Regulation of Ion Transport in the Intestine by Free Fatty Acid Receptor 2 and 3: Possible Involvement of the Diffuse Chemosensory System


Abstract
:1. Introduction
2. Diffuse Chemosensory System
3. Chemosensory Cells in the Intestine
3.1. Enteroendocrine Cell
3.2. Brush Cells
4. FFA Receptors in the Intestine
5. Involvement of FFA2 and FFA3 on Intestinal Ion Transport
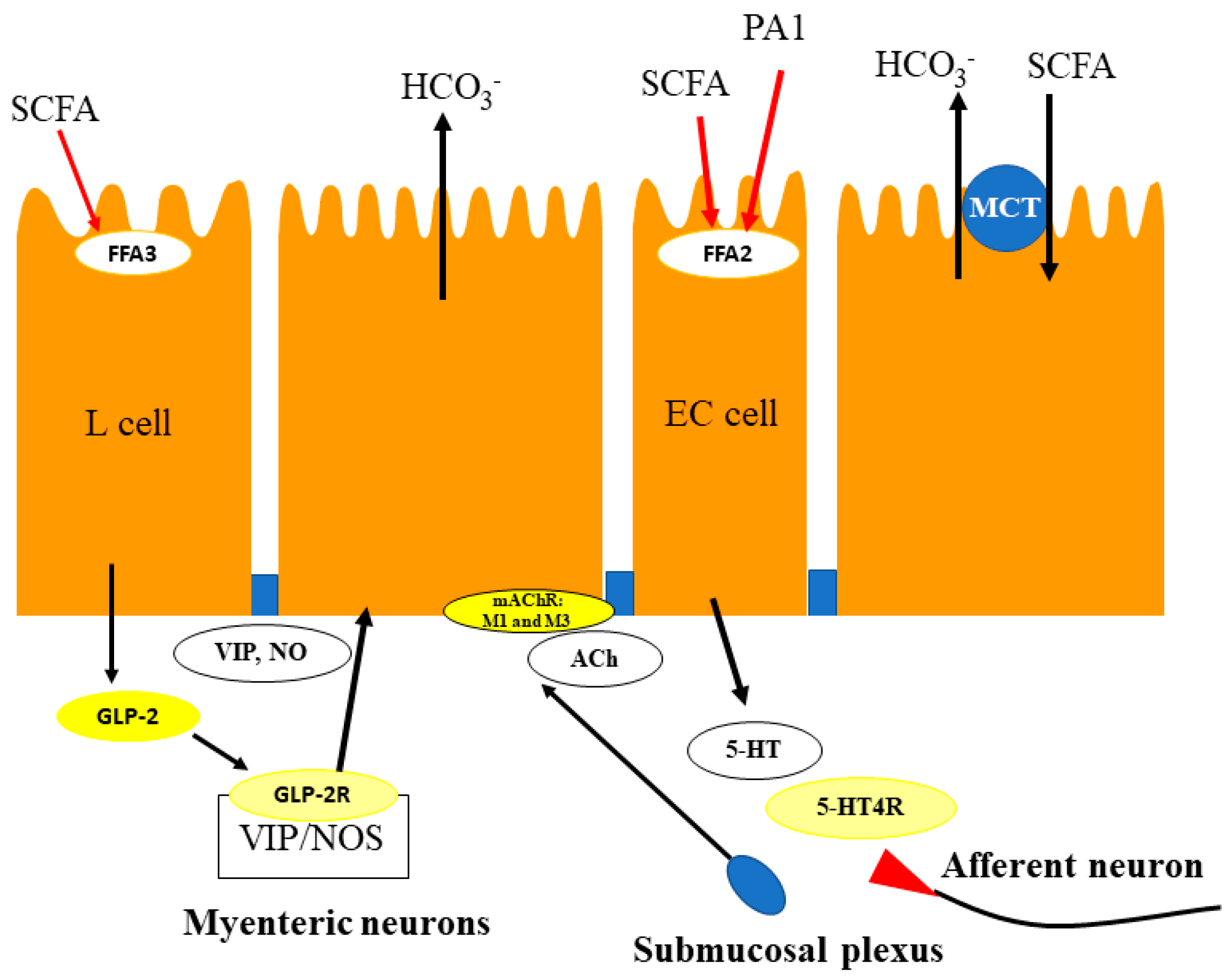
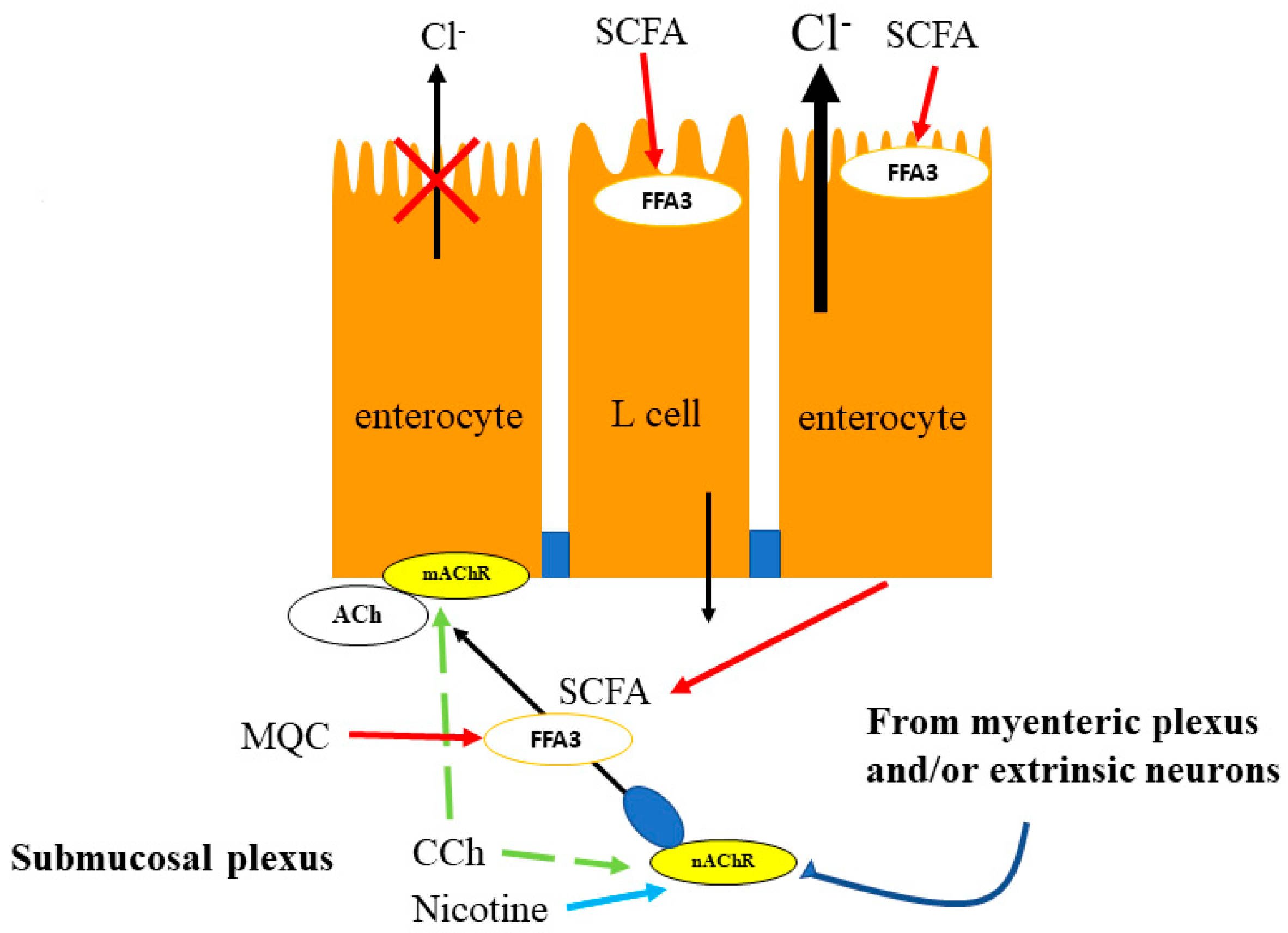
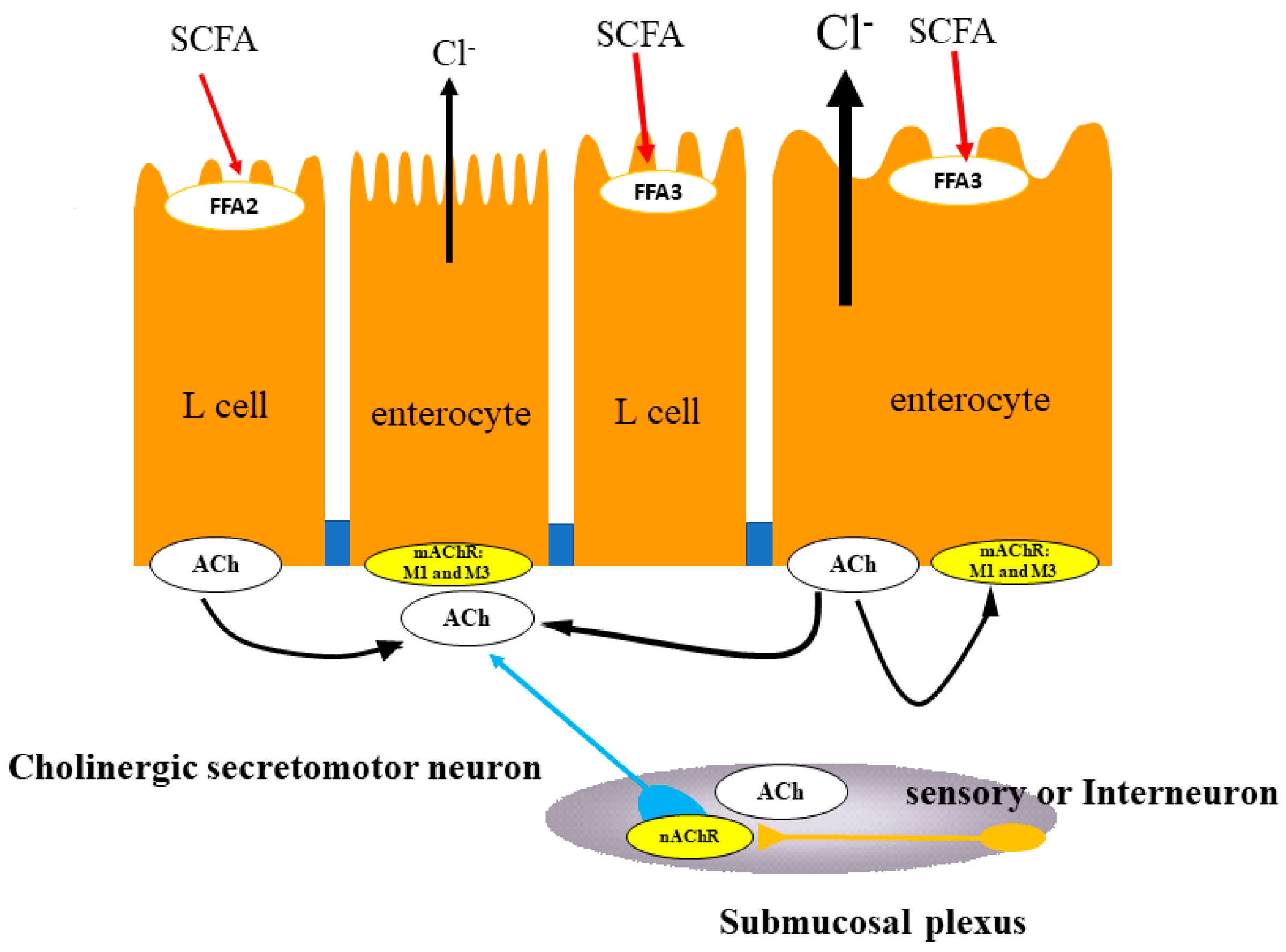
5.1. Duodenum
5.2. Proximal Colon
5.3. Distal Colon
6. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| ACh | acetylcholine |
| CGRP | calcitonin gene-related peptide |
| 4-CHCA | a-cyano-4-hydroxycinnamic acid |
| eNOS | endothelial nitric oxide synthase |
| FFA2 | free fatty acid receptor 2 |
| FFA3 | free fatty acid receptor s |
| 5-HT | 5-hydroxytriptamine |
| GLP-1 | glucagon-like peptide 1 |
| MQC | N-[2-methylphenyl]-[4-furan-3-yl]-2-methyl-5-oxo-1,4,5,6,7,8-hexahydroquinoline-3-carboxamide |
| NO | nitric oxide |
| PA1 | phenylacetamide 1 |
| PGE2 | prostaglandin E2 |
| PTX | pertussis toxin |
| SCFA | short-chain fatty acid |
| TTX | tetrodotoxin |
| VIP | vasoactive intestinal polypeptide |
References
- Gill, S.R.; Pop, M.; Deboy, R.T.; Eckburg, P.B.; Turnbaugh, P.J.; Samuel, B.S.; Gordon, J.I.; Relman, D.A.; Fraser-Liggett, C.M.; Nelson, K. Metagenomic analysis of the human distal gut microbiome. Science 2006, 312, 1355–1359. [Google Scholar] [CrossRef] [PubMed]
- Keely, S.; Kelly, C.J.; Weissmueller, T.; Burgess, A.; Wagner, B.D.; Robertson, C.E.; Harris, J.K.; Colagan, S.P. Activated fluid transport regulates bacterial-epithelial interactions and significantly shifts the murine colonic microbiome. Gut Microbes 2012, 3, 250–260. [Google Scholar] [CrossRef] [PubMed]
- Li, F. Taste perception: From the tongue to the testis. Mol. Hum. Reprod. 2013, 19, 346–360. [Google Scholar] [CrossRef] [PubMed]
- Henquin, J.-G. Do pancreatic β cells “Taste” nutrients to secrete insulin? Sci. Signal. 2012, 5, pe36. [Google Scholar] [CrossRef] [PubMed]
- Raybould, H.E. Does your gut taste? Sensory transduction in the gastrointestinal tract. News Physiol. Sci. 1998, 13, 275–280. [Google Scholar] [CrossRef] [PubMed]
- Dehkordi, O.; Rose, J.E.; Fatemi, M.; Allard, J.S.; Balan, K.V.; Young, J.K.; Fatima, S.; Millis, R.M.; Jayam-Trouth, A. Neuronal expression of bitter receptors and downstream signaling molecules in the rat brainstem. Brain Res. 2012, 1475, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, K.; Ishikawa, Y. Oral and extra-oral taste perception. Semin. Cell Dev. Biol. 2013, 24, 240–246. [Google Scholar] [CrossRef] [PubMed]
