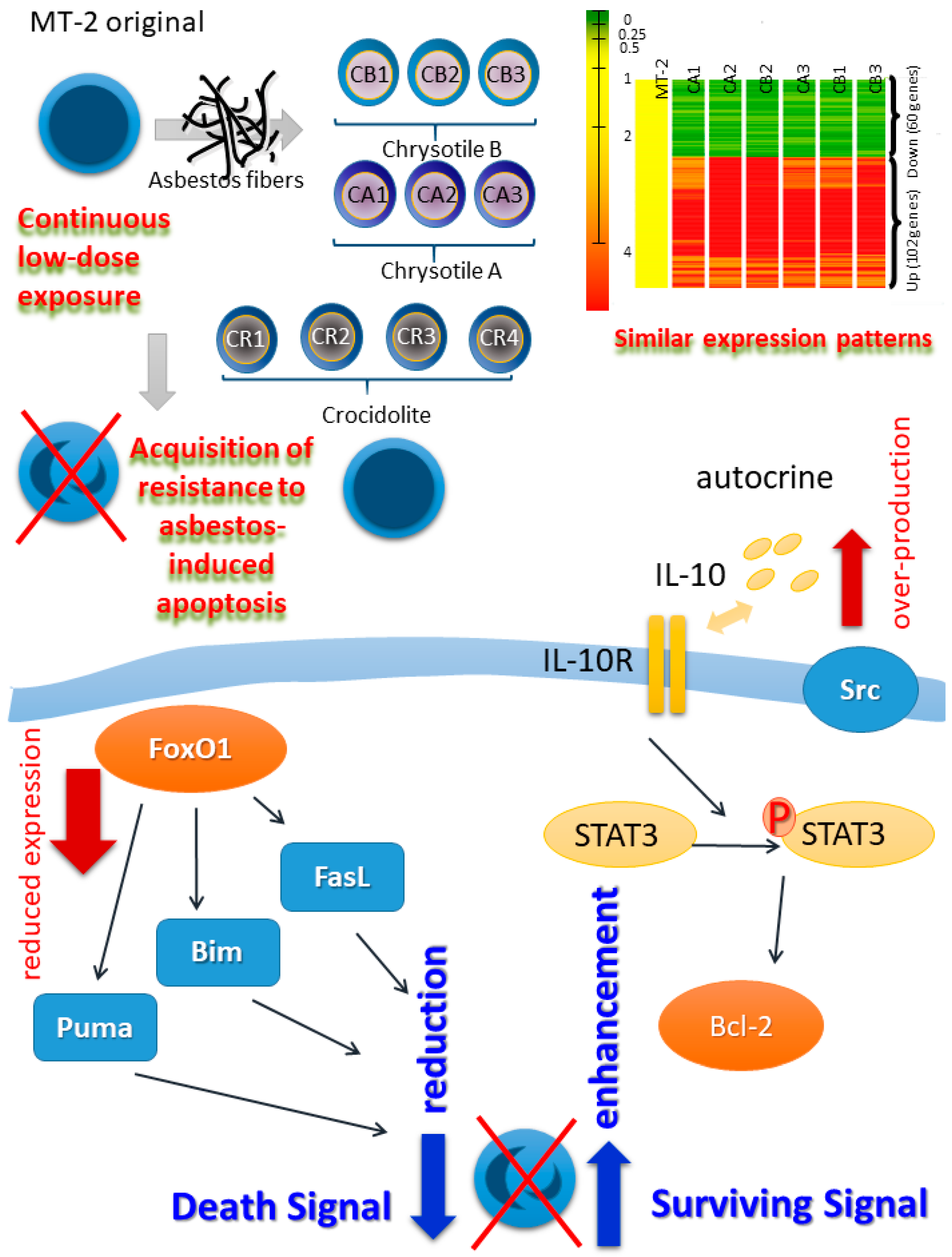
2. Establishment of a Human T Cell Model for Continuous and Low-Dose Exposure to Asbestos Fibers
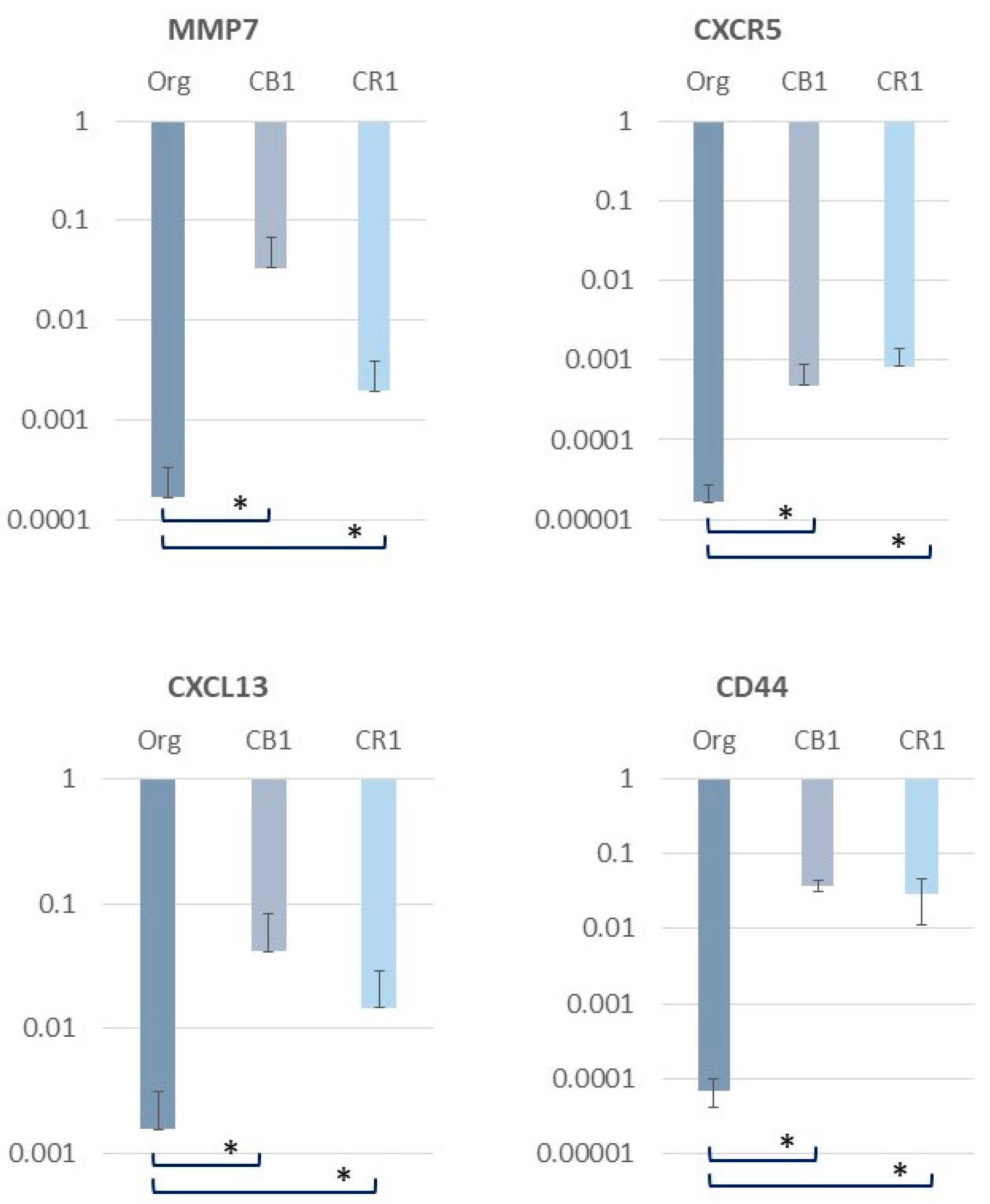
3. Alterations in Inflammation-Related Molecules in MT-2 Sublines Continuously Exposed to Asbestos
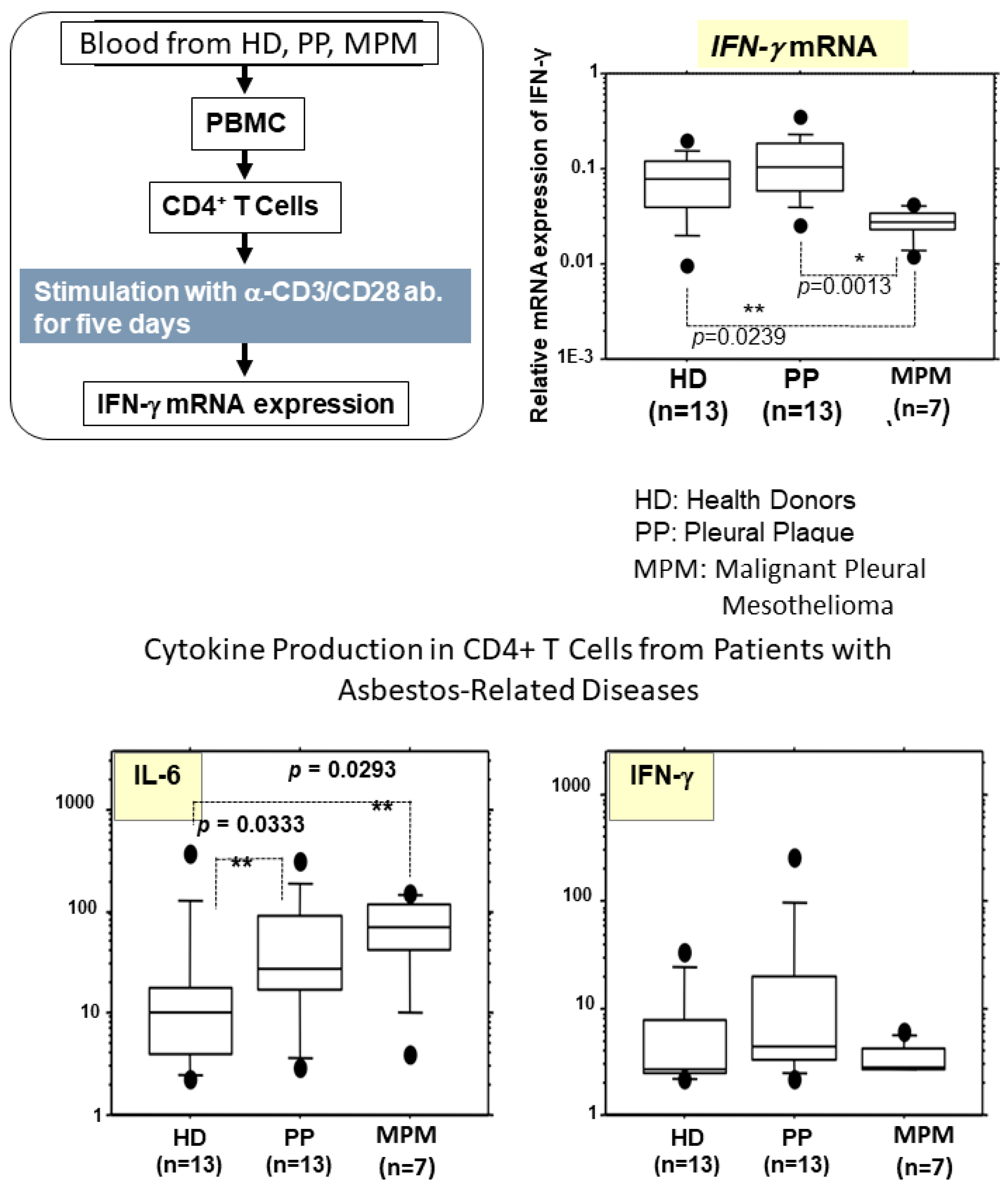
4. Alterations in Cytokine Production in Peripheral T Cells Derived from Asbestos-Exposed Patients
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| PP | pleural plaque |
| MPM | malignant pleural mesothelioma |
| IARC | International Agency for Research on Cancer |
| ROS | reactive oxygen species |
| RNS | reactive nitrogen species |
| AMs | alveolar macrophages |
| APC | antigen-presenting cells |
| ASC | apoptosis-associated speck-like protein containing a CARD |
| NLRs | nucleotide-binding oligomerization domain-like receptors (NOD-like receptors) |
| IL | interleukin |
| EBVLCLs | Epstein-Barr virus immortalized lymphoblastoid cell lines |
| HTLV | human T cell leukemia/lymphoma virus |
| MAPK | mitogen-activated protein kinase |
| FoxO1 | forkhead box protein O1 |
| STAT3 | signal transducer and activator of transcription 3 |
| CXCR3 | C-X-C motif chemokine receptor 3 |
| Treg | regulatory T cell |
| TGF | transforming growth factor |
| CDK-Is | cyclin-dependent kinases-inhibitors |
| MMP-7 | Matrix metalloproteinase-7 |
| CXCL13 | chemokine (C-X-C motif) ligand 13 |
| BLC | B lymphocyte chemoattractant |
| BCA-1 | B cell-attracting chemokine 1 |
| IFN | interferon |
| HD | healthy donors |
| NK | natural killer |
| CTLs | cytotoxic T lymphocytes |
References
- Lemen, R.A. Epidemiology of asbestos-related diseases and the knowledge that led to what is known today. In Asbestos: Risk Assessment, Epidemiology, and Health Effects, 2nd ed.; Dodson, R.F., Hammar, S.P., Eds.; CRC Press: Boca Ratonm, FL, USA, 2011. [Google Scholar]
