Role of Bone Morphogenetic Protein 7 (BMP7) in the Modulation of Corneal Stromal and Epithelial Cell Functions

 ,
,
Abstract
:1. Introduction
2. Results
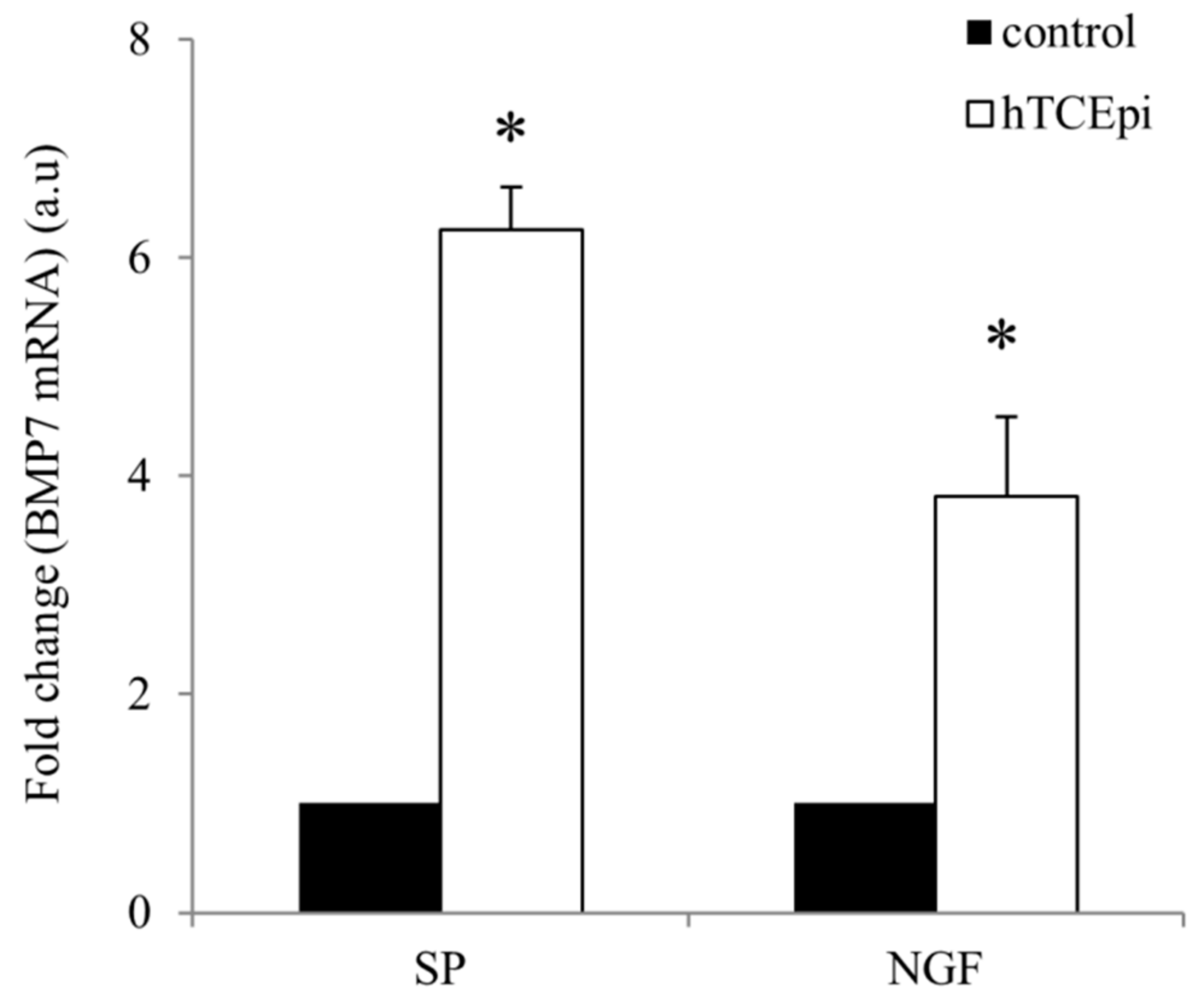
2.1. Elevation of BMP7 mRNA Expression after Treatment of Telomerase-Immortalized Human Corneal Epithelial (hTCEpi) Cells with Substance P (SP) and Nerve Growth Factor (NGF)
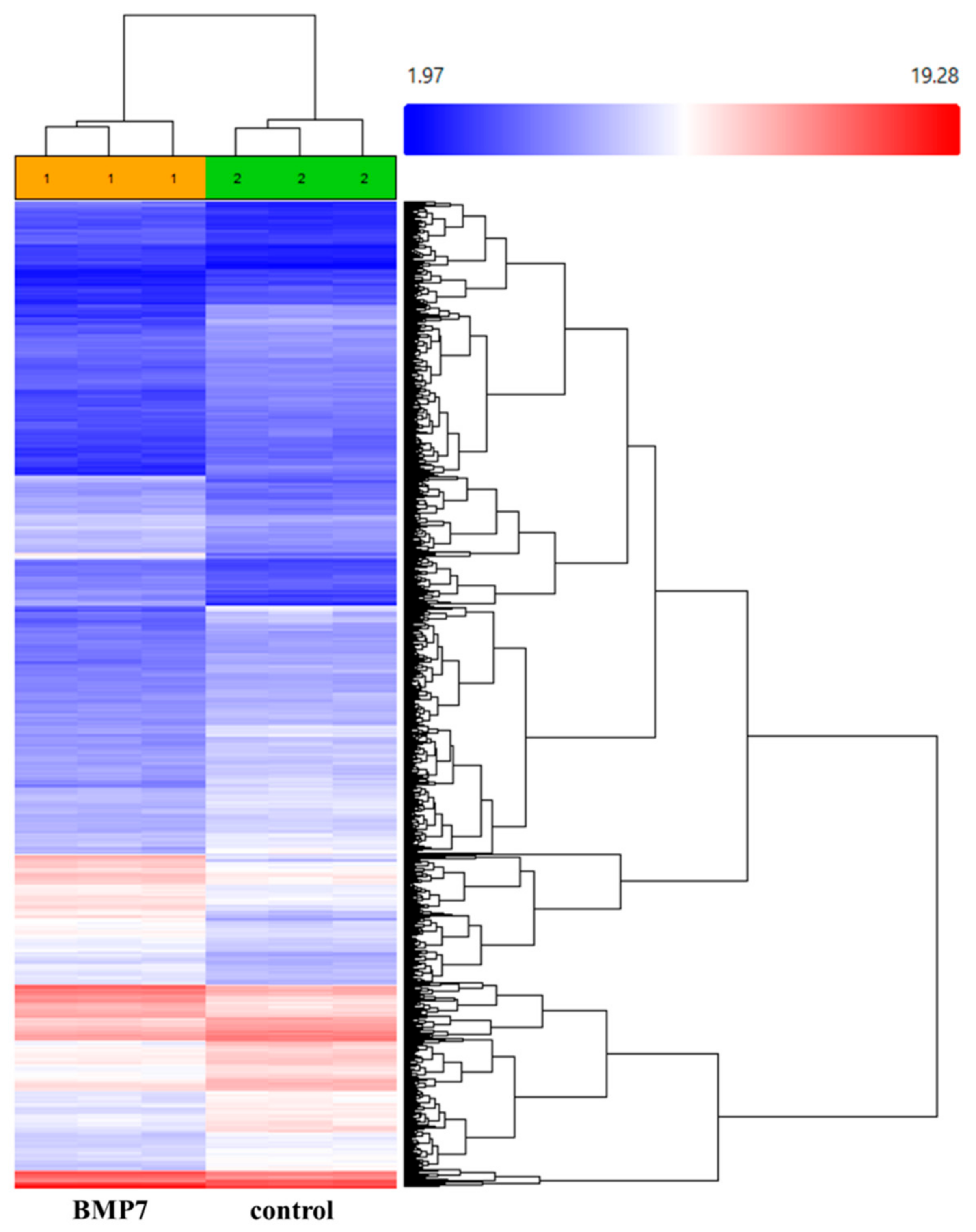
2.2. Gene Expression Profiling (GEP) of Recombinant Human BMP7 (rhBMP7) Stimulated hTCEpi Cells
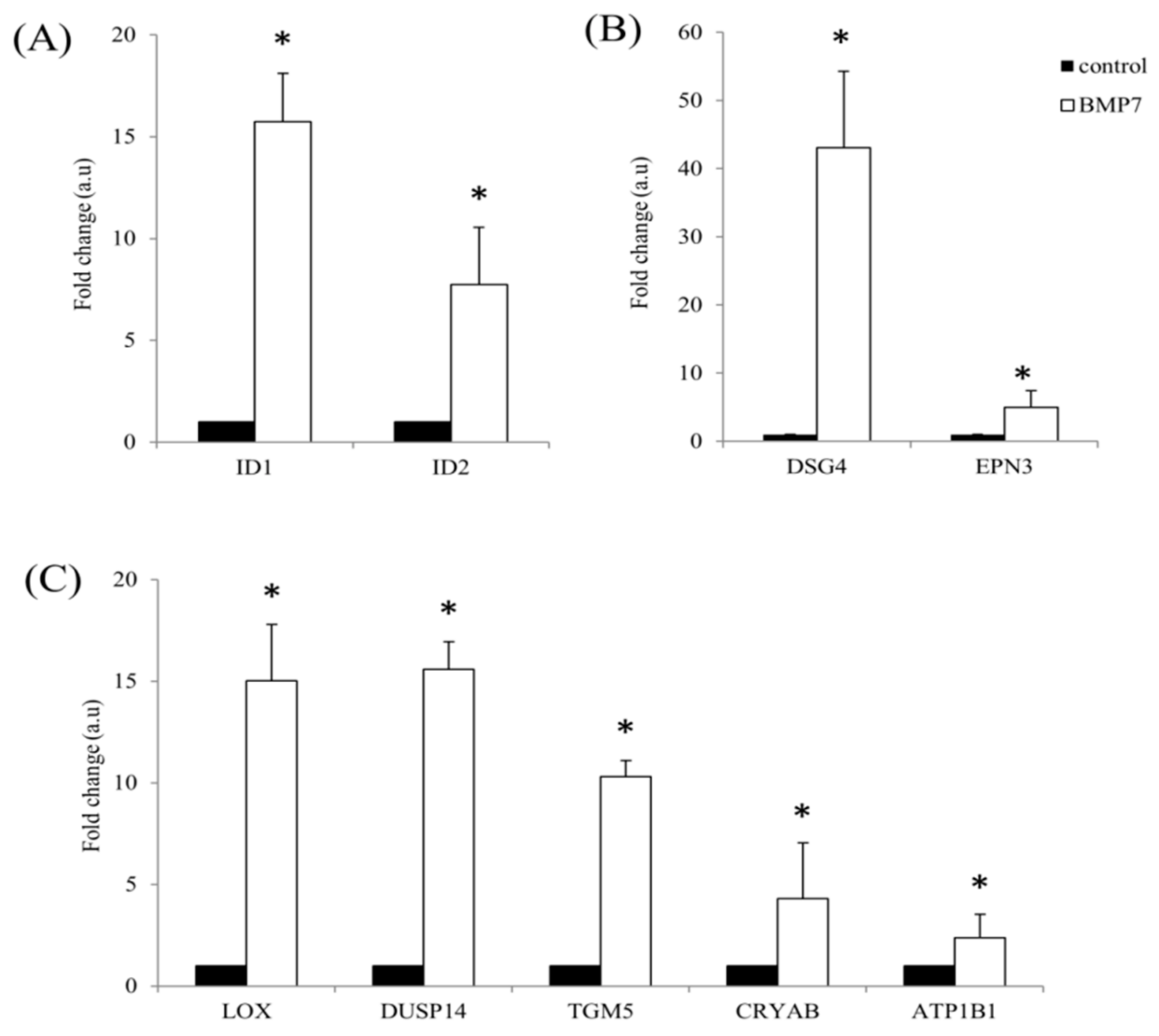
2.3. qRT-PCR Validation of the Transcriptome Analysis Data
2.4. Influence of rhBMP7 on the Migration of hTCEpi Cells
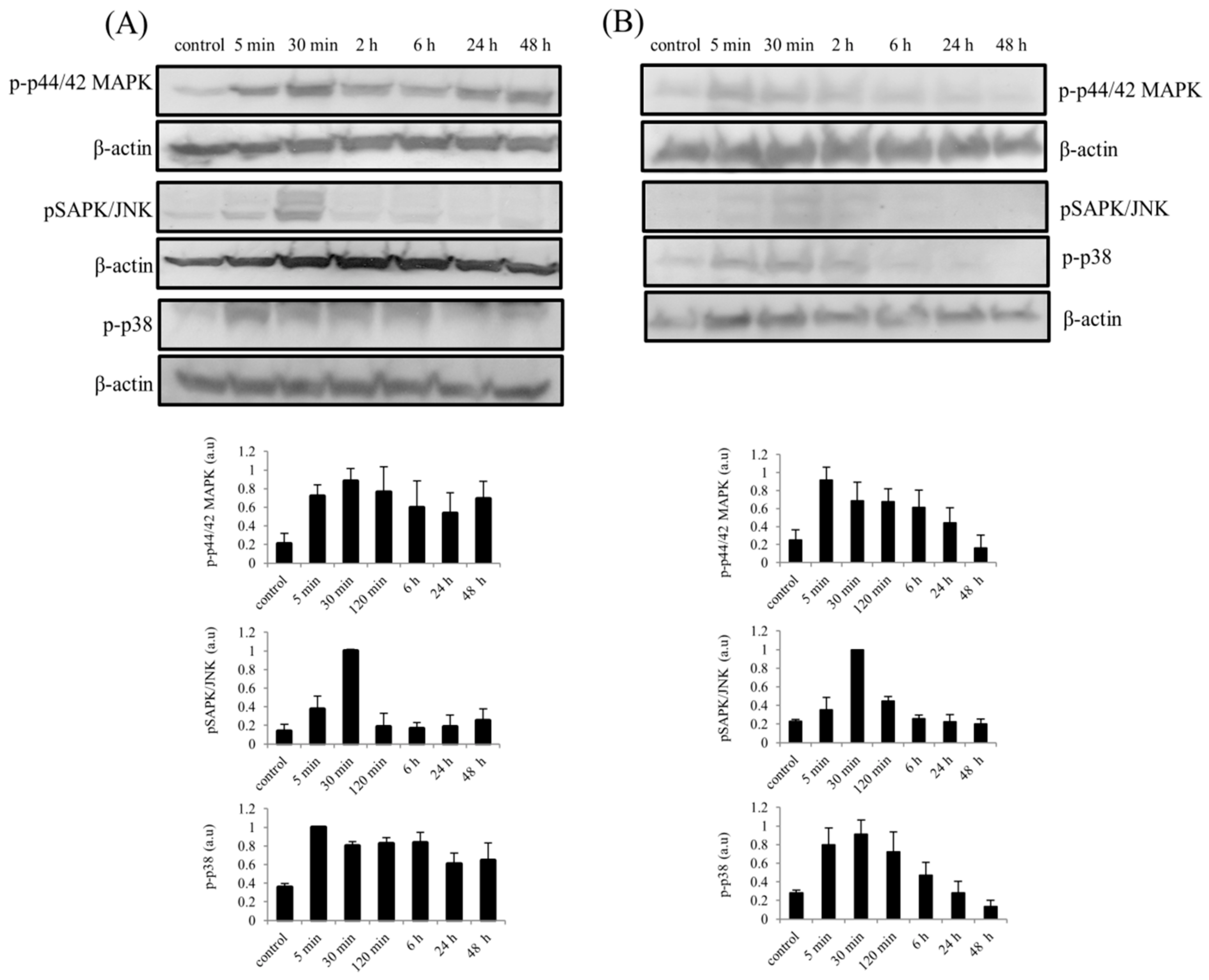
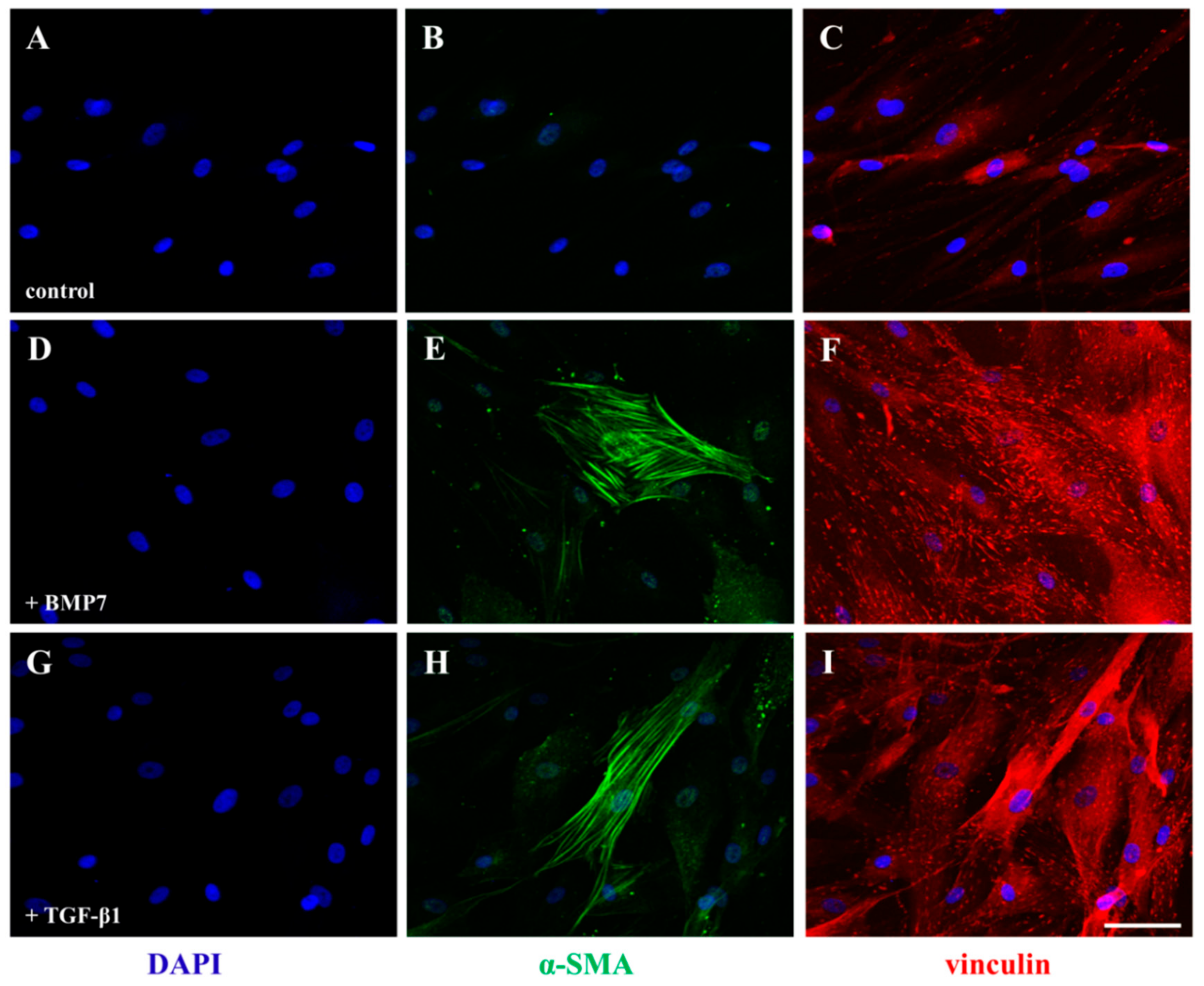
2.5. Stimulation of hTCEpi cells and Stromal Fibroblasts (SFs) by rhBMP7 Activates Vital Signaling Molecules
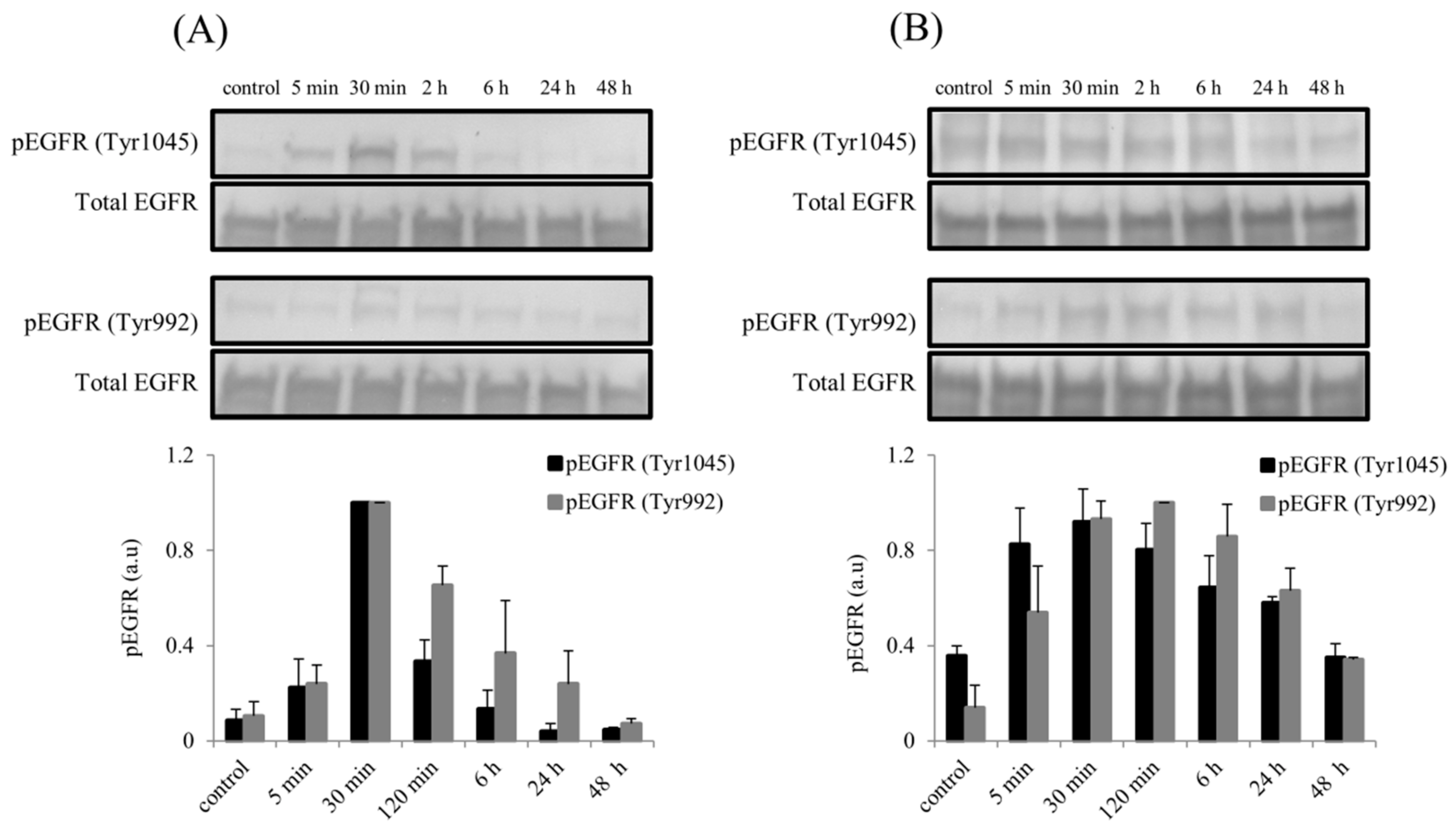
2.6. Elevated Phosphorylation of Epidermal Growth Factor Receptor (EGFR) Following rhBMP7 Stimulation
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Culture
4.3. Microarray Hybridization
4.4. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
4.5. Immunofluorescence
4.6. Immunoblotting
4.7. Scratch Assay
4.8. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| α-SMA | Alpha smooth muscle actin |
| ATP1β1 | ATPase, Na+/K+ transporting, beta 1 |
| BMP7 | Bone morphogenetic protein 7 |
| CRYAB | Crystallin alpha B |
| DSG4 | Desmoglein 4 |
| DUSP14 | Dual specificity phosphatase 14 |
| EGF | Epidermal growth factor |
| EGFR | Epidermal growth factor receptor |
| ERM | Ezrin/radixin/moesin |
| EPN3 | Epsin 3 |
| EMT | Epithelial-to-mesenchymal transition |
| ERK | Extracellular signal-regulated kinase |
| GEP | Gene expression profiling |
| hTCEpi | Telomerase-immortalized human corneal epithelial cell line |
| ID | Inhibitor of DNA-binding |
| LOX | Lysyl oxidase |
| MAPK | Mitogen-activated protein kinase |
| NGF | Nerve growth factor |
| SFs | Corneal stromal fibroblasts |
| SP | Substance P |
| TGF-β | Transforming growth factor beta |
| TGM5 | transglutaminase 5 |
| TNF | Tumor necrosis factor |
| ZEB1 | Zinc finger E-box-binding homeobox1 |
References
- Schultz, G.; Chegini, N.; Grant, M.; Khaw, P.; MacKay, S. Effects of growth factors on corneal wound healing. Acta Ophthalmol. Suppl. 1992, 60–66. [Google Scholar] [CrossRef]
- West-Mays, J.A.; Dwivedi, D.J. The keratocyte: Corneal stromal cell with variable repair phenotypes. Int. J. Biochem. Cell Biol. 2006, 38, 1625–1631. [Google Scholar] [CrossRef] [PubMed]
- Kowtharapu, B.S.; Murin, R.; Junemann, A.G.M.; Stachs, O. Role of corneal stromal cells on epithelial cell function during wound healing. Int. J. Mol. Sci. 2018, 19. [Google Scholar] [CrossRef] [PubMed]
- Imanishi, J.; Kamiyama, K.; Iguchi, I.; Kita, M.; Sotozono, C.; Kinoshita, S. Growth factors: Importance in wound healing and maintenance of transparency of the cornea. Prog. Retin. Eye Res. 2000, 19, 113–129. [Google Scholar] [CrossRef]
- Yu, F.S.; Yin, J.; Xu, K.; Huang, J. Growth factors and corneal epithelial wound healing. Brain Res. Bull. 2010, 81, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Yaraee, R.; Ghazanfari, T. Substance P potentiates TGF-β1 production in lung epithelial cell lines. Iran. J. Allergy Asthma Immunol. 2009, 8, 19–24. [Google Scholar] [PubMed]
- Hu, D.; Chen, B.; Zhu, X.; Tao, K.; Tang, C.; Wang, J. Substance P up-regulates the TGF-β1 mRNA expression of human dermal fibroblasts in vitro. Zhonghua Zheng Xing Wai Ke Za Zhi. 2002, 18, 234–236. [Google Scholar] [PubMed]
- Micera, A.; Lambiase, A.; Puxeddu, I.; Aloe, L.; Stampachiacchiere, B.; Levi-Schaffer, F.; Bonini, S.; Bonini, S. Nerve growth factor effect on human primary fibroblastic-keratocytes: Possible mechanism during corneal healing. Exp. Eye Res. 2006, 83, 747–757. [Google Scholar] [CrossRef] [PubMed]
- Saika, S. TGF-β signal transduction in corneal wound healing as a therapeutic target. Cornea 2004, 23, S25–S30. [Google Scholar] [CrossRef] [PubMed]
- Tandon, A.; Tovey, J.C.; Sharma, A.; Gupta, R.; Mohan, R.R. Role of transforming growth factor beta in corneal function, biology and pathology. Curr. Mol. Med. 2010, 10, 565–578. [Google Scholar] [PubMed]
- Bragdon, B.; Moseychuk, O.; Saldanha, S.; King, D.; Julian, J.; Nohe, A. Bone morphogenetic proteins: A critical review. Cell Signal 2011, 23, 609–620. [Google Scholar] [CrossRef] [PubMed]
- Wordinger, R.J.; Clark, A.F. Bone morphogenetic proteins and their receptors in the eye. Exp. Biol. Med. 2007, 232, 979–992. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Duffhues, G.; Hiepen, C.; Knaus, P.; Ten Dijke, P. Bone morphogenetic protein signaling in bone homeostasis. Bone 2015, 80, 43–59. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.N.; Green, J.; Wang, Z.; Deng, Y.; Qiao, M.; Peabody, M.; Zhang, Q.; Ye, J.; Yan, Z.; Denduluri, S.; et al. Bone morphogenetic protein (BMP) signaling in development and human diseases. Genes Dis. 2014, 1, 87–105. [Google Scholar] [CrossRef] [PubMed]