- Foster, S.R.; Porrello, E.R.; Purdue, B.; Chan, H.W.; Voigt, A.; Frenzel, S.; Hannan, R.D.; Moritz, K.M.; Simmons, D.G.; Molenaar, P.; et al. Expression, regulation and putative nutrient-sensing function of taste GPCRs in the heart. PLoS ONE 2013, 8, e64579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masubuchi, Y.; Nakagawa, Y.; Ma, J.; Sasaki, T.; Kitamura, T.; Yamamoto, Y.; Kurose, H.; Kojima, I.; Shibata, H. A novel regulatory function of sweet taste-sensing receptor in adipogenic differentiation of 3T3-L-1 cells. PLoS ONE 2013, 8, e54500. [Google Scholar] [CrossRef] [PubMed]
- Rozengurt, E. Taste receptors in the gastrointestinal tract. I. Bitter taste receptors and alpha-gustducin in the mammalian gut. Am. J. Physiol. 2006, 291, G171–G177. [Google Scholar]
- Rozenburt, E.; Sternini, C. taste receptor signaling in the mammalian gut. Curr. Opin. Pharmacol. 2007, 6, 557–562. [Google Scholar] [CrossRef] [PubMed]
- Sbarbati, A.; Bramanti, P.; Benati, D.; Merigo, F. The diffuse chemosensory system: Exploring the iceberg toward the definition of functional roles. Prog. Neurobiol. 2010, 91, 77–89. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.V.; Rozengurt, N.; Yang, M.; Young, S.H.; Sinnett-Smith, J.; Rozengurt, E. Expression of bitter taste receptors of the T2R family in the gastrointestinal tract and enteroendocrine STC-1 cells. Proc. Natl. Acad. Sci. USA 2002, 99, 2392–2397. [Google Scholar] [CrossRef] [PubMed]
- Mace, O.J.; Tehan, B.; Marshall, F. Pharmacology and Physiology of gastrointestinal enteroendocrine cells. Pharmacol. Res. Perspect. 2015, 3, e00155. [Google Scholar] [CrossRef] [PubMed]
- Sbarbati, A.; Osculati, F. Solitary chemosensory cells in mammals. Cell Tissues Organs 2003, 175, 51–55. [Google Scholar] [CrossRef] [PubMed]
- Sbarbati, A.; Osculati, F. The taste cell-related diffuse chemosensory system. Prog. Neurobiol. 2005, 75, 295–307. [Google Scholar] [CrossRef] [PubMed]
- Finger, T.E.; Bӧttger, B.; Hansen, A.; Anderson, K.T.; Alimohammadi, H.; Silver, W.L. Solitary chemoreceptor cells in the nasal cavity serve as sentinels of respiration. Proc. Natl. Acad. Sci. USA 2003, 100, 8981–8986. [Google Scholar] [CrossRef] [PubMed]
- Tizzano, M.; Gulbransen, B.D.; Vandenbeuch, A.; Clapp, T.R.; Herman, J.P.; Sibhatu, H.M.; Churchill, M.E.; Silver, W.L.; Kinnamon, S.C.; Finger, T.E. Nasal chemosensory cells use bitter taste signaling to detect irritants and bacterial signals. Proc. Natl. Acad. Sci. USA 2010, 107, 3210–3215. [Google Scholar] [CrossRef] [PubMed]
- Krasteva, G.; Canning, B.J.; Hartmann, P.; Veres, T.Z.; Papadakis, T.; Mühlffeld, C.; Schliecker, K.; Tallini, Y.N.; Braun, A.; Hackstein, H.; et al. Cholinergic chemosensory cells in the trachea regulate breathing. Proc. Natl. Acad. Sci. USA 2011, 108, 9478–9483. [Google Scholar] [CrossRef] [PubMed]
- Krasteva, G.; Canning, B.J.; Papadakis, T.; Kummer, W. Cholinergic brush cells in the trachea mediate respiratory responses to quorum sensing molecules. Life Sci. 2012, 91, 992–996. [Google Scholar] [CrossRef] [PubMed]