- Craighead, J.E. Diseases associated with asbestos industrial products and environmental exposure. In Asbestos and Its Diseases; Craighead, J.E., Gibbs, A.R., Eds.; Oxford University Press: New York, NY, USA, 2008. [Google Scholar]
- Hammar, S.P. Asbestosis. In Asbestos. Risk Assessment, Epidemiology, and Health Effects, 2nd ed.; Dodson, R.F., Hammar, S.P., Eds.; CRC Press: Boca Ratonm, FL, USA, 2011. [Google Scholar]
- Lazarus, A.A.; Philip, A. Asbestosis. Dis. Mon. 2011, 57, 14–26. [Google Scholar] [CrossRef] [PubMed]
- Craighead, J.E. Benign pleural and parenchymal diseases associated with asbestos exposure. In Asbestos and Its Diseases; Craighead, J.E., Gibbs, A.R., Eds.; Oxford University Press: New York, NY, USA, 2008. [Google Scholar]
- Heintz, N.H.; Janssen-Heininger, Y.M.; Mossman, B.T. Asbestos, lung cancers, and mesotheliomas: From molecular approaches to targeting tumor survival pathways. Am. J. Respir. Cell Mol. Biol. 2010, 42, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Attanoos, R.L. Asbestos-related lung disease. Surg. Pathol. Clin. 2010, 3, 109–127. [Google Scholar] [CrossRef] [PubMed]
- Lazarus, A.; Massoumi, A.; Hostler, J.; Hostler, D.C. Asbestos-related pleuropulmonary diseases: Benign and malignant. Postgrad. Med. 2012, 124, 116–130. [Google Scholar] [CrossRef] [PubMed]
- Wolff, H.; Vehmas, T.; Oksa, P.; Rantanen, J.; Vainio, H. Asbestos, asbestosis, and cancer, the Helsinki criteria for diagnosis and attribution 2014: Recommendations. Scand. J. Work Environ. Health 2015, 41, 5–15. [Google Scholar] [CrossRef] [PubMed]
- International Agency for Research on Cancer. A Review of Human Carcinogens: Arsenic, Metals, Fibres, and Dusts (Iarc Monographs on the Evaluation of the Carcinogenic Risks to Humans); IARC: Lyon, France, 2012. [Google Scholar]
- Kamp, D.W. Asbestos-induced lung diseases: An update. Transl. Res. 2009, 153, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Mahmood, N.; Khan, S.G.; Ali, S.; Athar, M.; Rahman, Q. Asbestos induced oxidative injury to DNA. Ann. Occup. Hyg. 1993, 37, 315–319. [Google Scholar] [CrossRef] [PubMed]