- Walsh, D.W.; Godson, C.; Brazil, D.P.; Martin, F. Extracellular BMP-antagonist regulation in development and disease: Tied up in knots. Trends Cell Biol. 2010, 20, 244–256. [Google Scholar] [CrossRef] [PubMed]
- Brazil, D.P.; Church, R.H.; Surae, S.; Godson, C.; Martin, F. BMP signalling: Agony and antagony in the family. Trends Cell Biol. 2015, 25, 249–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salazar, V.S.; Gamer, L.W.; Rosen, V. BMP signalling in skeletal development, disease and repair. Nat. Rev. Endocrinol. 2016, 12, 203–221. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Liu, Y.; Filas, B.; Gunhaga, L.; Beebe, D.C. Negative and positive auto-regulation of BMP expression in early eye development. Dev. Biol. 2015, 407, 256–264. [Google Scholar] [CrossRef] [PubMed]
- Kuribayashi, H.; Baba, Y.; Watanabe, S. BMP signaling participates in late phase differentiation of the retina, partly via upregulation of hey2. Dev. Neurobiol. 2014, 74, 1172–1183. [Google Scholar] [CrossRef] [PubMed]
- Faber, S.C.; Robinson, M.L.; Makarenkova, H.P.; Lang, R.A. BMP signaling is required for development of primary lens fiber cells. Development. 2002, 129, 3727–3737. [Google Scholar] [PubMed]
- Zhao, S.; Chen, Q.; Hung, F.C.; Overbeek, P.A. Bmp signaling is required for development of the ciliary body. Development. 2002, 129, 4435–4442. [Google Scholar] [PubMed]
- You, L.; Kruse, F.E.; Pohl, J.; Volcker, H.E. Bone morphogenetic proteins and growth and differentiation factors in the human cornea. Invest Ophthalmol Vis Sci. 1999, 40, 296–311. [Google Scholar] [PubMed]
- Mohan, R.R.; Kim, W.J.; Mohan, R.R.; Chen, L.; Wilson, S.E. Bone morphogenic proteins 2 and 4 and their receptors in the adult human cornea. Investig. Ophthalmol. Vis. Sci. 1998, 39, 2626–2636. [Google Scholar]
- Ljubimov, A.V.; Saghizadeh, M. Progress in corneal wound healing. Prog. Retin. Eye Res. 2015, 49, 17–45. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z. Transactivation of epidermal growth factor receptor by g protein-coupled receptors: Recent progress, challenges and future research. Int. J. Mol. Sci. 2016, 17. [Google Scholar] [CrossRef] [PubMed]
- Fischer, O.M.; Hart, S.; Gschwind, A.; Ullrich, A. EGFR signal transactivation in cancer cells. Biochem. Soc. Trans. 2003, 31, 1203–1208. [Google Scholar] [CrossRef] [PubMed]
- Forrester, S.J.; Kawai, T.; O’Brien, S.; Thomas, W.; Harris, R.C.; Eguchi, S. Epidermal growth factor receptor transactivation: Mechanisms, pathophysiology, and potential therapies in the cardiovascular system. Annu. Rev. Pharmacol. Toxicol. 2016, 56, 627–653. [Google Scholar] [CrossRef] [PubMed]
- McCarty, M.F. A role for cAMP-driven transactivation of egfr in cancer aggressiveness—Therapeutic implications. Med. Hypotheses 2014, 83, 142–147. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.P.; Dong, J.Z.; Xiong, L.J.; Shi, K.Q.; Zou, Z.L.; Zhang, S.N.; Cao, S.T.; Lin, Z.; Chen, Y.P. BMP-7 attenuates liver fibrosis via regulation of epidermal growth factor receptor. Int. J. Clin. Exp. Pathol. 2014, 7, 3537–3547. [Google Scholar] [PubMed]
- He, J.; Bazan, H.E. Epidermal growth factor synergism with TGF-β1 via PI-3 kinase activity in corneal keratocyte differentiation. Investig. Ophthalmol. Vis. Sci. 2008, 49, 2936–2945. [Google Scholar] [CrossRef] [PubMed]
- Lyu, J.; Lee, K.S.; Joo, C.K. Transactivation of EGFR mediates insulin-stimulated erk1/2 activation and enhanced cell migration in human corneal epithelial cells. Mol. Vis. 2006, 12, 1403–1410. [Google Scholar] [PubMed]
- Spix, J.K.; Chay, E.Y.; Block, E.R.; Klarlund, J.K. Hepatocyte growth factor induces epithelial cell motility through transactivation of the epidermal growth factor receptor. Exp. Cell Res. 2007, 313, 3319–3325. [Google Scholar] [CrossRef] [PubMed]
- Dudley, A.T.; Lyons, K.M.; Robertson, E.J. A requirement for bone morphogenetic protein-7 during development of the mammalian kidney and eye. Genes Dev. 1995, 9, 2795–2807. [Google Scholar] [CrossRef] [PubMed]
- Chung, J.K.; Park, S.A.; Hwang, H.S.; Kim, K.S.; Cho, Y.J.; You, Y.S.; Kim, Y.S.; Jang, J.W.; Lee, S.J. Effects of exogenous recombinant human bone morphogenic protein-7 on the corneal epithelial mesenchymal transition and fibrosis. Int. J. Ophthalmol. 2017, 10, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Saika, S.; Ikeda, K.; Yamanaka, O.; Flanders, K.C.; Nakajima, Y.; Miyamoto, T.; Ohnishi, Y.; Kao, W.W.; Muragaki, Y.; Ooshima, A. Therapeutic effects of adenoviral gene transfer of bone morphogenic protein-7 on a corneal alkali injury model in mice. Lab. Investig. 2005, 85, 474–486. [Google Scholar] [CrossRef] [PubMed]
- Tandon, A.; Sharma, A.; Rodier, J.T.; Klibanov, A.M.; Rieger, F.G.; Mohan, R.R. BMP7 gene transfer via gold nanoparticles into stroma inhibits corneal fibrosis in vivo. PLoS ONE 2013, 8, e66434. [Google Scholar] [CrossRef] [PubMed]
- Aomatsu, K.; Arao, T.; Sugioka, K.; Matsumoto, K.; Tamura, D.; Kudo, K.; Kaneda, H.; Tanaka, K.; Fujita, Y.; Shimomura, Y.; et al. TGF-β induces sustained upregulation of SNAI1 and SNAI2 through smad and non-smad pathways in a human corneal epithelial cell line. Investig. Ophthalmol. Vis. Sci. 2011, 52, 2437–2443. [Google Scholar] [CrossRef] [PubMed]