- Saunders, C.J.; Christensen, M.; Finger, T.E.; Tizzano, M. Cholinergic neurotransmission links solitary chemosensory cells to nasal inflammation. Proc. Natl. Acad. Sci. USA 2014, 111, 6075–6080. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.J.; Kofonow, J.M.; Rosen, P.L.; Siebert, A.P.; Chen, B.; Doghramji, L.; Xiong, G.; Adappa, N.D.; Palmer, J.N.; Kennedy, D.W.; et al. Bitter and sweet taste receptors regulate human upper respiratory innate immunity. J. Clin. Investig. 2014, 124, 1393–1405. [Google Scholar] [CrossRef] [PubMed]
- Hӧfer, D.; Püschel, B.; Drenckhahn, D. Taste receptor-like cells in the rat gut identified by expression of alpha-gustducin. Proc. Natl. Acad. Sci. USA 1996, 93, 6631–6634. [Google Scholar] [CrossRef]
- Dyer, J.; Daly, K.; Salmon, K.S.; Arora, D.K.; Kokrashvili, Z.; Margolskee, R.F.; Shirazi-Beechey, S.P. Intestinal glucose sensing and regulation of intestinal glucose absorption. Biochem. Soc. Trans. 2007, 35 Pt 5, 1191–1194. [Google Scholar] [CrossRef] [PubMed]
- Laffitte, A.; Neiers, F.; Briand, L. Functional roles of the sweet taste receptor in oral extraoral tissues. Curr. Opin. Clin. Nutr. Metab. Care 2014, 17, 379–385. [Google Scholar] [CrossRef] [PubMed]
- Habib, A.M.; Richards, P.; Cairns, L.S.; Rogers, G.J.; Bannon, C.A.; Parker, H.E.; Morley, T.C.; Yeo, G.S.; Reimann, F.; Gribble, F.M. Overlap of endocrine hormone expression in the mouse intestine revealed by transcriptional profiling and flow cytometry. Endocrinology 2012, 153, 3054–3065. [Google Scholar] [CrossRef] [PubMed]
- Christensen, L.W.; Kuhre, R.E.; Janus, C.; Svebdsen, B.; Holst, J.J. Vascular, but not luminal, activation of FFAR1 (GPR40) stimulates GLP-1 secretion from isolated perfused rat small intestine. Physiol. Rep. 2015, 3, e12551. [Google Scholar] [CrossRef] [PubMed]
- Sato, A. Tuft cells. Anat. Sci. Int. 2007, 82, 187–199. [Google Scholar] [CrossRef] [PubMed]
- Gerbe, F.; van Es, J.H.; Makrini, L.; Brulin, B.; Mellitzer, G.; Robine, S.; Romagnolo, B.; Shroyer, N.F.; Bourgaux, J.F.; Pignodel, C.; et al. Distinct ATOH1 and neurog3 requirements define tuft cells as a new secretory cell type in the intestinal epithelium. J. Cell Biol. 2011, 192, 767–780. [Google Scholar] [CrossRef] [PubMed]
- Ley, R.E.; Turnbaugh, P.J.; Klein, S.; Gordon, J.I. Microbial Ecology: Human gut microbiota associated with obesity. Nature 2006, 444, 1022–1023. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, F.; Tremaroli, V.; Nielsen, J.; Backhed, F. Assessing the human gut microbiota in metabolic diseases. Diabetes 2013, 62, 3341–3349. [Google Scholar] [CrossRef] [PubMed]
- Flint, H.J.; Scott, K.P.; Louis, P.; Duncan, S.H. The role of the gut microbiota in nutrition and health. Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 577–689. [Google Scholar] [CrossRef] [PubMed]