- Simeonova, P.P.; Luster, M.I. Iron and reactive oxygen species in the asbestos-induced tumor necrosis factor-alpha response from alveolar macrophages. Am. J. Respir. Cell Mol. Biol. 1995, 12, 676–683. [Google Scholar] [CrossRef] [PubMed]
- Pociask, D.A.; Sime, P.J.; Brody, A.R. Asbestos-derived reactive oxygen species activate TGF-beta1. Lab. Invest. 2004, 84, 1013–1023. [Google Scholar] [CrossRef] [PubMed]
- Upadhyay, D.; Kamp, D.W. Asbestos-induced pulmonary toxicity: Role of DNA damage and apoptosis. Exp. Biol. Med. 2003, 228, 650–659. [Google Scholar] [CrossRef]
- Liu, G.; Beri, R.; Mueller, A.; Kamp, D.W. Molecular mechanisms of asbestos-induced lung epithelial cell apoptosis. Chem. Biol. Interact. 2010, 188, 309–318. [Google Scholar] [CrossRef] [PubMed]
- Toyokuni, S. Mechanisms of asbestos-induced carcinogenesis. Nagoya J. Med. Sci. 2009, 71, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Dostert, C.; Pétrilli, V.; Van Bruggen, R.; Steele, C.; Mossman, B.T.; Tschopp, J. Innate immune activation through Nalp3 inflammasome sensing of asbestos and silica. Science 2008, 320, 674–677. [Google Scholar] [CrossRef] [PubMed]
- Ng, G.; Chau, E.M.; Shi, Y. Recent developments in immune activation by uric acid crystals. Arch. Immunol. Ther. Exp. 2010, 58, 273–277. [Google Scholar] [CrossRef] [PubMed]
- Rastrick, J.; Birrell, M. The role of the inflammasome in fibrotic respiratory diseases. Min. Med. 2014, 105, 9–23. [Google Scholar]
- Buder-Hoffmann, S.; Palmer, C.; Vacek, P.; Taatjes, D.; Mossman, B. Different accumulation of activated extracellular signal-regulated kinases (ERK 1/2) and role in cell-cycle alterations by epidermal growth factor, hydrogen peroxide, or asbestos in pulmonary epithelial cells. Am. J. Respir. Cell Mol. Biol. 2001, 24, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Aljandali, A.; Pollack, H.; Yeldandi, A.; Li, Y.; Weitzman, S.A.; Kamp, D.W. Asbestos causes apoptosis in alveolar epithelial cells: Role of iron-induced free radicals. J. Lab. Clin. Med. 2001, 137, 330–339. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, S.; Choe, N.; Iwagaki, A.; Hemenway, D.R.; Kagan, E. Asbestos exposure induces MCP-1 secretion by pleural mesothelial cells. Exp. Lung Res. 2000, 26, 241–255. [Google Scholar] [PubMed]
