SPAG17 Is Required for Male Germ Cell Differentiation and Fertility

 , and
, and
Abstract
:
1. Introduction
2. Results
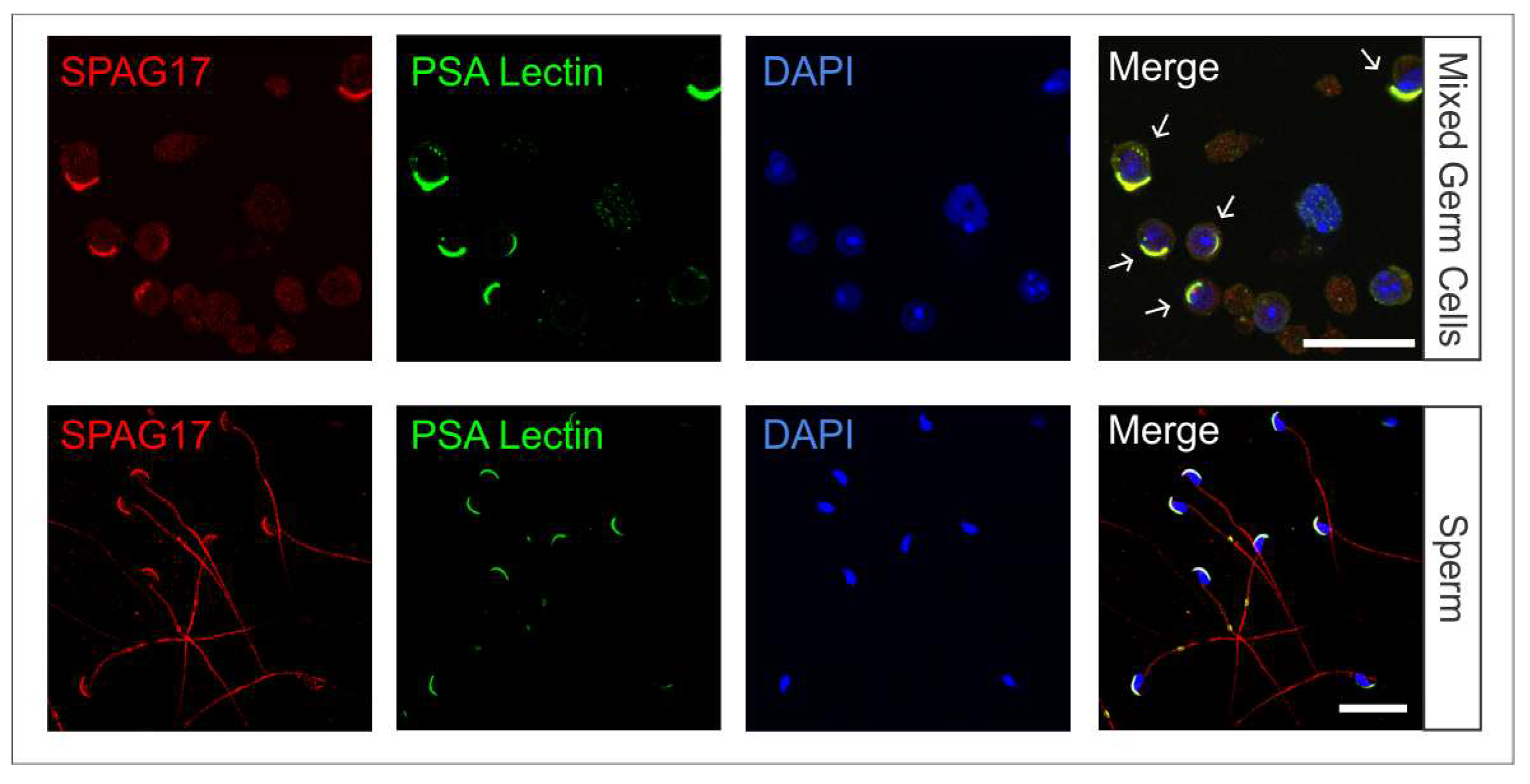
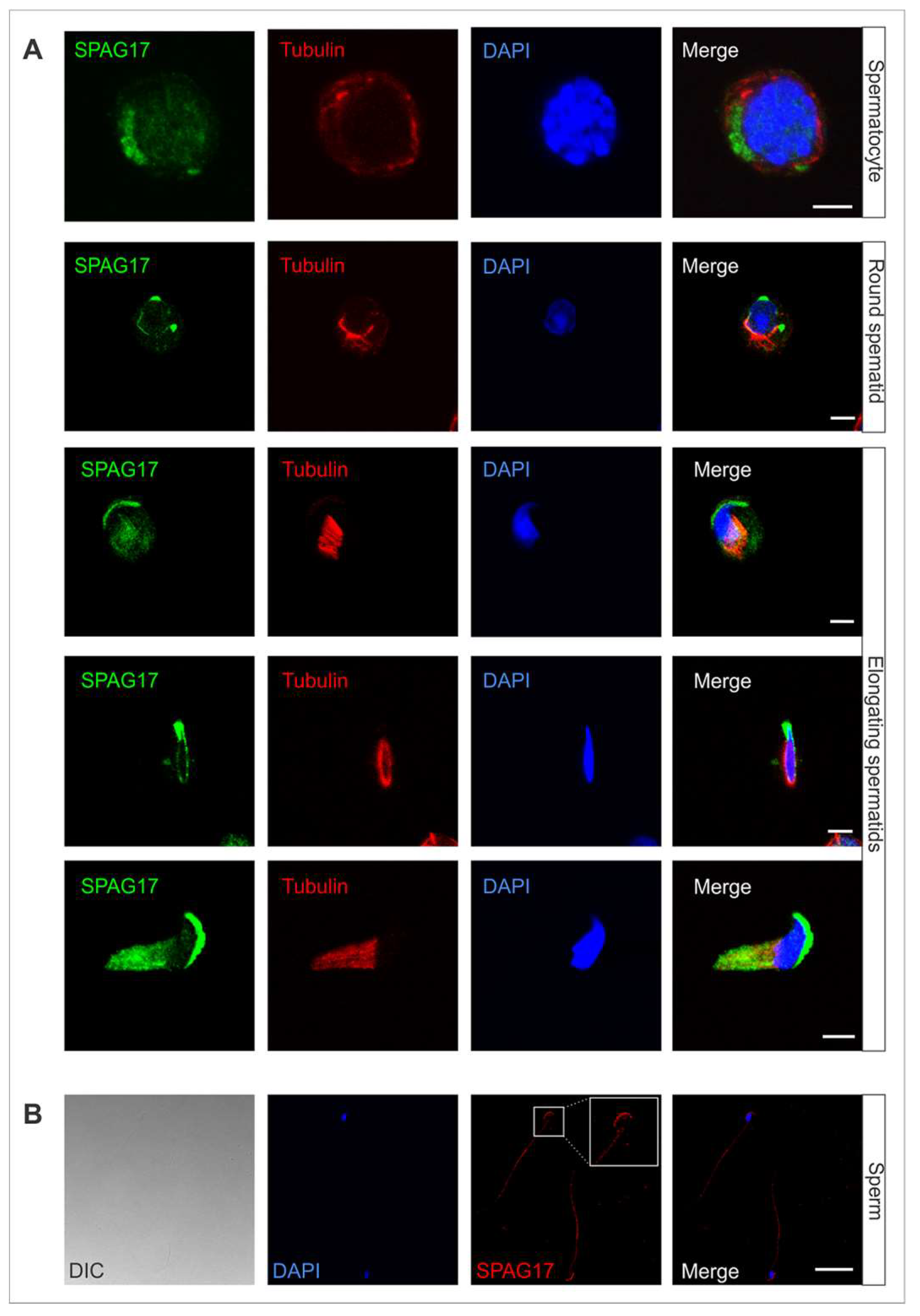
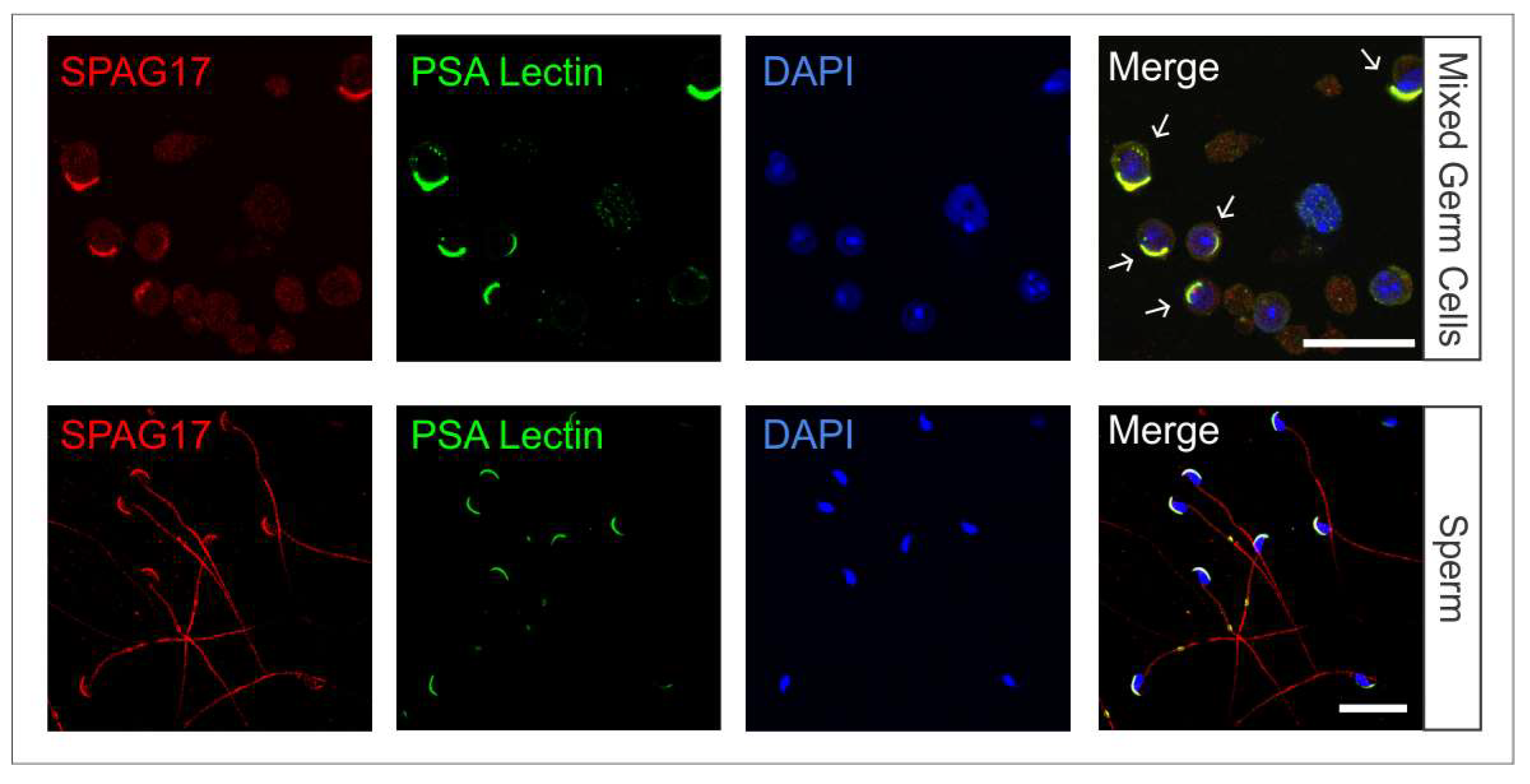
2.1. Localization of SPAG17 in Male Germ Cells
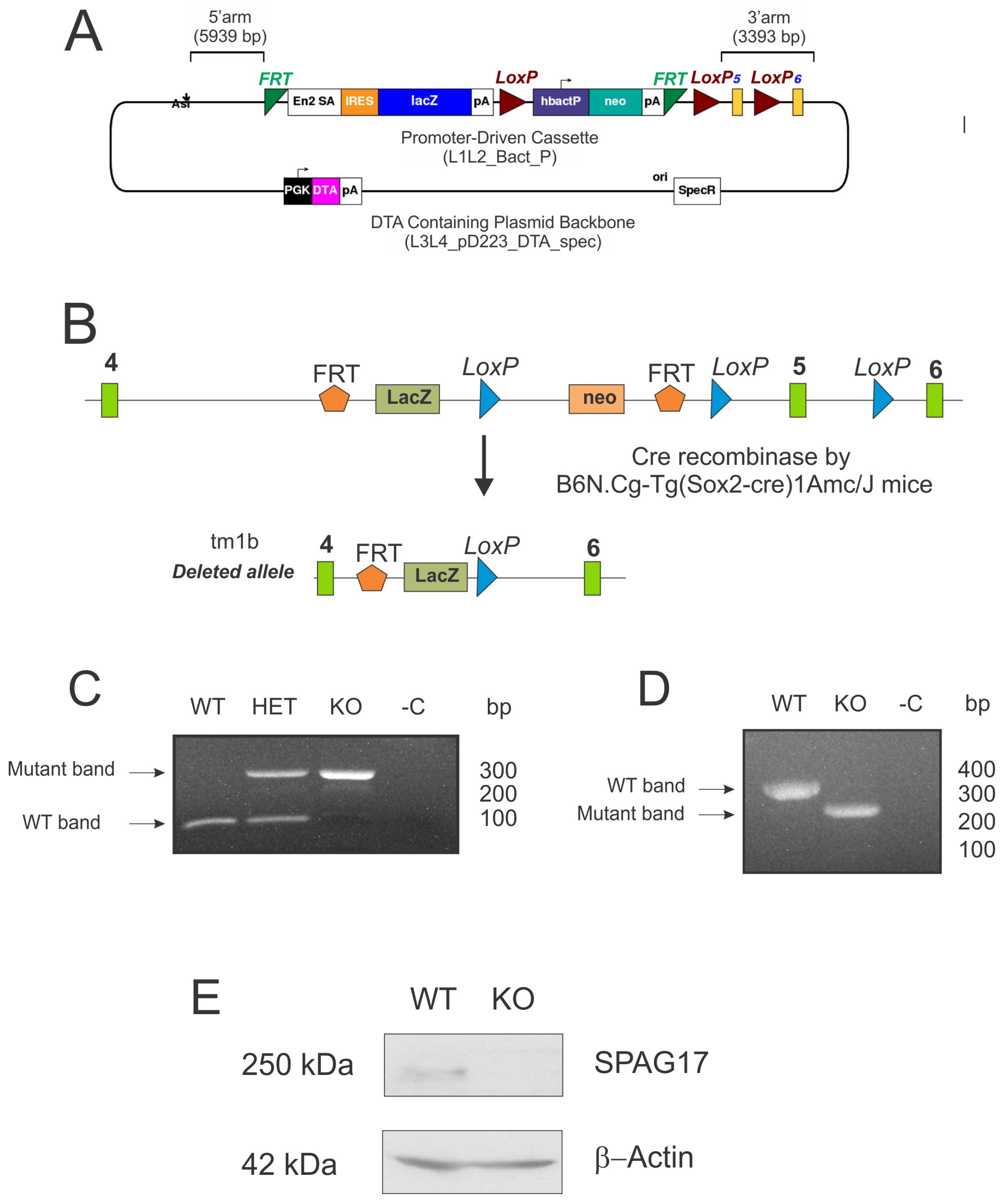
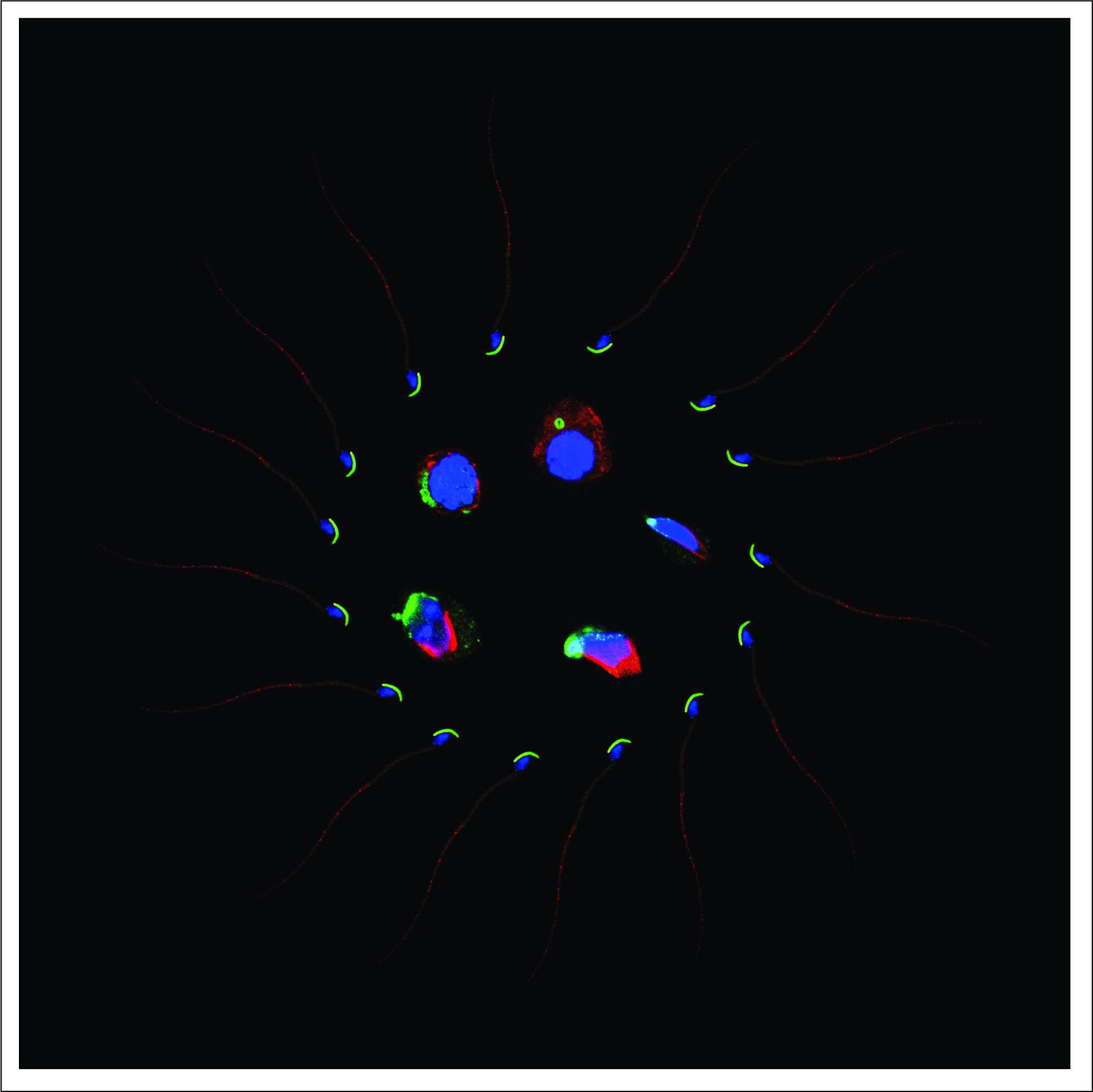
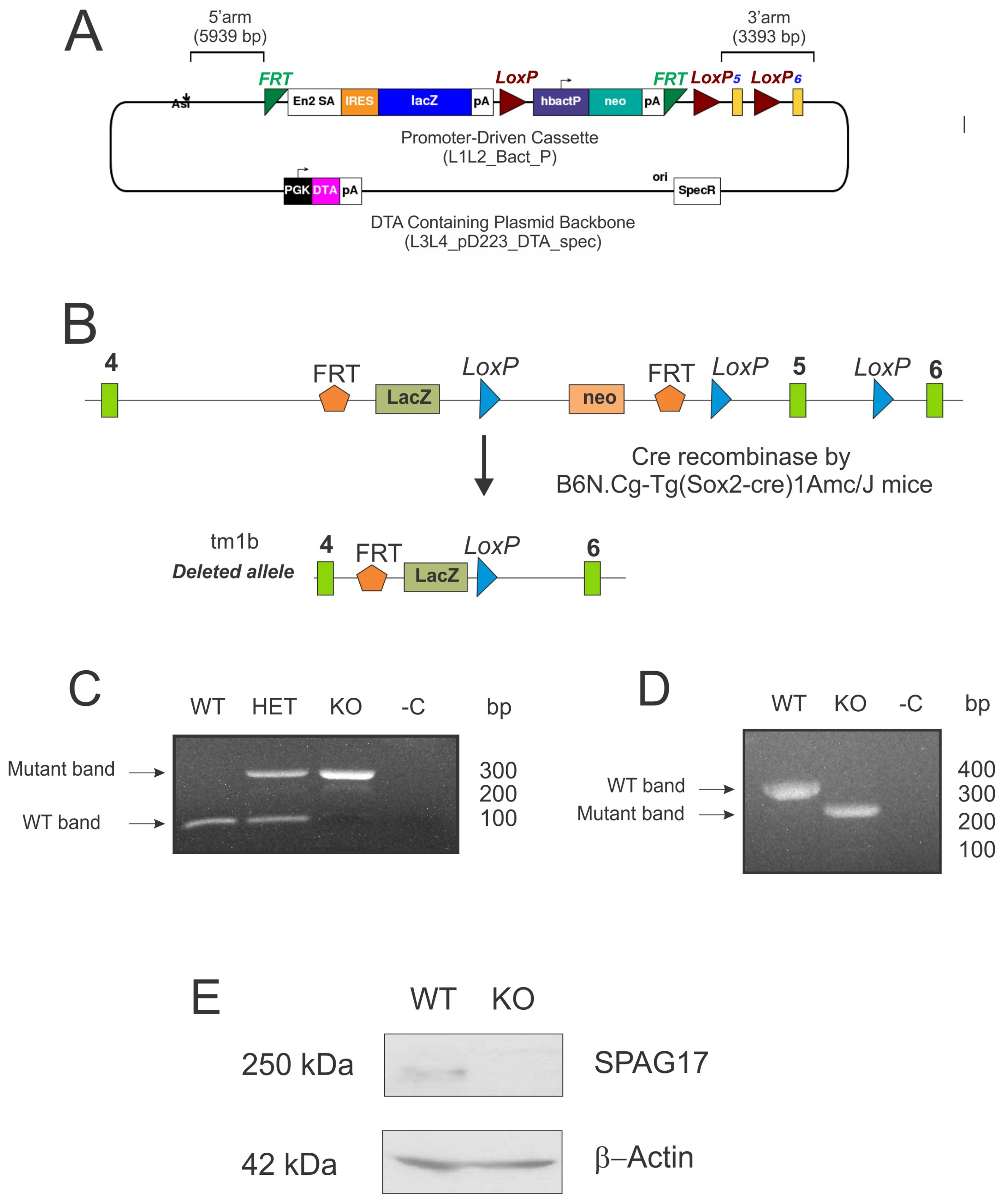
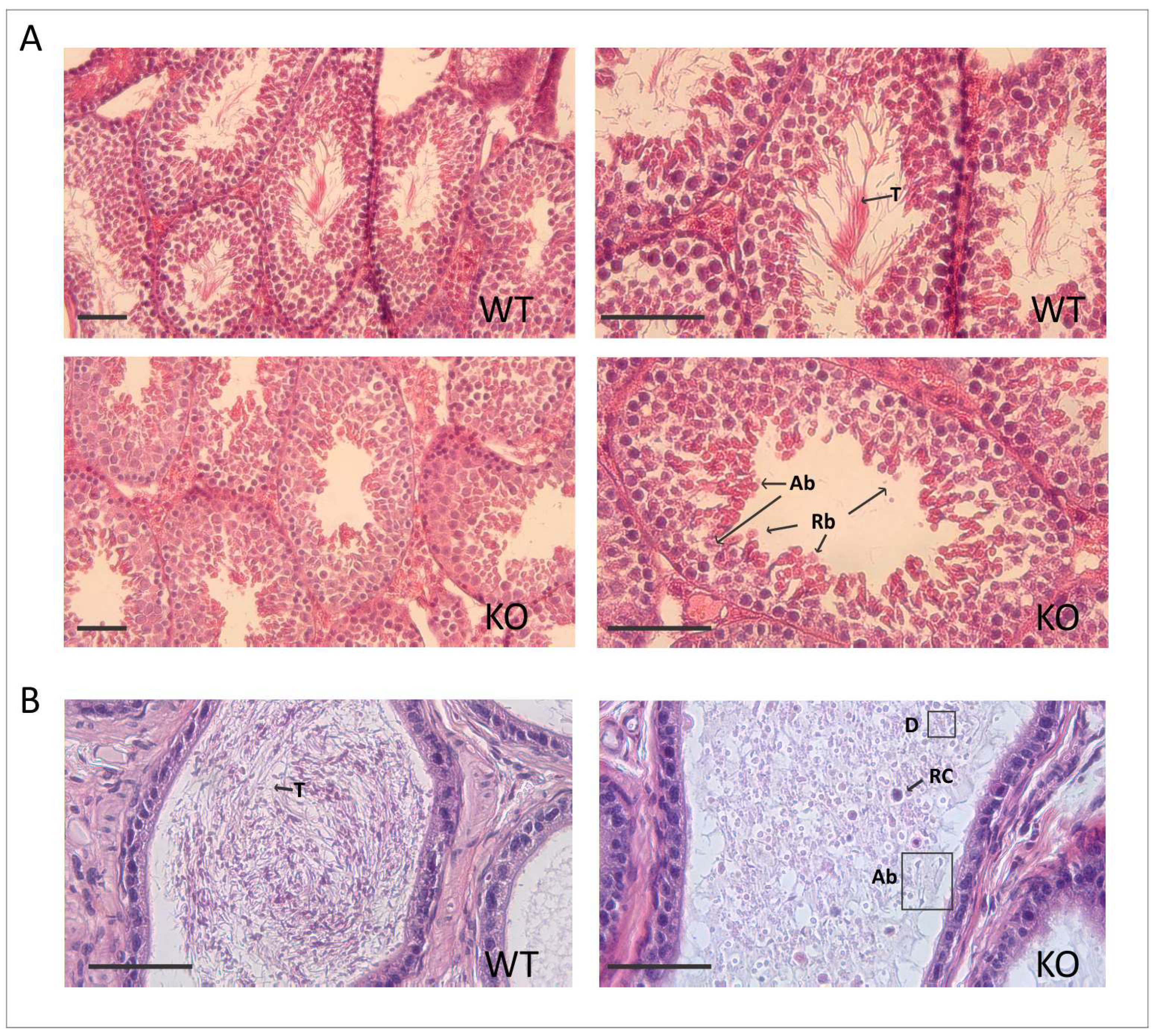
2.2. Deletion of Spag17 Causes Male Infertility
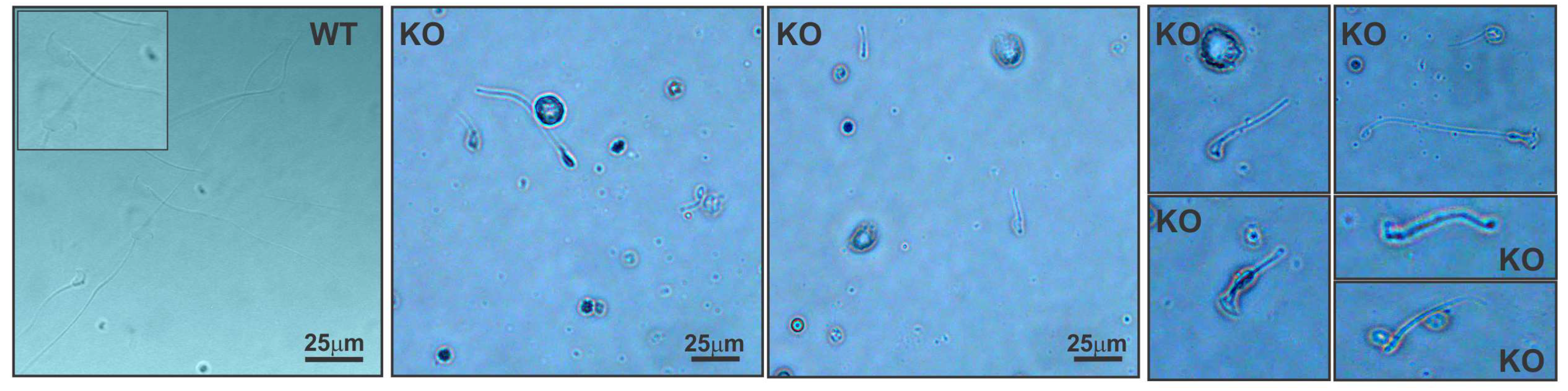
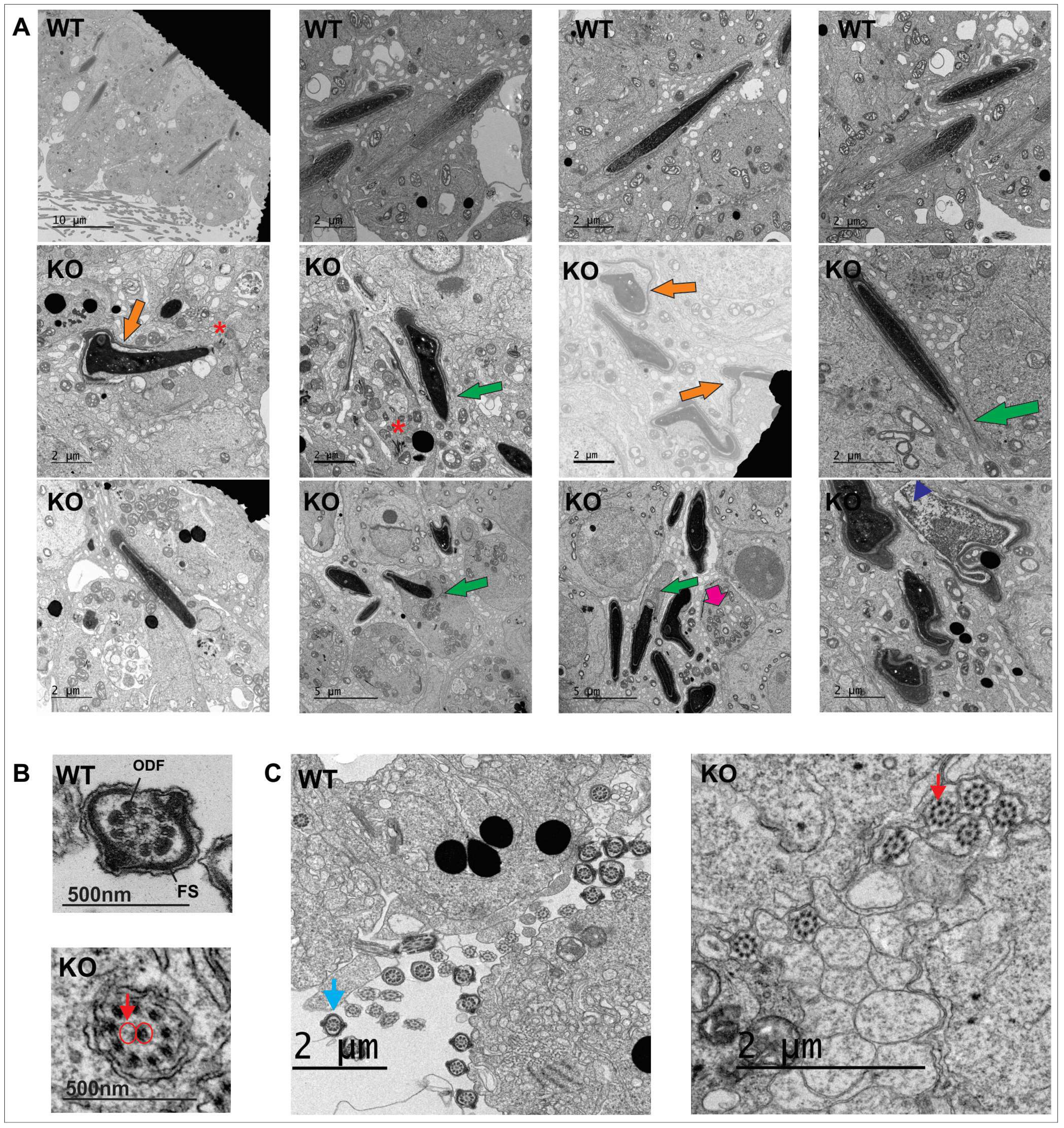
2.3. Spag17 Knockout Mice Display Defects in Manchette Structure and in the Morphology of the Sperm Head, Acrosome, and Tail
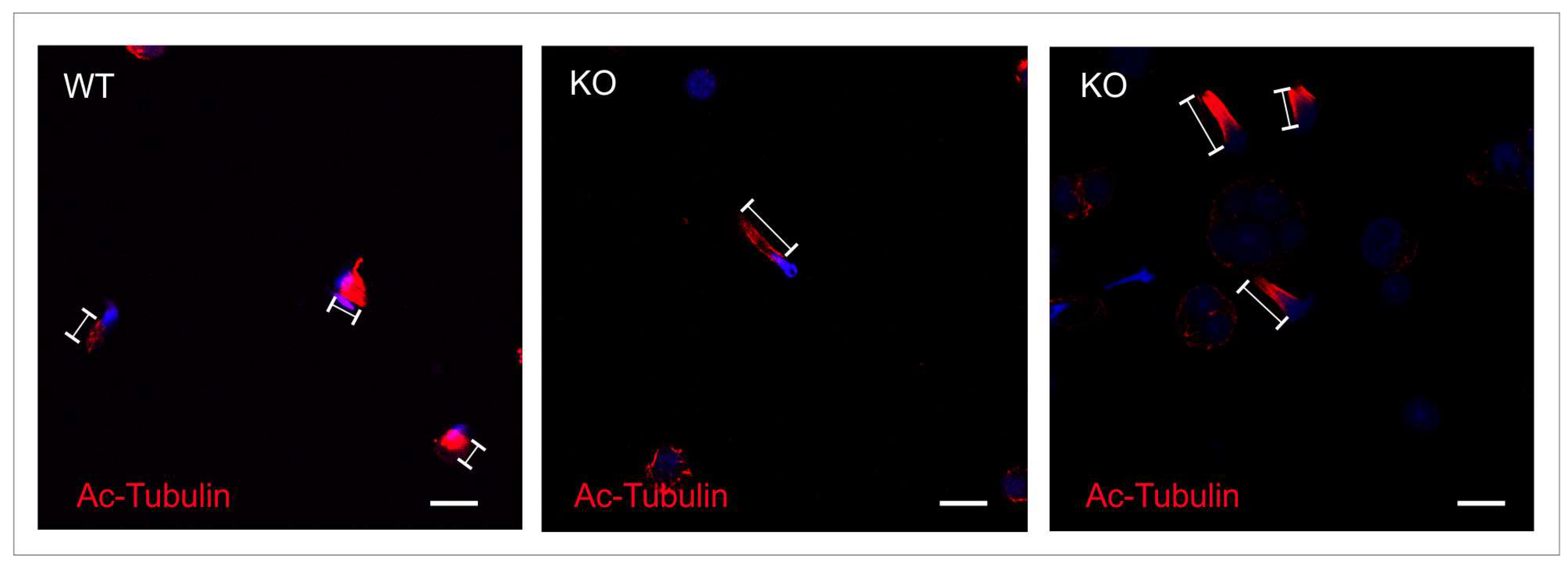
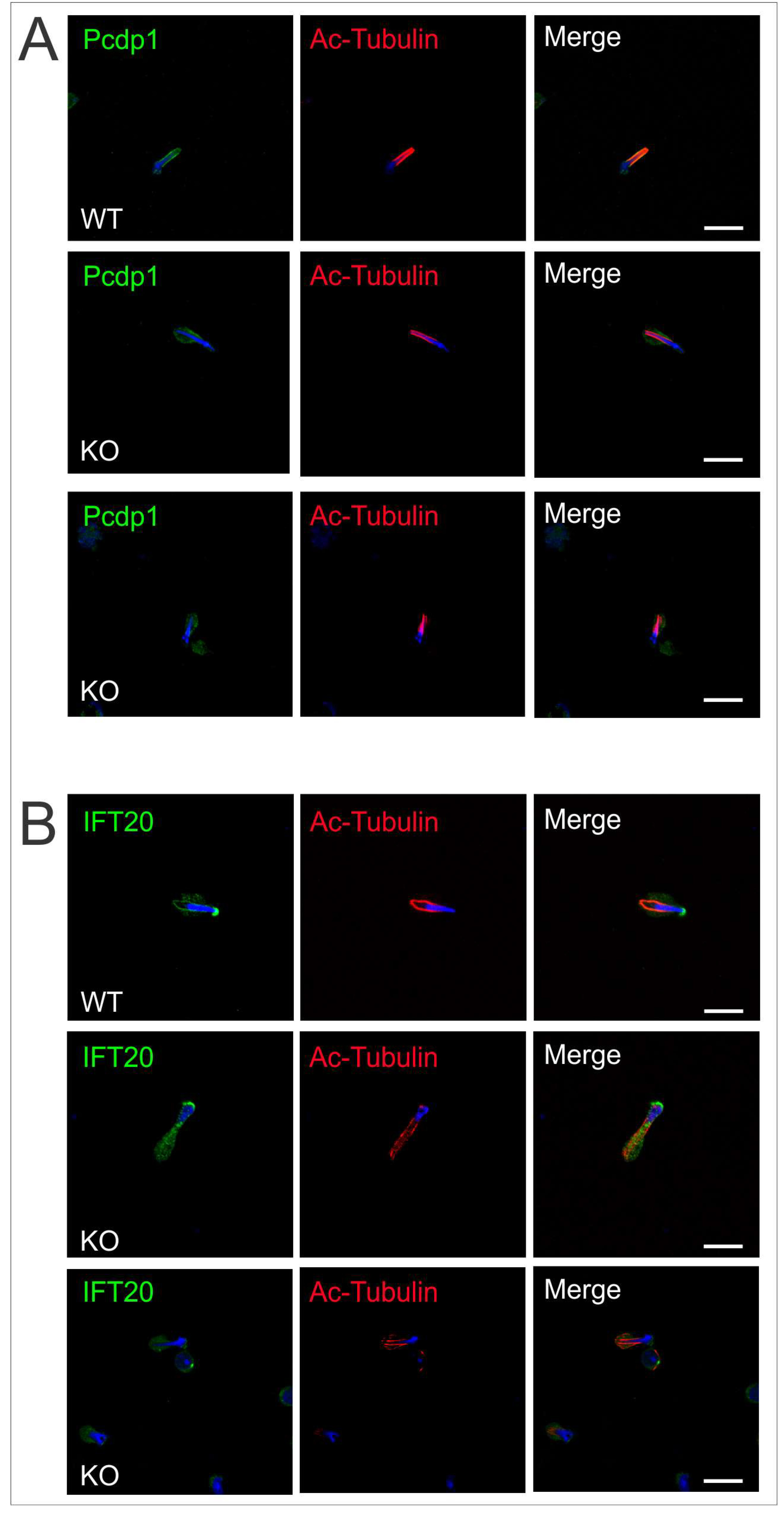
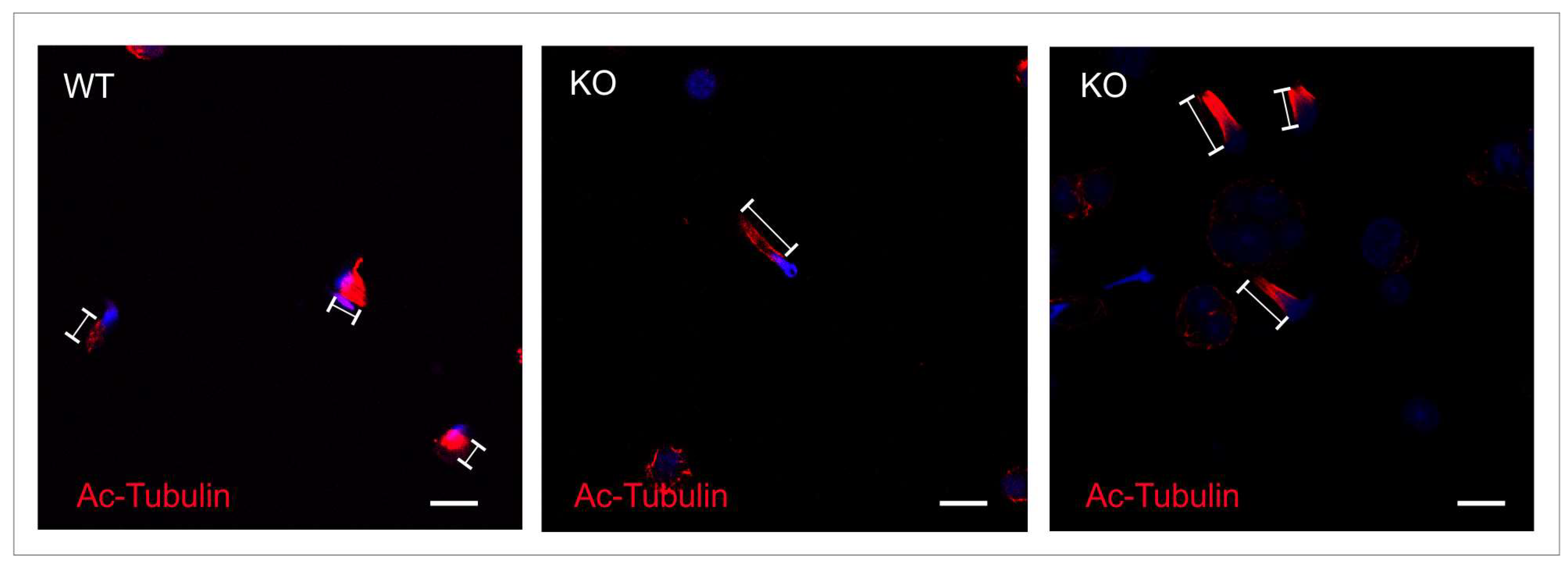
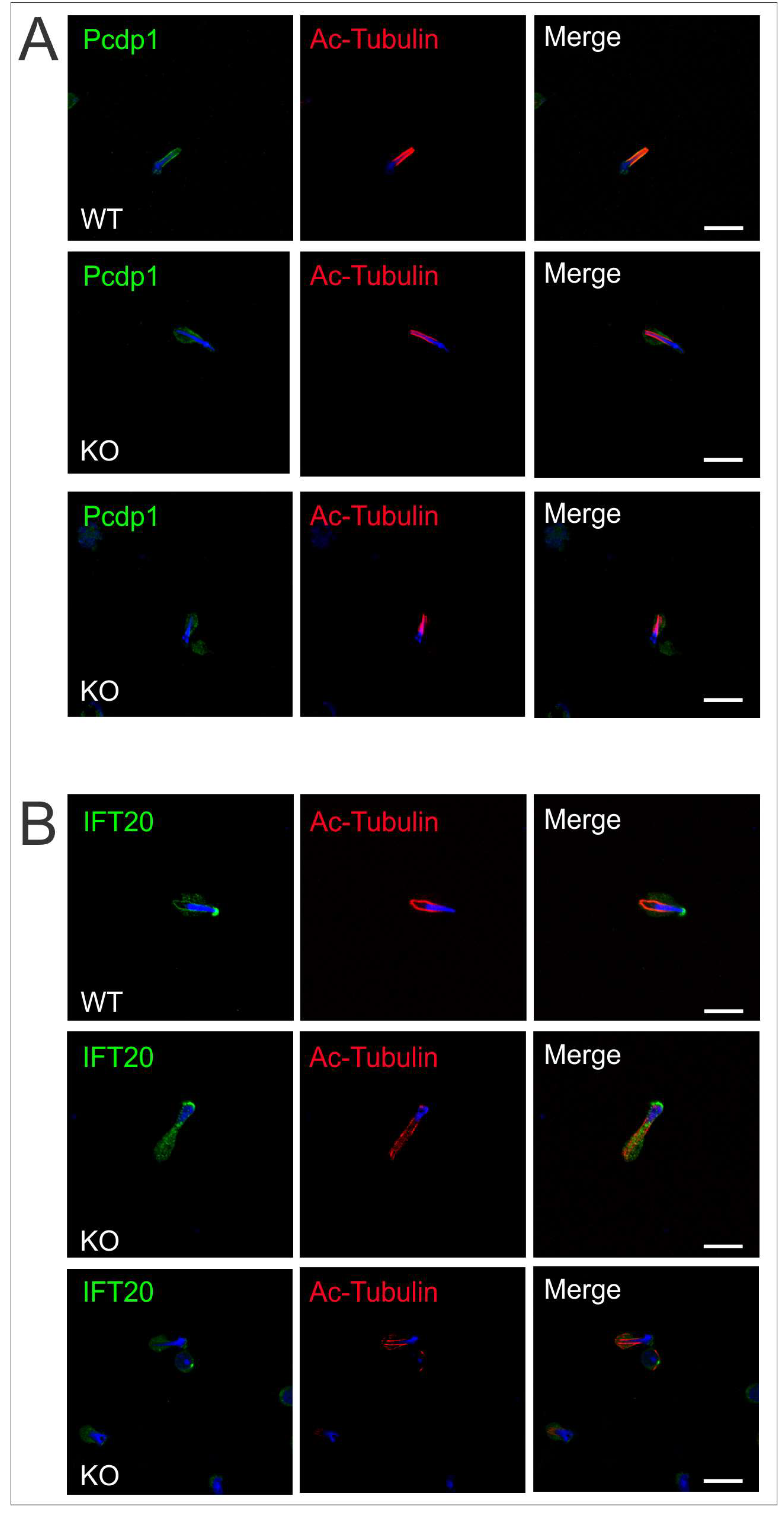
2.4. Intramanchette Transport Is Disrupted in Spag17 Knockout Mice
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Mixed Germ Cell Isolation