- Lim, M.; Chuong, C.M.; Roy-Burman, P. PI3K, ERK signaling in BMP7-induced epithelial-mesenchymal transition (EMT) of PC-3 prostate cancer cells in 2- and 3-dimensional cultures. Horm. Cancer 2011, 2, 298–309. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Fink, M.K.; Ghosh, A.; Tripathi, R.; Sinha, P.R.; Sharma, A.; Hesemann, N.P.; Chaurasia, S.S.; Giuliano, E.A.; Mohan, R.R. Novel combination BMP7 and HGF gene therapy instigates selective myofibroblast apoptosis and reduces corneal haze in vivo. Investig. Ophthalmol. Vis. Sci. 2018, 59, 1045–1057. [Google Scholar] [CrossRef] [PubMed]
- Kowtharapu, B.S.; Stahnke, T.; Wree, A.; Guthoff, R.F.; Stachs, O. Corneal epithelial and neuronal interactions: Role in wound healing. Exp. Eye Res. 2014, 125, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Segklia, A.; Seuntjens, E.; Elkouris, M.; Tsalavos, S.; Stappers, E.; Mitsiadis, T.A.; Huylebroeck, D.; Remboutsika, E.; Graf, D. BMP7 regulates the survival, proliferation, and neurogenic properties of neural progenitor cells during corticogenesis in the mouse. PLoS ONE 2012, 7, e34088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.N.; Lapage, J.; Hirschberg, R. Loss of tubular bone morphogenetic protein-7 in diabetic nephropathy. J. Am. Soc. Nephrol. 2001, 12, 2392–2399. [Google Scholar] [PubMed]
- Hata, A.; Lagna, G.; Massague, J.; Hemmati-Brivanlou, A. Smad6 inhibits BMP/Smad1 signaling by specifically competing with the smad4 tumor suppressor. Genes Dev. 1998, 12, 186–197. [Google Scholar] [CrossRef] [PubMed]
- Wilson, S.E.; Liu, J.J.; Mohan, R.R. Stromal-epithelial interactions in the cornea. Prog. Retin. Eye Res. 1999, 18, 293–309. [Google Scholar] [CrossRef]
- Wilson, S.E.; Netto, M.; Ambrosio, R., Jr. Corneal cells: Chatty in development, homeostasis, wound healing, and disease. Am. J. Ophthalmol. 2003, 136, 530–536. [Google Scholar] [CrossRef]
- Oswald, D.J.; Lee, A.; Trinidad, M.; Chi, C.; Ren, R.; Rich, C.B.; Trinkaus-Randall, V. Communication between corneal epithelial cells and trigeminal neurons is facilitated by purinergic (p2) and glutamatergic receptors. PLoS ONE 2012, 7, e44574. [Google Scholar] [CrossRef] [PubMed]
- Lambiase, A.; Manni, L.; Bonini, S.; Rama, P.; Micera, A.; Aloe, L. Nerve growth factor promotes corneal healing: Structural, biochemical, and molecular analyses of rat and human corneas. Investig. Ophthalmol. Vis. Sci. 2000, 41, 1063–1069. [Google Scholar]
- Kim, S.J.; Park, K.; Rudkin, B.B.; Dey, B.R.; Sporn, M.B.; Roberts, A.B. Nerve growth factor induces transcription of transforming growth factor-beta 1 through a specific promoter element in PC12 cells. J. Biol. Chem. 1994, 269, 3739–3744. [Google Scholar] [PubMed]
- Araki-Sasaki, K.; Aizawa, S.; Hiramoto, M.; Nakamura, M.; Iwase, O.; Nakata, K.; Sasaki, Y.; Mano, T.; Handa, H.; Tano, Y. Substance P-induced cadherin expression and its signal transduction in a cloned human corneal epithelial cell line. J. Cell Physiol. 2000, 182, 189–195. [Google Scholar] [CrossRef]
- Yang, L.; Sui, W.; Li, Y.; Qi, X.; Wang, Y.; Zhou, Q.; Gao, H. Substance P inhibits hyperosmotic stress-induced apoptosis in corneal epithelial cells through the mechanism of Akt activation and reactive oxygen species scavenging via the neurokinin-1 receptor. PLoS ONE 2016, 11, e0149865. [Google Scholar] [CrossRef] [PubMed]
- Blanco-Mezquita, T.; Martinez-Garcia, C.; Proenca, R.; Zieske, J.D.; Bonini, S.; Lambiase, A.; Merayo-Lloves, J. Nerve growth factor promotes corneal epithelial migration by enhancing expression of matrix metalloprotease-9. Investig. Ophthalmol. Vis. Sci. 2013, 54, 3880–3890. [Google Scholar] [CrossRef] [PubMed]
- Ortega, J.A.; Alcantara, S. BDNF/MAPK/ERK-induced BMP7 expression in the developing cerebral cortex induces premature radial glia differentiation and impairs neuronal migration. Cereb Cortex 2010, 20, 2132–2144. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.; Holtzman, D.A.; Chau, S.; Chickering, T.; Woolf, E.A.; Holmgren, L.M.; Bodorova, J.; Gearing, D.P.; Holmes, W.E.; Brivanlou, A.H. Twisted gastrulation can function as a BMP antagonist. Nature 2001, 410, 483–487. [Google Scholar] [CrossRef] [PubMed]
- Sotillo Rodriguez, J.E.; Mansky, K.C.; Jensen, E.D.; Carlson, A.E.; Schwarz, T.; Pham, L.; MacKenzie, B.; Prasad, H.; Rohrer, M.D.; Petryk, A.; Gopalakrishnan, R. Enhanced osteoclastogenesis causes osteopenia in twisted gastrulation-deficient mice through increased BMP signaling. J. Bone Miner. Res. 2009, 24, 1917–1926. [Google Scholar] [CrossRef] [PubMed]
- Phillips, M.A.; Qin, Q.; Hu, Q.; Zhao, B.; Rice, R.H. Arsenite suppression of BMP signaling in human keratinocytes. Toxicol. Appl. Pharmacol. 2013, 269, 290–296. [Google Scholar] [CrossRef] [PubMed]
- Miyazono, K.; Maeda, S.; Imamura, T. BMP receptor signaling: Transcriptional targets, regulation of signals, and signaling cross-talk. Cytokine Growth Factor Rev. 2005, 16, 251–263. [Google Scholar] [CrossRef] [PubMed]
- Miyazono, K.; Kamiya, Y.; Morikawa, M. Bone morphogenetic protein receptors and signal transduction. J. Biochem. 2010, 147, 35–51. [Google Scholar] [CrossRef] [PubMed]