- Lagerstrom, M.C.; Schioth, H.B. Structural diversity of G protein-coupled receptors and significance for drug discovery. Nat. Rev. Drug Discov. 2008, 7, 339–357. [Google Scholar] [CrossRef] [PubMed]
- Stoddart, L.A.; Smith, N.J.; Miligan, G. International Union of Pharmacology. LXXI. Free fatty acid receptors FFA1, -2, and -3: Pharmacology and pathophysiological functions. Pharmacol. Rev. 2008, 60, 405–417. [Google Scholar] [CrossRef] [PubMed]
- Davenport, A.P.; Alexander, S.P.; Sharman, J.L.; Pawson, A.J.; Benson, H.E.; Monaghan, A.E.; Liew, W.C.; Mpamhanga, C.P.; Bonner, T.I.; Neubig, R.R.; et al. International Union of Basic and Clinical Pharmacology. LXXXVIII. G-protein coupled receptor list: Recommendations for New Pairings with Cognate Ligands. Pharmacol. Rev. 2013, 65, 967–986. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.J.; Goldsworthy, S.M.; Bares, A.A.; Eilert, M.M.; Tcheang, L.; Daniels, D.; Muir, A.I.; Wigglesworth, M.J.; Kinghorn, I.; Fraser, N.J.; et al. The orphan G protein-coupled receptor GPR41 and GPR43 are activated by propionate and other short chain carboxylic acids. J. Biol. Chem. 2003, 278, 11312–11319. [Google Scholar] [CrossRef] [PubMed]
- Le Poul, E.; Loison, C.; Struyf, S.; Springael, J.Y.; Lannoy, V.; Decobecq, M.E.; Brezillon, S.; Dupriez, V.; Vassart, G.; van Damme, J.; et al. Functional characterization of human receptors for short chain fatty acids and their role in polymorphonuclear cell activation. J. Biol. Chem. 2003, 278, 25481–25489. [Google Scholar] [CrossRef] [PubMed]
- Sawzdargo, M.; George, S.R.; Nguyen, T.; Xu, S.; Kolakowski, L.F.; O’Dowd, B.F. A cluster of four novel human g protein-coupled receptor genes occurring in close proximity to CD22 gene on chromosome 19q13.1. Biochem. Biophys. Res. Commun. 1997, 239, 543–547. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, N.E.; Kotarsky, K.; Owman, C.; Olde, B. Identification of a free fatty acid receptor, FFA2r, expressed on leukocytes and activated by short-chain fatty acids. Biochem. Biophys. Res. Commun. 2003, 303, 1047–1052. [Google Scholar] [CrossRef]
- Karaki, S.; Mitsui, R.; Haysshi, H.; Kato, I.; Sugiya, H.; Iwanaga, T.; Furness, J.B.; Kuwahara, A. Short-chain fatty acid receptor, GPR43, is expressed by enteroendocrine cells and mucosal mast cells in rat intestine. Cell Tissue Res. 2006, 324, 353–360. [Google Scholar] [CrossRef] [PubMed]
- Karaki, S.; Tazoe, H.; Hayashi, H.; Kashiwabara, H.; Tooyama, K.; Suzuki, Y.; Kuwahara, A. Expression of the short-chain fatty acid receptor, GPR43, in the human colon. J. Mol. Histol. 2008, 39, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Tazoe, H.; Otomo, Y.; Kaji, I.; Tanaka, R.; Karaki, S.I.; Kuwahara, A. Roles of short-chain fatty acids receptor, GPR41 and GPR43 on colonic functions. J. Physiol. Pharmacol. 2008, 59 (Suppl. 2), 251–262. [Google Scholar] [PubMed]