- Jung, M.; Davis, W.P.; Taatjes, D.J.; Churg, A.; Mossman, B.T. Asbestos and cigarette smoke cause increased DNA strand breaks and necrosis in bronchiolar epithelial cells in vivo. Free Radic. Biol. Med. 2000, 28, 1295–1299. [Google Scholar] [CrossRef]
- Riganti, C.; Aldieri, E.; Bergandi, L.; Fenoglio, I.; Costamagna, C.; Fubini, B.; Bosia, A.; Ghigo, D. Crocidolite asbestos inhibits pentose phosphate oxidative pathway and glucose 6-phosphate dehydrogenase activity in human lung epithelial cells. Free Radic. Biol. Med. 2002, 32, 938–949. [Google Scholar] [CrossRef]
- Maeda, M.; Yamamoto, S.; Hatayama, T.; Mastuzaki, H.; Lee, S.; Kumagai-Takei, N.; Yoshitome, K.; Nishimura, Y.; Kimura, Y.; Otsuki, T. T cell alteration caused by exposure to asbestos. In Biological Effects of Fibrous and Particulate Substances; Otsuki, T., Holian, A., Yoshioka, Y., Eds.; Springer: Tokyo, Japan, 2015. [Google Scholar]
- Hyodoh, F.; Takata-Tomokuni, A.; Miura, Y.; Sakaguchi, H.; Hatayama, T.; Hatada, S.; Katsuyama, H.; Matsuo, Y.; Otsuki, T. Inhibitory effects of anti-oxidants on apoptosis of a human polyclonal T-cell line, MT-2, induced by an asbestos, chrysotile-A. Scand. J. Immunol. 2005, 61, 442–448. [Google Scholar] [CrossRef] [PubMed]
- Maeda, M.; Yamamoto, S.; Chen, Y.; Kumagai-Takei, N.; Hayashi, H.; Matsuzaki, H.; Lee, S.; Hatayama, T.; Miyahara, N.; Katoh, M.; et al. Resistance to asbestos-induced apoptosis with continuous exposure to crocidolite on a human T cell. Sci. Total Environ. 2012, 429, 174–182. [Google Scholar] [CrossRef] [PubMed]