- Spradling, K.D.; McDaniel, A.E.; Lohi, J.; Pilcher, B.K. Epsin 3 is a novel extracellular matrix-induced transcript specific to wounded epithelia. J. Biol. Chem. 2001, 276, 29257–29267. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.Y.; Chen, S.Z.; Zhang, W.T.; Wang, S.S.; Liu, Y.; Li, X.; Sun, X.; Li, Y.M.; Wen, B.; Lei, Q.Y.; et al. Induction of EMT-like response by BMP4 via up-regulation of lysyl oxidase is required for adipocyte lineage commitment. Stem Cell Res. 2013, 10, 278–287. [Google Scholar] [CrossRef] [PubMed]
- Kasashima, H.; Yashiro, M.; Kinoshita, H.; Fukuoka, T.; Morisaki, T.; Masuda, G.; Sakurai, K.; Kubo, N.; Ohira, M.; Hirakawa, K. Lysyl oxidase is associated with the epithelial-mesenchymal transition of gastric cancer cells in hypoxia. Gastric Cancer 2016, 19, 431–442. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhu, Y.; Tan, J.; Meng, X.; Xie, H.; Wang, R. Lysyl oxidase promotes epithelial-to-mesenchymal transition during paraquat-induced pulmonary fibrosis. Mol. Biosyst. 2016, 12, 499–507. [Google Scholar] [CrossRef] [PubMed]
- Chan, A.Y.; Bailly, M.; Zebda, N.; Segall, J.E.; Condeelis, J.S. Role of cofilin in epidermal growth factor-stimulated actin polymerization and lamellipod protrusion. J. Cell Biol. 2000, 148, 531–542. [Google Scholar] [CrossRef] [PubMed]
- Kiuchi, T.; Nagai, T.; Ohashi, K.; Mizuno, K. Measurements of spatiotemporal changes in G-actin concentration reveal its effect on stimulus-induced actin assembly and lamellipodium extension. J. Cell Biol. 2011, 193, 365–380. [Google Scholar] [CrossRef] [PubMed]
- Moriyama, K.; Iida, K.; Yahara, I. Phosphorylation of ser-3 of cofilin regulates its essential function on actin. Genes Cells 1996, 1, 73–86. [Google Scholar] [CrossRef] [PubMed]
- Kiuchi, T.; Ohashi, K.; Kurita, S.; Mizuno, K. Cofilin promotes stimulus-induced lamellipodium formation by generating an abundant supply of actin monomers. J. Cell Biol. 2007, 177, 465–476. [Google Scholar] [CrossRef] [PubMed]
- Louvet-Vallee, S. ERM proteins: From cellular architecture to cell signaling. Biol. Cell 2000, 92, 305–316. [Google Scholar] [CrossRef]
- Bosanquet, D.C.; Ye, L.; Harding, K.G.; Jiang, W.G. FERM family proteins and their importance in cellular movements and wound healing (review). Int. J. Mol. Med. 2014, 34, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Ng, T.; Parsons, M.; Hughes, W.E.; Monypenny, J.; Zicha, D.; Gautreau, A.; Arpin, M.; Gschmeissner, S.; Verveer, P.J.; Bastiaens, P.I.; et al. Ezrin is a downstream effector of trafficking PKC-integrin complexes involved in the control of cell motility. EMBO J. 2001, 20, 2723–2741. [Google Scholar] [CrossRef] [PubMed]
- Webb, P.G.; Spillman, M.A.; Baumgartner, H.K. Claudins play a role in normal and tumor cell motility. BMC Cell Biol. 2013, 14. [Google Scholar] [CrossRef] [PubMed]
- Ikari, A.; Takiguchi, A.; Atomi, K.; Sato, T.; Sugatani, J. Decrease in claudin-2 expression enhances cell migration in renal epithelial Madin-Darby canine kidney cells. J. Cell. Physiol. 2011, 226, 1471–1478. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.Y.; Guilford, P.; Thiery, J.P. Early events in cell adhesion and polarity during epithelial-mesenchymal transition. J. Cell. Sci. 2012, 125, 4417–4422. [Google Scholar] [CrossRef] [PubMed]
- Lamouille, S.; Xu, J.; Derynck, R. Molecular mechanisms of epithelial-mesenchymal transition. Nat. Rev. Mol. Cell Biol. 2014, 15, 178–196. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; El-Naggar, S.; Darling, D.S.; Higashi, Y.; Dean, D.C. Zeb1 links epithelial-mesenchymal transition and cellular senescence. Development 2008, 135, 579–588. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Zhao, Y.; Zhou, Y.; Xie, Y.; Ju, P.; Long, Y.; Liu, J.; Ni, D.; Cao, F.; Lyu, Z.; et al. Zeb1 is a potential regulator of Six2 in the proliferation, apoptosis and migration of metanephric mesenchyme cells. Int J. Mol. Sci. 2016, 17. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Lamouille, S.; Derynck, R. TGF-beta-induced epithelial to mesenchymal transition. Cell Res. 2009, 19, 156–172. [Google Scholar] [CrossRef] [PubMed]
- Peinado, H.; Olmeda, D.; Cano, A. Snail, Zeb and bHLH factors in tumour progression: An alliance against the epithelial phenotype? Nat. Rev. Cancer 2007, 7, 415–428. [Google Scholar] [CrossRef] [PubMed]
- Graham, T.R.; Zhau, H.E.; Odero-Marah, V.A.; Osunkoya, A.O.; Kimbro, K.S.; Tighiouart, M.; Liu, T.; Simons, J.W.; O’Regan, R.M. Insulin-like growth factor-i-dependent up-regulation of ZEB1 drives epithelial-to-mesenchymal transition in human prostate cancer cells. Cancer Res. 2008, 68, 2479–2488. [Google Scholar] [CrossRef] [PubMed]
- Barriere, G.; Fici, P.; Gallerani, G.; Fabbri, F.; Rigaud, M. Epithelial mesenchymal transition: A double-edged sword. Clin. Transl. Med. 2015, 4. [Google Scholar] [CrossRef] [PubMed]
- Terai, K.; Call, M.K.; Liu, H.; Saika, S.; Liu, C.Y.; Hayashi, Y.; Chikama, T.; Zhang, J.; Terai, N.; Kao, C.W.; et al. Crosstalk between TGF-β and MAPK signaling during corneal wound healing. Investig. Ophthalmol. Vis. Sci. 2011, 52, 8208–8215. [Google Scholar] [CrossRef] [PubMed]