- Tazoe, H.; Otomo, Y.; Karaki, S.; Kato, I.; Fukami, Y.; Terasaki, M.; Kuwahara, A. Expression of short-chain fatty acid receptor GPR41 in human colon. Biomed. Res. 2009, 30, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Nøhr, M.K.; Egerod, K.L.; Christensen, S.H.; Gille, A.; Offermanns, S.; Schwartz, T.W.; Moller, M. Expression of the short chain fatty acid receptor GRP41/FFA3 in autonomic and somatic sensory ganglia. Neuroscience 2015, 290, 126–137. [Google Scholar] [CrossRef] [PubMed]
- Kaji, I.; Akiba, Y.; Konno, K.; Watanabe, M.; Kimura, S.; Iwanaga, T.; Kuri, A.; Iwamoto, K.I.; Kuwahara, A.; Kaunitz, J.D. neural FFA3 activation inversely regulates anion secretion evoked by nicotinic ACh receptor activation in rat. J. Physiol. 2016, 594, 3339–3352. [Google Scholar] [CrossRef] [PubMed]
- Natarajan, N.; Pluznick, J.L. Olfaction in the kidney: ’smelling’ gut microbial metabolites. Exp. Physiol. 2016, 101, 478–481. [Google Scholar] [CrossRef] [PubMed]
- Høverstat, T.; Bjorneklett, A.; Midtvedt, T.; Fausa, O.; Bohmer, T. Short-chain fatty acids in the proximal gastrointestinal tract of healthy subjects. Scand. J. Gastroenterol. 1984, 19, 1053–1058. [Google Scholar]
- Ovesen, L.; Bendtsen, F.; Tage-Jensen, U.; Pedersen, N.T.; Gram, B.R.; Rune, S.J. Intraluminal pH in the stomach, duodenum, and proximal jejunum in normal subjects and patients with exocrine pancreatic insufficiency. Gastroenterology 1986, 90, 958–962. [Google Scholar] [CrossRef]
- Rune, S.J.; Viskum, K. Duodenal pH values in normal controls and in patients with duodenal ulcer. Gut 1969, 10, 569–571. [Google Scholar] [CrossRef] [PubMed]
- Nedergaard, M.; Goldman, S.A.; Desai, S.; Pulsinelli, W.A. Acid-induced death in neurons and glia. J. Neurosci. 1991, 11, 2489–2497. [Google Scholar] [PubMed]
- Kivilaakso, E.; Kiviluoto, T. Intracellular pH in isolated ulcerogenic conditions. Gastroenterology 1988, 95, 1198–1205. [Google Scholar] [CrossRef]
- Illman, R.J.; Topping, D.L.; Trimble, R.P. Effects of food restriction and starvation-refeeding on volatile fatty acid concentrations in rats. J. Nutr. 1986, 116, 1694–1700. [Google Scholar] [CrossRef] [PubMed]
- Cummings, J.H.; Pomare, E.W.; Branch, W.J.; Naylor, C.P.; Macfarlance, G.T. Short chain fatty acids in human large intestine, portal, hepatic and venous blood. Gut 1987, 28, 1221–1227. [Google Scholar] [CrossRef] [PubMed]
- Akiba, Y.; Inoue, T.; Kaji, I.; Higashiyama, M.; Narimatsu, K.; Iwamoto, K.; Watanabe, M.; Guth, P.H.; Engel, E.; Kuwahara, A.; et al. Short-chain fatty acid sensing in rat duodenum. J. Physiol. 2015, 593, 585–599. [Google Scholar] [CrossRef] [PubMed]
- Allen, A.; Flemstrӧm, G. Gastroduodenal mucus bicarbonate barrier: Protection against acid and pepsin. Am. J. Physiol. 2005, 288, C1–C19. [Google Scholar] [CrossRef] [PubMed]