4.3. Sperm Isolation
4.4. Immunofluorescence
4.5. Mouse Genotyping
4.6. RT-PCR
4.7. Histology
4.8. Transmission Electron Microscopy (TEM)
Supplementary Material
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| IMT | Intramanchette transport |
| IFT | Intraflagellar transport |
| CPC | Central pair complex |
References
- Mitchell, D.; Smith, B. Analysis of the central pair microtubule complex in Chlamydomonas reinhardtii. Methods Cell Biol. 2009, 92, 197–213. [Google Scholar] [PubMed]
- Goduti, D.; Smith, E. Analyses of functional domains within the PF6 protein of the central apparatus reveal a role for PF6 sub-complex members in regulating flagellar beat frequency. Cytoskeleton 2012, 69, 179–194. [Google Scholar] [CrossRef] [PubMed]
- Rupp, G.; O’Toole, E.; Porter, M.E. The Chlamydomonas PF6 Locus Encodes a Large Alanine/Proline-Rich Polypeptide That Is Required for Assembly of a Central Pair Projection and Regulates Flagellar Motility. Mol. Biol. Cell 2001, 12, 739–751. [Google Scholar] [CrossRef] [PubMed]
- Wargo, M.J.; Dymek, E.E.; Smith, E.F. Calmodulin and PF6 are components of a complex that localizes to the C1 microtubule of the flagellar central apparatus. J. Cell Sci. 2005, 118, 4655–4665. [Google Scholar] [CrossRef] [PubMed]
- Teves, M.E.; Nagarkatti-Gude, D.R.; Zhang, Z.; Strauss, J.F. Mammalian axoneme central pair complex proteins: Broader roles revealed by gene knockout phenotypes. Cytoskeleton 2016, 73, 3–22. [Google Scholar] [CrossRef] [PubMed]
- Teves, M.E.; Zhang, Z.; Costanzo, R.M.; Henderson, S.C.; Corwin, F.D.; Zweit, J.; Sundaresan, G.; Subler, M.; Salloum, F.N.; Rubin, B.K.; et al. Sperm-associated antigen-17 gene is essential for motile cilia function and neonatal survival. Am. J. Respir Cell Mol. Biol. 2013, 48, 765–772. [Google Scholar] [CrossRef] [PubMed]
- Teves, M.E.; Sundaresan, G.; Cohen, D.J.; Hyzy, S.L.; Kajan, I.; Kajan, I.; Maczis, M.; Zhang, Z.; Costanzo, R.M.; Zweit, J.; et al. Spag17 Deficiency Results in Skeletal Malformations and Bone Abnormalities. PLoS ONE 2015, 10, e0125936. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Sha, Y.W.; Mei, L.B.; Ji, Z.Y.; Qiu, P.P.; Ji, H.; Li, P.; Wang, T.; Li, L. A familial study of twins with severe asthenozoospermia identified a homozygous SPAG17 mutation by whole-exome sequencing. Clin. Genet. 2017, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Meistrich, M.L.; Hess, R.A. Assessment of Spermatogenesis through Staging of Seminiferous Tubules. In Spermatogenesis: Methods and Protocols; Carrell, T.D., Aston, I.K., Eds.; Humana Press: Totowa, NJ, USA, 2013; pp. 299–307. [Google Scholar]
- Kierszenbaum, A.L.; Tres, L.L. The acrosome-acroplaxome-manchette complex and the shaping of the spermatid head. Arch. Histol. Cytol. 2004, 67, 271–284. [Google Scholar] [CrossRef] [PubMed]
- Lehti, M.S.; Sironen, A. Formation and function of the manchette and flagellum during spermatogenesis. Reproduction 2016, 151, R43–R54. [Google Scholar] [CrossRef] [PubMed]
- Horowitz, E.; Zhang, Z.; Jones, B.H.; Moss, S.B.; Ho, C.; Wood, J.R.; Wang, X.; Sammel, M.D.; Strauss, J.F., III. Patterns of expression of sperm flagellar genes: Early expression of genes encoding axonemal proteins during the spermatogenic cycle and shared features of promoters of genes encoding central apparatus proteins. Mol. Hum. Reprod. 2005, 11, 307–317. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Jones, B.H.; Tang, W.; Moss, S.B.; Wei, Z.; Ho, C.; Pollack, M.; Horowitz, E.; Bennett, J.; Baker, M.E.; et al. Dissecting the Axoneme Interactome: The Mammalian Orthologue of Chlamydomonas PF6 Interacts with Sperm-Associated Antigen 6, The Mammalian Orthologue of Chlamydomonas PF6. Mol. Cell. Proteom. 2005, 4, 914–923. [Google Scholar] [CrossRef] [PubMed]
- Western, P.; Maldonado-Saldivia, J.; van den Bergen, J.; Hajkova, P.; Saitou, M.; Barton, S.; Surani, M.A. Analysis of Esg1 Expression in Pluripotent Cells and the Germline Reveals Similarities with Oct4 and Sox2 and Differences between Human Pluripotent Cell Lines. Stem Cells 2005, 23, 1436–1442. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.-R.; Batool, A.; Wang, Y.-Q.; Hao, X.-X.; Chang, C.-S.; Cheng, C.Y.; Liu, Y.X. The control of male fertility by spermatid-specific factors: Searching for contraceptive targets from spermatozoon’s head to tail. Cell Death Dis. 2016, 7, e2472. [Google Scholar] [CrossRef] [PubMed]
- Lee, L.; Campagna, D.R.; Pinkus, J.L.; Mulhern, H.; Wyatt, T.A.; Sisson, J.H.; Pavlik, J.A.; Pinkus, G.S.; Fleming, M.D. Primary Ciliary Dyskinesia in Mice Lacking the Novel Ciliary Protein Pcdp1. Mol. Cell. Biol. 2008, 28, 949–957. [Google Scholar] [CrossRef] [PubMed]
- Kierszenbaum, A.L. Intramanchette transport (IMT): Managing the making of the spermatid head, centrosome, and tail. Mol. Reprod. Dev. 2002, 63, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Li, W.; Zhang, Y.; Zhang, L.; Teves, M.E.; Liu, H.; Strauss, J.F.; Pazour, G.J.; Foster, J.A.; Hess, R.A.; et al. Intraflagellar transport protein IFT20 is essential for male fertility and spermiogenesis in mice. Mol. Biol. Cell 2016, 27, 3705–3716. [Google Scholar] [CrossRef] [PubMed]