- Okada, Y.; Saika, S.; Shirai, K.; Yamanaka, O.; Kitano, A.; Wang, Z.; Yang, H.; Reinach, P. JNK MAPK signaling contributes in vivo to injury-induced corneal epithelial migration. Ophthalmic Res. 2009, 42, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Zelenka, P.S.; Arpitha, P. Coordinating cell proliferation and migration in the lens and cornea. Semin Cell. Dev. Biol. 2008, 19, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Law, B.K.; Chytil, A.M.; Brown, K.A.; Aakre, M.E.; Moses, H.L. Activation of the ERK pathway is required for TGF-β1-induced EMT in vitro. Neoplasia 2004, 6, 603–610. [Google Scholar] [CrossRef] [PubMed]
- Blank, U.; Brown, A.; Adams, D.C.; Karolak, M.J.; Oxburgh, L. BMP7 promotes proliferation of nephron progenitor cells via a JNK-dependent mechanism. Development 2009, 136, 3557–3566. [Google Scholar] [CrossRef] [PubMed]
- Carreira, A.C.; Alves, G.G.; Zambuzzi, W.F.; Sogayar, M.C.; Granjeiro, J.M. Bone morphogenetic proteins: Structure, biological function and therapeutic applications. Arch. Biochem. Biophys. 2014, 561, 64–73. [Google Scholar] [CrossRef] [PubMed]
- Zeisberg, M.; Hanai, J.; Sugimoto, H.; Mammoto, T.; Charytan, D.; Strutz, F.; Kalluri, R. BMP-7 counteracts TGF-β1-induced epithelial-to-mesenchymal transition and reverses chronic renal injury. Nat. Med. 2003, 9, 964–968. [Google Scholar] [CrossRef] [PubMed]
- Sharma, G.D.; He, J.; Bazan, H.E. p38 and ERK1/2 coordinate cellular migration and proliferation in epithelial wound healing: Evidence of cross-talk activation between MAP kinase cascades. J. Biol. Chem. 2003, 278, 21989–21997. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Lu, L. Pathway-specific effect of caffeine on protection against UV irradiation-induced apoptosis in corneal epithelial cells. Investig. Ophthalmol. Vis. Sci. 2007, 48, 652–660. [Google Scholar] [CrossRef] [PubMed]
- Kubilus, J.K.; Beazley, K.E.; Talbot, C.J.; Linsenmayer, T.F. Nuclear ferritin mediated regulation of JNK signaling in corneal epithelial cells. Exp. Eye Res. 2016, 145, 337–340. [Google Scholar] [CrossRef] [PubMed]
- Lin, A. Activation of the JNK signaling pathway: Breaking the brake on apoptosis. Bioessays 2003, 25, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Peng, J.; Feng, D.; Chu, L.; Li, X.; Jin, Z.; Lin, Z.; Zeng, Q. Role of extracellular signal-regulated kinase, p38 kinase, and activator protein-1 in transforming growth factor-beta1-induced α smooth muscle actin expression in human fetal lung fibroblasts in vitro. Lung 2006, 184, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Sun, S.Q.; Hassid, A.; Ostrom, R.S. cAMP inhibits transforming growth factor-beta-stimulated collagen synthesis via inhibition of extracellular signal-regulated kinase 1/2 and smad signaling in cardiac fibroblasts. Mol. Pharmacol. 2006, 70, 1992–2003. [Google Scholar] [CrossRef] [PubMed]
- Xing, D.; Bonanno, J.A. Effect of cAMP on TGF-β1-induced corneal keratocyte-myofibroblast transformation. Investig. Ophthalmol. Vis. Sci. 2009, 50, 626–633. [Google Scholar] [CrossRef] [PubMed]
- Sebe, A.; Leivonen, S.K.; Fintha, A.; Masszi, A.; Rosivall, L.; Kahari, V.M.; Mucsi, I. Transforming growth factor-beta-induced α-smooth muscle cell actin expression in renal proximal tubular cells is regulated by p38beta mitogen-activated protein kinase, extracellular signal-regulated protein kinase1,2 and the smad signalling during epithelial-myofibroblast transdifferentiation. Nephrol. Dial. Transplant. 2008, 23, 1537–1545. [Google Scholar] [CrossRef] [PubMed]
- Stambe, C.; Atkins, R.C.; Tesch, G.H.; Masaki, T.; Schreiner, G.F.; Nikolic-Paterson, D.J. The role of p38α mitogen-activated protein kinase activation in renal fibrosis. J. Am. Soc. Nephrol. 2004, 15, 370–379. [Google Scholar] [CrossRef] [PubMed]
- Bodnar, R.J. Epidermal growth factor and epidermal growth factor receptor: The yin and yang in the treatment of cutaneous wounds and cancer. Adv. Wound Care 2013, 2, 24–29. [Google Scholar] [CrossRef] [PubMed]
- Wells, A. EGF receptor. Int. J. Biochem. Cell. Biol. 1999, 31, 637–643. [Google Scholar] [CrossRef]
- Zieske, J.D.; Takahashi, H.; Hutcheon, A.E.; Dalbone, A.C. Activation of epidermal growth factor receptor during corneal epithelial migration. Investig. Ophthalmol. Vis. Sci. 2000, 41, 1346–1355. [Google Scholar]
- Nakamura, Y.; Sotozono, C.; Kinoshita, S. The epidermal growth factor receptor (EGFR): Role in corneal wound healing and homeostasis. Exp. Eye Res. 2001, 72, 511–517. [Google Scholar] [CrossRef] [PubMed]
- Scholze, A.R.; Foo, L.C.; Mulinyawe, S.; Barres, B.A. BMP signaling in astrocytes downregulates EGFR to modulate survival and maturation. PLoS ONE 2014, 9, e110668. [Google Scholar] [CrossRef] [PubMed]
- Katz, M.; Amit, I.; Yarden, Y. Regulation of MAPKs by growth factors and receptor tyrosine kinases. Biochim. Biophys. Acta 2007, 1773, 1161–1176. [Google Scholar] [CrossRef] [PubMed]
- Sturla, L.M.; Amorino, G.; Alexander, M.S.; Mikkelsen, R.B.; Valerie, K.; Schmidt-Ullrichr, R.K. Requirement of Tyr-992 and Tyr-1173 in phosphorylation of the epidermal growth factor receptor by ionizing radiation and modulation by SHP2. J. Biol Chem. 2005, 280, 14597–14604. [Google Scholar] [CrossRef] [PubMed]