- Vattay, P.; Feil, W.; Klimesch, S.; Wenzl, E.; Starlinger, M.; Schiessel, R. Acid stimulated alkaline secretion in the rabbit duodenum is passive and correlates with mucosal damage. Gut 1988, 29, 284–290. [Google Scholar] [CrossRef] [PubMed]
- Tolhurst, G.; Heffron, H.; Lam, Y.S.; Parker, H.E.; Habib, A.M.; Diakogiannaki, E.; Cameron, J.; Grosse, J.; Reimann, F.; Gribble, F.M. Short-chain fatty acids stimulate glucagon-like peptide-1 secretion via G-protein-coupled receptor FFAR2. Diabetes 2012, 61, 364–371. [Google Scholar] [CrossRef] [PubMed]
- Akiba, Y.; Watanabe, C.; Mizumori, M.; Kaunitz, J.D. Luminal l-glutamate enhances duodenal mucosal defense mechanisms via multiple glutamate receptors in rats. Am. J. Physiol. 2009, 297, G781–G791. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.H.; Inoue, T.; Higashiyama, M.; Guth, P.H.; Engel, E.; Kaunitz, J.D.; Akiba, Y. Umami receptor activation increases duodenal bicarbonate secretion via glucagon-like peptide-2 release in rats. J. Pharmacol. Exp. Ther. 2011, 339, 464–473. [Google Scholar] [CrossRef] [PubMed]
- Ruppin, H.; Bar-Meir, S.; Soergel, K.H.; Wood, C.M.; Schmitt, M.G., Jr. Absorption of short-chain fatty acids by the colon. Gastroenterology 1980, 78, 1500–1507. [Google Scholar] [PubMed]
- Umesaki, Y.; Yajima, T.; Tohyama, K.; Mutai, M. Characterization of acetate uptake by the colonic epithelial cells of the rat. Pflugers Arch. 1980, 388, 205–209. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.D.; Delzenne, N.M. The gut microbiome as therapeutic target. Pharmacol. Ther. 2011, 130, 202–212. [Google Scholar] [CrossRef] [PubMed]
- Soldavini, J.; Kaunitz, J.D. Pathobiology and potential therapeutic value of intestinal short-chain fatty acids in gut inflammation and obesity. Dig. Dis. Sci. 2013, 58, 2756–2766. [Google Scholar] [CrossRef] [PubMed]
- Yajima, T. Contractile effect of short-chain fatty acids on the isolated colon of the rat. J. Physiol. 1985, 368, 667–678. [Google Scholar] [CrossRef] [PubMed]
- Karaki, S.I.; Kuwahara, A. Propionate-induced epithelial K+ and Cl−/HCO3− secretion and free fatty acid receptor 2 (FFA2, GPR43) expression in guinea-pig distal colon. Pflugers Arch. 2011, 461, 141–152. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Rhee, P.L.; Lee, J.H.; Kim, J.J.; Rhee, J.C.; Kim, S.J.; Lee, J. Segmental heterogeneity of electrogenic secretions in human ascending colon and rectum. Int. J. Colorectal Dis. 2006, 21, 357–364. [Google Scholar] [CrossRef] [PubMed]
- Kuwahara, A.; Radowicz-Cooke, H.J. Epithelial transport in guinea-pig proximal colon: Influence of enteric neurones. J. Physiol. 1988, 395, 271–284. [Google Scholar] [CrossRef] [PubMed]