- Miura, Y.; Nishimura, Y.; Katsuyama, H.; Maeda, M.; Hayashi, H.; Dong, M.; Hyodoh, F.; Tomita, M.; Matsuo, Y.; Uesaka, A.; et al. Involvement of IL-10 and Bcl-2 in resistance against an asbestos-induced apoptosis of T cells. Apoptosis 2006, 11, 1825–1835. [Google Scholar] [CrossRef] [PubMed]
- Maeda, M.; Nishimura, Y.; Hayashi, H.; Kumagai, N.; Chen, Y.; Murakami, S.; Miura, Y.; Hiratsuka, J.; Kishimoto, T.; Otsuki, T. Reduction of CXC chemokine receptor 3 in an in vitro model of continuous exposure to asbestos in a human T-cell line, MT-2. Am. J. Respir. Cell Mol. Biol. 2011, 45, 470–479. [Google Scholar] [CrossRef] [PubMed]
- Maeda, M.; Nishimura, Y.; Hayashi, H.; Kumagai, N.; Chen, Y.; Murakami, S.; Miura, Y.; Hiratsuka, J.; Kishimoto, T.; Otsuki, T. Decreased CXCR3 expression in CD4+ T cells exposed to asbestos or derived from asbestos-exposed patients. Am. J. Respir. Cell Mol. Biol. 2011, 45, 795–803. [Google Scholar] [CrossRef] [PubMed]
- Matsuzaki, H.; Lee, S.; Maeda, M.; Kumagai-Takei, N.; Nishimura, Y.; Otsuki, T. FoxO1 regulates apoptosis induced by asbestos in the MT-2 human T-cell line. J. Immunotoxicol. 2016, 13, 620–627. [Google Scholar] [CrossRef] [PubMed]
- Ying, C.; Maeda, M.; Nishimura, Y.; Kumagai-Takei, N.; Hayashi, H.; Matsuzaki, H.; Lee, S.; Yoshitome, K.; Yamamoto, S.; Hatayama, T.; et al. Enhancement of regulatory T cell-like suppressive function in MT-2 by long-term and low-dose exposure to asbestos. Toxicology 2015, 338, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Matsuzaki, H.; Maeda, M.; Yamamoto, S.; Kumagai-Takei, N.; Hatayama, T.; Ikeda, M.; Yoshitome, K.; Nishimura, Y.; Otsuki, T. Accelerated cell cycle progression of human regulatory T cell-like cell line caused by continuous exposure to asbestos fibers. Int. J. Oncol. 2017, 50, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Abacherli, L.E.; Nadler, S.T.; Wang, Y.; Li, Q.; Parks, W.C. MMP7 shedding of syndecan-1 facilitates re-epithelialization by affecting α2β1 integrin activation. PLoS ONE 2009, 4. [Google Scholar] [CrossRef] [PubMed]