- Lehti, M.S.; Zhang, F.-P.; Kotaja, N.; Sironen, A. SPEF2 functions in microtubule-mediated transport in elongating spermatids to ensure proper male germ cell differentiation. Development 2017, 144, 2683–2693. [Google Scholar]
- Córdova-Fletes, C.; Becerra-Solano, L.E.; Rangel-Sosa, M.M.; Rivas-Estilla, A.M.; Alberto Galán-Huerta, K.; Ortiz-López, R.; Rojas-Martínez, A.; Juárez-Vázquez, C.I.; García-Ortiz, J.E. Uncommon runs of homozygosity disclose homozygous missense mutations in two ciliopathy-related genes (SPAG17 and WDR35) in a patient with multiple brain and skeletal anomalies. Eur. J. Med. Genet. 2017, 61, 161–167. [Google Scholar] [PubMed]
- Antony, D.; Nampoory, N.; Bacchelli, C.; Melhem, M.; Wu, K.; James, C.T.; Beales, P.L.; Hubank, M.; Thomas, D. Exome sequencing for the differential diagnosis of ciliary chondrodysplasias: Example of a WDR35 mutation case and review of the literature. Eur. J. Med. Genet. 2017, 60, 658–666. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotype | Cell Count (1 × 10 −6/mL) | Male Fertility | Litter Size |
|---|---|---|---|
| WT | 5.73 ± 0.79 | 6/6 | 8.5 ± 0.9 |
| KO | 0.01 ± 0.003 * | 0/6 | 0 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kazarian, E.; Son, H.; Sapao, P.; Li, W.; Zhang, Z.; Strauss III, J.F.; Teves, M.E. SPAG17 Is Required for Male Germ Cell Differentiation and Fertility. Int. J. Mol. Sci. 2018, 19, 1252. https://doi.org/10.3390/ijms19041252
Kazarian E, Son H, Sapao P, Li W, Zhang Z, Strauss III JF, Teves ME. SPAG17 Is Required for Male Germ Cell Differentiation and Fertility. International Journal of Molecular Sciences. 2018; 19(4):1252. https://doi.org/10.3390/ijms19041252
Chicago/Turabian StyleKazarian, Elizabeth, HyunYoung Son, Paulene Sapao, Wei Li, Zhibing Zhang, Jerome F. Strauss III, and Maria E. Teves. 2018. "SPAG17 Is Required for Male Germ Cell Differentiation and Fertility" International Journal of Molecular Sciences 19, no. 4: 1252. https://doi.org/10.3390/ijms19041252