- Kimura, I.; Inoue, D.; Maeda, T.; Hara, T.; Ichimura, A.; Miyauchi, S.; Kobayashi, M.; Hirasawa, A.; Tsujimoto, G. Short-chain fatty acids and ketones directly regulate sympathetic nervous system via G protein-coupled receptor 41 (GPR41). Proc. Natl. Acad. Sci. USA 2011, 108, 8030–8035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Won, Y.J.; Lu, V.B.; Puhl, H.L., 3rd; Ikeda, S.R. β-hydroxybutyrate modulates N-type calcium channels in rat sympathetic neurons by acting as an agonist for the G-protein-coupled receptor FFA3. J. Neurosci. 2013, 33, 19314–19325. [Google Scholar] [CrossRef] [PubMed]
- Berthoud, H.R.; Carlson, N.R.; Powley, T.L. Topography of efferent vagal innervation of the rat gastrointestinal tract. Am. J. Physiol. 1991, 260 Pt 2, R200–R207. [Google Scholar] [CrossRef] [PubMed]
- Palouzier, B.; Barrit-Chamoin, M.G.; Portalier, P.; Ternaux, J.P. Cholinergic neurons in the rat nodose ganglia. Neurosci. Lett. 1987, 80, 147–152. [Google Scholar] [CrossRef]
- Bellier, J.P.; Kimura, H. Acetylcholine synthesis by choline acetyltransferase of a peripheral type as demonstrated in adult rat dorsal root ganglion. J. Neurochem. 2007, 101, 1607–1618. [Google Scholar] [CrossRef] [PubMed]
- Diener, M.; Knobloch, S.F.; Bridges, R.J.; Keilmann, T.; Rummel, W. Cholinergic-mediated secretion in the rat colon: Neuronal and epithelial muscarinic responses. Eur. J. Pharmacol. 1989, 168, 777–793. [Google Scholar] [CrossRef]
- Yajima, T. Luminal propionate-induced secretory response in the rat distal colon in vitro. J. Physiol. 1988, 403, 559–575. [Google Scholar] [CrossRef] [PubMed]
- Mitsui, R.; Ono, S.; Karaki, S.; Kuwahara, A. Neural and non-neural mediation of propionate-induced contractile responses in the rat distal colon. Neurogastroenterol. Motil. 2005, 17, 585–594. [Google Scholar] [CrossRef] [PubMed]
- Yajima, T.; Inoue, R.; Matsumoto, M.; Yajima, M. Non-neuronal release of ACh plays a key role in secretory response to luminal propionate in rat colon. J. Physiol. 2011, 589 Pt 4, 953–962. [Google Scholar] [CrossRef] [PubMed]
- Sqires, P.E.; Rumsey, R.D.; Edwards, C.A.; Read, N.W. Effect of short-chain fatty acids on contractile activity and fluid flow in rat colon in vitro. Am. J. Physiol. 1992, 262 Pt 1, G813–G817. [Google Scholar] [CrossRef] [PubMed]
- Cherbut, C.; Ferrier, L.; Roze, C.; Anini, Y.; Blottiere, H.; Lecannu, G.; Galmiche, J.P. Short-chain fatty acids modify colonic motility through nerves and polypeptide YY release in the rat. Am. J. Physiol. 1998, 275 Pt 1, G1415–G1422. [Google Scholar] [CrossRef] [PubMed]
- Kaji, I.; Akiba, Y.; Said, H.; Narimatsu, K.; Kaunitz, J.D. Luminal 5-HT stimulates colonic bicarbonate secretion in rats. Br. J. Pharmacol. 2015, 172, 4655–4670. [Google Scholar] [CrossRef] [PubMed]
- Furness, J.B. The Enteric Nervous System; Blackwell Publishing: Oxford, UK, 2006; pp. 80–102. ISBN 978-1-4051-3376-0. [Google Scholar]