- Nathan, N.; Corvol, H.; Amselem, S.; Clement, A. Biomarkers in Interstitial lung diseases. Paediatr. Respir. Rev. 2015, 16, 219–224. [Google Scholar] [CrossRef] [PubMed]
- Rohani, M.G.; Parks, W.C. Matrix remodeling by MMPs during wound repair. Matrix Biol. 2015, 44–46, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Moser, B.; Schaerli, P.; Loetscher, P. CXCR5+ T cells: Follicular homing takes center stage in T-helper-cell responses. Trends Immunol. 2002, 23, 250–254. [Google Scholar] [CrossRef]
- Müller, G.; Lipp, M. Shaping up adaptive immunity: The impact of CCR7 and CXCR5 on lymphocyte trafficking. Microcirculation 2003, 10, 325–334. [Google Scholar] [CrossRef] [PubMed]
- Moser, B. CXCR5, the Defining Marker for Follicular B Helper T (TFH) Cells. Front. Immunol. 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- Finch, D.K.; Ettinger, R.; Karnell, J.L.; Herbst, R.; Sleeman, M.A. Effects of CXCL13 inhibition on lymphoid follicles in models of autoimmune disease. Eur. J. Clin. Invest. 2013, 43, 501–509. [Google Scholar] [CrossRef] [PubMed]
- Litsiou, E.; Semitekolou, M.; Galani, I.E.; Morianos, I.; Tsoutsa, A.; Kara, P.; Rontogianni, D.; Bellenis, I.; Konstantinou, M.; Potaris, K.; et al. CXCL13 production in B cells via Toll-like receptor/lymphotoxin receptor signaling is involved in lymphoid neogenesis in chronic obstructive pulmonary disease. Am. J. Respir. Crit. Care Med. 2013, 187, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Vuga, L.J.; Tedrow, J.R.; Pandit, K.V.; Tan, J.; Kass, D.J.; Xue, J.; Chandra, D.; Leader, J.K.; Gibson, K.F.; Kaminski, N.; et al. C-X-C motif chemokine 13 (CXCL13) is a prognostic biomarker of idiopathic pulmonary fibrosis. Am. J. Respir. Crit. Care Med. 2014, 189, 966–974. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Gupta, P.; Kloecker, G.H.; Singh, S.; Lillard, J.W., Jr. Expression and clinical significance of CXCR5/CXCL13 in human non-small cell lung carcinoma. Int. J. Oncol. 2014, 45, 2232–2240. [Google Scholar] [CrossRef] [PubMed]