- Levkowitz, G.; Waterman, H.; Ettenberg, S.A.; Katz, M.; Tsygankov, A.Y.; Alroy, I.; Lavi, S.; Iwai, K.; Reiss, Y.; Ciechanover, A.; et al. Ubiquitin ligase activity and tyrosine phosphorylation underlie suppression of growth factor signaling by c-Cbl/Sli-1. Mol. Cell 1999, 4, 1029–1040. [Google Scholar] [CrossRef]
- Uchiyama-Tanaka, Y.; Matsubara, H.; Mori, Y.; Kosaki, A.; Kishimoto, N.; Amano, K.; Higashiyama, S.; Iwasaka, T. Involvement of HB-EGF and EGF receptor transactivation in TGF-β-mediated fibronectin expression in mesangial cells. Kidney Int. 2002, 62, 799–808. [Google Scholar] [CrossRef] [PubMed]
- Iwabu, A.; Smith, K.; Allen, F.D.; Lauffenburger, D.A.; Wells, A. Epidermal growth factor induces fibroblast contractility and motility via a protein kinase c delta-dependent pathway. J. Biol. Chem. 2004, 279, 14551–14560. [Google Scholar] [CrossRef] [PubMed]
- Tumur, Z.; Katebzadeh, S.; Guerra, C.; Bhushan, L.; Alkam, T.; Henson, B.S. RhoC mediates epidermal growth factor-stimulated migration and invasion in head and neck squamous cell carcinoma. Neoplasia 2015, 17, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Klemke, R.L.; Cai, S.; Giannini, A.L.; Gallagher, P.J.; de Lanerolle, P.; Cheresh, D.A. Regulation of cell motility by mitogen-activated protein kinase. J. Cell Biol. 1997, 137, 481–492. [Google Scholar] [CrossRef] [PubMed]
- Haase, I.; Evans, R.; Pofahl, R.; Watt, F.M. Regulation of keratinocyte shape, migration and wound epithelialization by IGF-1- and EGF-dependent signalling pathways. J. Cell Sci. 2003, 116, 3227–3238. [Google Scholar] [CrossRef] [PubMed]
- Jester, J.V.; Huang, J.; Barry-Lane, P.A.; Kao, W.W.; Petroll, W.M.; Cavanagh, H.D. Transforming growth factor(beta)-mediated corneal myofibroblast differentiation requires actin and fibronectin assembly. Investig. Ophthalmol. Vis. Sci. 1999, 40, 1959–1967. [Google Scholar]
- Hinz, B.; Celetta, G.; Tomasek, J.J.; Gabbiani, G.; Chaponnier, C. Alpha-smooth muscle actin expression upregulates fibroblast contractile activity. Mol. Biol. Cell 2001, 12, 2730–2741. [Google Scholar] [CrossRef] [PubMed]
- Tacke, F.; Gabele, E.; Bataille, F.; Schwabe, R.F.; Hellerbrand, C.; Klebl, F.; Straub, R.H.; Luedde, T.; Manns, M.P.; Trautwein, C.; et al. Bone morphogenetic protein 7 is elevated in patients with chronic liver disease and exerts fibrogenic effects on human hepatic stellate cells. Dig. Dis. Sci. 2007, 52, 3404–3415. [Google Scholar] [CrossRef] [PubMed]
- Pocsik, E.; Mihalik, R.; Ali-Osman, F.; Aggarwal, B.B. Cell density-dependent regulation of cell surface expression of two types of human tumor necrosis factor receptors and its effect on cellular response. J. Cell Biochem. 1994, 54, 453–464. [Google Scholar] [CrossRef] [PubMed]
- Hamburger, A.W.; Mehta, D.; Pinnamaneni, G.; Chen, L.C.; Reid, Y. Density-dependent regulation of epidermal growth factor receptor expression. Pathobiology 1991, 59, 329–334. [Google Scholar] [CrossRef] [PubMed]
- Petridou, S.; Maltseva, O.; Spanakis, S.; Masur, S.K. TGF-β receptor expression and smad2 localization are cell density dependent in fibroblasts. Investig. Ophthalmol. Vis. Sci. 2000, 41, 89–95. [Google Scholar]
- Nakamura, K.; Kurosaka, D.; Yoshino, M.; Oshima, T.; Kurosaka, H. Injured corneal epithelial cells promote myodifferentiation of corneal fibroblasts. Investig. Ophthalmol. Vis. Sci. 2002, 43, 2603–2608. [Google Scholar]
- Meng, X.M.; Chung, A.C.; Lan, H.Y. Role of the TGF-β/BMP-7/Smad pathways in renal diseases. Clin. Sci. 2013, 124, 243–254. [Google Scholar] [CrossRef] [PubMed]
- Yasmin, N.; Bauer, T.; Modak, M.; Wagner, K.; Schuster, C.; Koffel, R.; Seyerl, M.; Stockl, J.; Elbe-Burger, A.; Graf, D.; et al. Identification of bone morphogenetic protein 7 (BMP7) as an instructive factor for human epidermal langerhans cell differentiation. J. Exp. Med. 2013, 210, 2597–2610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robertson, D.M.; Li, L.; Fisher, S.; Pearce, V.P.; Shay, J.W.; Wright, W.E.; Cavanagh, H.D.; Jester, J.V. Characterization of growth and differentiation in a telomerase-immortalized human corneal epithelial cell line. Investig. Ophthalmol. Vis. Sci. 2005, 46, 470–478. [Google Scholar] [CrossRef] [PubMed]
- Dean, C.; Ito, M.; Makarenkova, H.P.; Faber, S.C.; Lang, R.A. BMP7 regulates branching morphogenesis of the lacrimal gland by promoting mesenchymal proliferation and condensation. Development 2004, 131, 4155–4165. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.; Zhou, X.; Zhou, X.; Pi, C.; Xu, R.; Wan, M.; Yang, J.; Zhou, Y.; Liu, C.; Sun, J.; et al. BMP7 and EREG contribute to the inductive potential of dental mesenchyme. Sci Rep. 2015, 5, 9903. [Google Scholar] [CrossRef] [PubMed]
- Ellman, M.B.; Kim, J.; An, H.S.; Chen, D.; Kc, R.; Li, X.; Xiao, G.; Yan, D.; Suh, J.; van Wjnen, A.J.; et al. Lactoferricin enhances BMP7-stimulated anabolic pathways in intervertebral disc cells. Gene 2013, 524, 282–291. [Google Scholar] [CrossRef] [PubMed]