- Bergman, E.N. Energy contributions of volatile fatty acids from the gastrointestinal tract in various species. Physiol. Rev. 1990, 70, 567–590. [Google Scholar] [CrossRef] [PubMed]
- Iwanaga, T.; Kishimoto, A. Cellular distribution of monocarboxylate transporters: A review. Biomed. Res. 2015, 36, 279–301. [Google Scholar] [CrossRef] [PubMed]
- Samuel, B.S.; Shaito, A.; Motoike, T.; Rey, F.E.; Backhed, F.; Manchester, J.K.; Hammer, R.E.; Williams, S.C.; Crowley, J.; Yanagisawa, M.; et al. Effects of the gut microbiota on host adiposity are modulated by the short-chain fatty acid-binding G-protein-coupled receptor, Gpr41. Proc. Natl. Acad. Sci. USA 2008, 105, 16767–16772. [Google Scholar] [CrossRef] [PubMed]
- Hirota, C.L.; Mckay, D.M. Cholinergic regulation of epithelial ion transport in the mammalian intestine. Br. J. Pharmacol. 2006, 149, 463–479. [Google Scholar] [CrossRef] [PubMed]
- Hubel, K.A.; Russ, L. Mechanisms of the secretory response to luminal propionate in rat descending colon in vitro. J. Auton. Nerv. Syst. 1993, 43, 219–229. [Google Scholar] [CrossRef]
- O’Malley, K.E.; Farrell, C.B.; O’Boyle, K.M.; Baird, A.W. Cholinergic activation of Cl− secretion in rat colonic epithelia. Eur. J. Pharmacol. 1995, 275, 83–89. [Google Scholar] [CrossRef]
- Harberberger, R.; Schultheiss, G.; Dienner, M. Epithelial muscarinic M1 receptors contribute to carbachol-induced ion secretion in mouse colon. Eur. J. Pharmacol. 2006, 530, 229–233. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Cell Types | Gut Hormones | Gastric Body | Gastric Antrum | Small Intestine | Colon |
|---|---|---|---|---|---|
| EC | Serotonin | + | + | + | + |
| D | Somatostatin | + | + | + | + |
| ECL | Unknown | + | |||
| A | Glucagon | + (in fetus) | |||
| X = A-like | Ghrelin | + | |||
| G | Gastrin | + | + | ||
| M = I | CCK | + | |||
| S | Secretin | + | |||
| K | GIP | + | |||
| Mo | Motilin | + | |||
| N | Neurotensin | ||||
| L | GLP/PYY | + | + |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kuwahara, A.; Kuwahara, Y.; Inui, T.; Marunaka, Y. Regulation of Ion Transport in the Intestine by Free Fatty Acid Receptor 2 and 3: Possible Involvement of the Diffuse Chemosensory System. Int. J. Mol. Sci. 2018, 19, 735. https://doi.org/10.3390/ijms19030735
Kuwahara A, Kuwahara Y, Inui T, Marunaka Y. Regulation of Ion Transport in the Intestine by Free Fatty Acid Receptor 2 and 3: Possible Involvement of the Diffuse Chemosensory System. International Journal of Molecular Sciences. 2018; 19(3):735. https://doi.org/10.3390/ijms19030735
Chicago/Turabian StyleKuwahara, Atsukazu, Yuko Kuwahara, Toshio Inui, and Yoshinori Marunaka. 2018. "Regulation of Ion Transport in the Intestine by Free Fatty Acid Receptor 2 and 3: Possible Involvement of the Diffuse Chemosensory System" International Journal of Molecular Sciences 19, no. 3: 735. https://doi.org/10.3390/ijms19030735