- Lesley, J.; Hyman, R.; English, N.; Catterall, J.B.; Turner, G.A. CD44 in inflammation and metastasis. Glycoconj. J. 1997, 14, 611–622. [Google Scholar] [CrossRef] [PubMed]
- Heldin, P.; Karousou, E.; Bernert, B.; Porsch, H.; Nishitsuka, K.; Skandalis, S.S. Importance of hyaluronan-CD44 interactions in inflammation and tumorigenesis. Connect. Tissue Res. 2008, 49, 215–218. [Google Scholar] [CrossRef] [PubMed]
- Johnson, P.; Ruffell, B. CD44 and its role in inflammation and inflammatory diseases. Inflamm. Allergy Drug Targets 2009, 8, 208–220. [Google Scholar] [CrossRef] [PubMed]
- Sasai, M.; Nakamura, H.; Sougawa, N.; Sakurai, Y.; Suzuki, M.; Lee, C.M. Novel hyaluronan formulation enhances the efficacy of boron neutron capture therapy for murine mesothelioma. Anticancer. Res. 2016, 36, 907–911. [Google Scholar] [PubMed]
- Murakami, S.; Nishimura, Y.; Maeda, M.; Kumagai, N.; Hayashi, H.; Chen, Y.; Kusaka, M.; Kishimoto, T.; Otsuki, T. Cytokine alteration and speculated immunological pathophysiology in silicosis and asbestos-related diseases. Environ. Health Prev. Med. 2009, 14, 216–222. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, Y.; Miura, Y.; Maeda, M.; Kumagai, N.; Murakami, S.; Hayashi, H.; Fukuoka, K.; Nakano, T.; Otsuki, T. Impairment in cytotoxicity and expression of NK cell- activating receptors on human NK cells following exposure to asbestos fibers. Int. J. Immunopathol. Pharmacol. 2009, 22, 579–590. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, Y.; Maeda, M.; Kumagai, N.; Hayashi, H.; Miura, Y.; Otsuki, T. Decrease in phosphorylation of ERK following decreased expression of NK cell-activating receptors in human NK cell line exposed to asbestos. Int. J. Immunopathol. Pharmacol. 2009, 22, 879–888. [Google Scholar] [CrossRef] [PubMed]