- You, L.; Kruse, F.E. Differential effect of activin a and BMP-7 on myofibroblast differentiation and the role of the smad signaling pathway. Investig. Ophthalmol. Vis. Sci. 2002, 43, 72–81. [Google Scholar]
- Song, Q.H.; Singh, R.P.; Richardson, T.P.; Nugent, M.A.; Trinkaus-Randall, V. Transforming growth factor-beta1 expression in cultured corneal fibroblasts in response to injury. J. Cell Biochem. 2000, 77, 186–199. [Google Scholar] [CrossRef]
- Jester, J.V.; Barry-Lane, P.A.; Cavanagh, H.D.; Petroll, W.M. Induction of alpha-smooth muscle actin expression and myofibroblast transformation in cultured corneal keratocytes. Cornea 1996, 15, 505–516. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Gassmann, M.; Grenacher, B.; Rohde, B.; Vogel, J. Quantifying western blots: Pitfalls of densitometry. Electrophoresis 2009, 30, 1845–1855. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pathway Name | p-Value | KEGG |
|---|---|---|
| TGF-β signaling pathway | 5.780 × 10−5 | KEGG: 04350 |
| Cell cycle | 2.556 × 10−4 | KEGG: 04110 |
| Jak-STAT signaling pathway | 0.018 | KEGG: 04630 |
| MAPK signaling pathway | 0.040 | KEGG: 04010 |
| Osteoclast differentiation | 0.042 | KEGG: 04380 |
| Biological Processes | Differentially Expressed Genes/Total Genes | GO Accession | p-Value |
|---|---|---|---|
| Mitotic cell cycle | 111/216 | GO: 0000278 | 4.000 × 10−9 |
| Cell cycle | 149/321 | GO: 0007049 | 5.300 × 10−8 |
| Cytoskeleton organization | 82/153 | GO: 0007010 | 5.500 × 10−8 |
| Intermediate filament cytoskeleton organization | 10/10 | GO: 0045104 | 1.600 × 10−5 |
| Regulation of cell cycle | 82/171 | GO: 0051726 | 2.300 × 10−5 |
| Microtubule cytoskeleton organization | 42/73 | GO: 0000226 | 1.300 × 10−5 |
| Intermediate filament organization | 8/8 | GO: 0045109 | 1.500× 10−4 |
| Developmental process | 252/655 | GO: 0032502 | 3.100× 10−4 |
| Tissue development | 100/234 | GO: 0009888 | 8.300 × 10−4 |
| Negative regulation of protein phosphorylation | 26/46 | GO: 0001933 | 9.000 × 10−4 |
| Cell proliferation | 111/272 | GO: 0008283 | 0.003 |
| Epithelium development | 67/153 | GO: 0060429 | 0.003 |
| TGF-β receptor signaling pathway | 11/16 | GO: 0007179 | 0.004 |
| Inactivation of MAPK activity | 5/5 | GO: 0000188 | 0.004 |
| Regulation of microtubule cytoskeleton organization | 13/21 | GO: 0070507 | 0.007 |
| Positive regulation of Wnt signaling pathway | 12/19 | GO: 0030177 | 0.007 |
| Positive regulation of cytoskeleton organization | 26/52 | GO: 0051493 | 0.009 |
| MAPK cascade | 41/90 | GO: 0000165 | 0.009 |
| Signal transduction by protein phosphorylation | 42/93 | GO: 0023014 | 0.010 |
| Endothelial cell development | 4/4 | GO: 0001885 | 0.012 |
| Desmosome organization | 4/4 | GO: 0002934 | 0.012 |
| SMAD protein complex assembly | 4/4 | GO: 0007183 | 0.012 |
| Osteoblast proliferation | 4/4 | GO: 0033687 | 0.012 |
| Regulation of protein serine/threonine kinase activity | 31/66 | GO: 0071900 | 0.013 |
| Cell surface receptor signaling pathway | 102/258 | GO: 0007166 | 0.014 |
| Fibroblast proliferation | 10/16 | GO: 0048144 | 0.016 |
| SMAD protein signal transduction | 5/6 | GO: 0060395 | 0.018 |
| Regulation of actin polymerization or depolymerization | 8/12 | GO: 0008064 | 0.018 |
| Regulation of phosphorylation | 68/166 | GO: 0042325 | 0.019 |
| VEGF production | 7/10 | GO: 0010573 | 0.019 |
| Interleukin-2 production | 7/10 | GO: 0032623 | 0.019 |
| Cell adhesion | 79/197 | GO: 0007155 | 0.021 |
| Epithelial cell differentiation | 41/94 | GO: 0030855 | 0.021 |
| Response to BMP | 11/18 | GO: 0071772 | 0.024 |
| Regulation of ROS metabolic process | 14/26 | GO: 2000377 | 0.024 |
| Vesicle mediated transport | 51/122 | GO: 0016192 | 0.027 |
| Chemokine production | 9/15 | GO: 0032602 | 0.030 |
| Cell migration | 62/153 | GO: 0016477 | 0.031 |
| Cellular response to TGF-β stimulus | 14/27 | GO: 0071560 | 0.035 |
| Lipoxygenase pathway | 3/3 | GO: 0019372 | 0.037 |
| Intermediate filament bundle assembly | 3/3 | GO: 0045110 | 0.037 |
| Regulation of TGF-β receptor signaling pathway | 7/11 | GO: 0017015 | 0.038 |
| Cell differentiation | 151/407 | GO: 0030154 | 0.039 |
| Regulation of Interleukin-2 production | 6/9 | GO: 0032663 | 0.042 |
| Response to growth factor | 34/79 | GO: 0070848 | 0.042 |
| BMP signaling pathway | 10/18 | GO: 0030509 | 0.043 |
| ATP generation from ADP | 5/7 | GO: 0006757 | 0.045 |
| SMAD protein import to nucleus | 4/5 | GO: 0007184 | 0.045 |
| Regulation of cyclin dependent protein kinase activity | 12/23 | GO: 1904029 | 0.047 |
| Regeneration | 18/38 | GO: 0031099 | 0.049 |
| Fold Change (Down Regulated) | p-Value | Gene Symbol | Fold Change (Up Regulated) | p-Value | Gene Symbol |
|---|---|---|---|---|---|
| −64.32 | 0.00018 | HIST1H3G | 8.05 | 0.000406 | ALOX12B |
| −58.47 | 0.000071 | TOP2A | 8.36 | 0.000067 | BAMBI |
| −46.84 | 0.000005 | ANLN | 8.37 | 0.000013 | ID2 |
| −42.33 | 0.000046 | TFPI2 | 8.39 | 0.000018 | LCE3E |
| −41.85 | 0.000309 | HIST2H3A | 8.46 | 0.000245 | BDNF |
| −37.77 | 0.000009 | NCAPG | 8.49 | 0.000242 | DLX3 |
| −36.57 | 0.000029 | CCNA2 | 8.51 | 0.000018 | FAM46B |
| −36.48 | 0.000107 | RRM2 | 8.54 | 0.00008 | CGB2 |
| −34.74 | 0.000308 | HIST2H3A; HIST2H3C | 8.78 | 0.000024 | HK2 |
| −29.49 | 0.00016 | TTK | 8.83 | 0.000403 | FPR3 |
| −27.57 | 0.000001 | FAM111B | 8.84 | 0.00021 | BMP2 |
| −25.32 | 7.63 × 10−7 | NUSAP1 | 9 | 0.000011 | CHRNA3 |
| −24.3 | 0.000036 | PLK1 | 9.3 | 0.000094 | ULK3 |
| −20.9 | 0.000371 | DLGAP5 | 9.58 | 0.000057 | DUSP1 |
| −19.5 | 0.000003 | MKI67 | 9.6 | 0.000018 | GCOM1 |
| −16.79 | 0.000208 | IL1B | 9.74 | 0.000781 | GABRA3 |
| −16.57 | 0.000092 | CCNB2 | 9.74 | 0.000021 | SDR9C7 |
| −16.55 | 0.000032 | ASPM | 9.74 | 0.000281 | RNF39 |
| −16.27 | 0.000055 | KIF18A | 9.81 | 0.000849 | CYP4F22 |
| −16.25 | 0.000072 | DTL | 9.92 | 0.00001 | DUSP14 |
| −15.81 | 0.000586 | HIST1H1B | 10.18 | 0.000032 | MXD1 |
| −15.57 | 0.00003 | GPR50 | 10.34 | 0.00026 | ST3GAL4 |
| −15.31 | 0.000043 | CENPF | 10.44 | 0.000002 | SPOPL |
| −15.23 | 0.000017 | SHCBP1 | 10.58 | 0.000063 | SLC7A8 |
| −14.83 | 0.000305 | PBK | 10.67 | 0.00079 | LCE3C |
| −14.71 | 0.000777 | CKAP2L | 10.72 | 0.000034 | PSCA |
| −14.61 | 0.00001 | KIF20A | 10.89 | 0.000199 | DLX2 |
| −14.2 | 0.000094 | LMNB1 | 11.02 | 0.000084 | LCE3D |
| −14.13 | 0.000024 | MELK | 11.17 | 0.000127 | PLBD1 |
| −13.16 | 0.000482 | PRC1 | 12.56 | 0.000338 | KPRP |
| −12.65 | 0.000038 | DEPDC1 | 12.88 | 0.000074 | ATP1B1 |
| −12.56 | 0.000045 | TACC3 | 13.24 | 0.000107 | CGB1 |
| −12.53 | 0.000212 | HELLS | 13.68 | 0.000006 | EPN3 |
| −12.45 | 0.000029 | NCAPG2 | 14.25 | 0.000003 | C10orf99 |
| −12.25 | 0.000355 | BUB1 | 14.57 | 0.000011 | C5orf46 |
| −12.06 | 0.000473 | CASC5 | 14.65 | 0.000002 | BPGM |
| −11.89 | 0.000104 | NDC1 | 14.84 | 0.000014 | GNA14 |
| −11.81 | 0.000635 | FAM83D | 14.93 | 0.000294 | C1QTNF3-AMACR |
| −11.8 | 0.000488 | CCNE2 | 14.96 | 0.000013 | CGB5; CGB8 |
| −11.43 | 0.000527 | KIF23 | 14.97 | 0.000096 | SERPINB12 |
| −11.17 | 0.000167 | CDC20 | 15.66 | 0.000279 | MARCH3 |
| −11.1 | 0.000901 | H2AFX | 16.07 | 3.57 × 10−7 | VSIG8 |
| −10.97 | 0.000061 | KIF2C | 16.67 | 0.000159 | USP2 |
| −10.77 | 0.000126 | STIL | 17.7 | 0.000147 | SLAMF9 |
| −10.75 | 0.000582 | NCAPH | 18.03 | 0.000376 | SPAG17 |
| −10.57 | 0.000444 | FOXM1 | 18.8 | 0.000024 | CGB8 |
| −10.23 | 0.00036 | CCNB1 | 18.93 | 0.000319 | CLDN17 |
| −10.09 | 0.000184 | TPX2 | 18.98 | 0.000083 | NPR3 |
| −10.06 | 0.000045 | MCM5 | 19.8 | 0.000229 | CGB; CGB5 |
| −9.91 | 0.000196 | XRCC2 | 19.94 | 0.000024 | LOX |
| −9.78 | 0.000011 | KIF14 | 20 | 0.000026 | HPGD |
| −9.44 | 0.000053 | ARHGAP11A | 26.9 | 0.000814 | SPINK7 |
| −9.4 | 0.000046 | DIAPH3 | 28.3 | 0.000024 | CRYAB |
| −9.26 | 0.000008 | MCM3 | 29.87 | 0.000029 | DIO2 |
| −8.97 | 0.000141 | CDC6 | 37.64 | 0.000037 | ID1 |
| −8.93 | 0.000088 | ARHGAP11B | 43.55 | 3.21 × 10−7 | DSG4 |
| −8.73 | 0.000689 | SGOL1 | 56.48 | 0.000029 | SMAD6 |
| −8.61 | 0.00009 | FANCA | 64.3 | 0.000012 | AMACR |
| −8.5 | 0.000047 | HJURP | 96.26 | 0.000007 | TGM5 |
| −8.39 | 0.000032 | ADORA2B | 98.81 | 0.000061 | SPINK6 |
| −8.01 | 0.000659 | TMEM97 | 135.1 | 9.72 × 10−7 | FABP4 |
| Target | Primer | Primer Sequence (5’-3’) | GenBank Accession |
|---|---|---|---|
| ATP1B1 | fwd | TGAATTTAAGCCCACATATCAGGACCG | NM_001677 |
| rev | CATGTCATCCCTCTGGGCTGAATCT | ||
| CRYAB | fwd | GGGAGATGTGATTGAGGTGCATGG | NM_001289807 |
| rev | AGGACCCCATCAGATGACAGGGAT | ||
| DSG4 | fwd | GCAGCCTGTCGAGAAGGAGAGGACA | NM_001134453 |
| rev | GTGATGTTAATTTCCCCAGTGCGAGG | ||
| EPN3 | fwd | CTGCGCCTGAGCCGGCAG | NM_017957 |
| rev | TCAGGCTCTCTGTCCCGCTGATG | ||
| ID1 | fwd | AACGGCGAGATCAGCGCCCT | NM_002165 |
| rev | TGTTCTCCCTCAGATCCGGCGA | ||
| BMP7 | fwd | TCACAGCCACCAGCAACCACTG | NM_001719 |
| rev | ACCATGAAGGGCTGCTTGTTCT | ||
| ID2 | fwd | GCAGATCGCCCTGGACTCGCA | NM_002166 |
| rev | CACCGCTTATTCAGCCACACAGTGC | ||
| LOX | fwd | TTACCCAGCCGACCAAGATATTCCTG | NM_002317 |
| rev | TTGTGGCCTTCAGCCACTCTCCTC | ||
| TGM5 | fwd | AGGCTGGCAGGTGCTGGACG | NM_004245 |
| rev | CCTGGACGAGCCAGGACATGC | ||
| DUSP14 | fwd | CAGCGCACACTGGACTCTTGAGG | NM_007026 |
| rev | GTCCTTGGTAGCGTGCTGTGACCT | ||
| GAPDH | fwd | GGATATTGTTGCCATCAATGACCC | NM_002046 |
| rev | TCTCGCTCCTGGAAGATGGTGA |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kowtharapu, B.S.; Prakasam, R.K.; Murín, R.; Koczan, D.; Stahnke, T.; Wree, A.; Jünemann, A.G.M.; Stachs, O. Role of Bone Morphogenetic Protein 7 (BMP7) in the Modulation of Corneal Stromal and Epithelial Cell Functions. Int. J. Mol. Sci. 2018, 19, 1415. https://doi.org/10.3390/ijms19051415
Kowtharapu BS, Prakasam RK, Murín R, Koczan D, Stahnke T, Wree A, Jünemann AGM, Stachs O. Role of Bone Morphogenetic Protein 7 (BMP7) in the Modulation of Corneal Stromal and Epithelial Cell Functions. International Journal of Molecular Sciences. 2018; 19(5):1415. https://doi.org/10.3390/ijms19051415
Chicago/Turabian StyleKowtharapu, Bhavani S., Ruby Kala Prakasam, Radovan Murín, Dirk Koczan, Thomas Stahnke, Andreas Wree, Anselm G. M. Jünemann, and Oliver Stachs. 2018. "Role of Bone Morphogenetic Protein 7 (BMP7) in the Modulation of Corneal Stromal and Epithelial Cell Functions" International Journal of Molecular Sciences 19, no. 5: 1415. https://doi.org/10.3390/ijms19051415