- Kumagai-Takei, N.; Nishimura, Y.; Maeda, M.; Hayashi, H.; Matsuzaki, H.; Lee, S.; Hiratsuka, J.; Otsuki, T. Effect of asbestos exposure on differentiation of cytotoxic T lymphocytes in mixed lymphocyte reaction of human peripheral blood mononuclear cells. Am. J. Respir. Cell Mol. Biol. 2013, 49, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Kumagai-Takei, N.; Nishimura, Y.; Maeda, M.; Hayashi, H.; Matsuzaki, H.; Lee, S.; Kishimoto, T.; Fukuoka, K.; Nakano, T.; Otsuki, T. Functional properties of CD8+ lymphocytes in patients with pleural plaque and malignant mesothelioma. J. Immunol. Res. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Kumagai-Takei, N.; Nishimura, Y.; Matsuzaki, H.; Lee, S.; Yoshitome, K.; Hayashi, H.; Otsuki, T. The suppressed induction of human mature cytotoxic T lymphocytes caused by asbestos is not due to interleukin-2 insufficiency. J. Immunol. Res. 2016, 2016. [Google Scholar] [CrossRef] [PubMed]



© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kumagai-Takei, N.; Yamamoto, S.; Lee, S.; Maeda, M.; Masuzzaki, H.; Sada, N.; Yu, M.; Yoshitome, K.; Nishimura, Y.; Otsuki, T. Inflammatory Alteration of Human T Cells Exposed Continuously to Asbestos. Int. J. Mol. Sci. 2018, 19, 504. https://doi.org/10.3390/ijms19020504
Kumagai-Takei N, Yamamoto S, Lee S, Maeda M, Masuzzaki H, Sada N, Yu M, Yoshitome K, Nishimura Y, Otsuki T. Inflammatory Alteration of Human T Cells Exposed Continuously to Asbestos. International Journal of Molecular Sciences. 2018; 19(2):504. https://doi.org/10.3390/ijms19020504
Chicago/Turabian StyleKumagai-Takei, Naoko, Shoko Yamamoto, Suni Lee, Megumi Maeda, Hidenori Masuzzaki, Nagisa Sada, Min Yu, Kei Yoshitome, Yasumitsu Nishimura, and Takemi Otsuki. 2018. "Inflammatory Alteration of Human T Cells Exposed Continuously to Asbestos" International Journal of Molecular Sciences 19, no. 2: 504. https://doi.org/10.3390/ijms19020504
APA StyleKumagai-Takei, N., Yamamoto, S., Lee, S., Maeda, M., Masuzzaki, H., Sada, N., Yu, M., Yoshitome, K., Nishimura, Y., & Otsuki, T. (2018). Inflammatory Alteration of Human T Cells Exposed Continuously to Asbestos. International Journal of Molecular Sciences, 19(2), 504. https://doi.org/10.3390/ijms19020504