Reproduction of Characteristics of Extracellular Matrices in Specific Longitudinal Depth Zone Cartilage within Spherical Organoids in Response to Changes in Osmotic Pressure

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
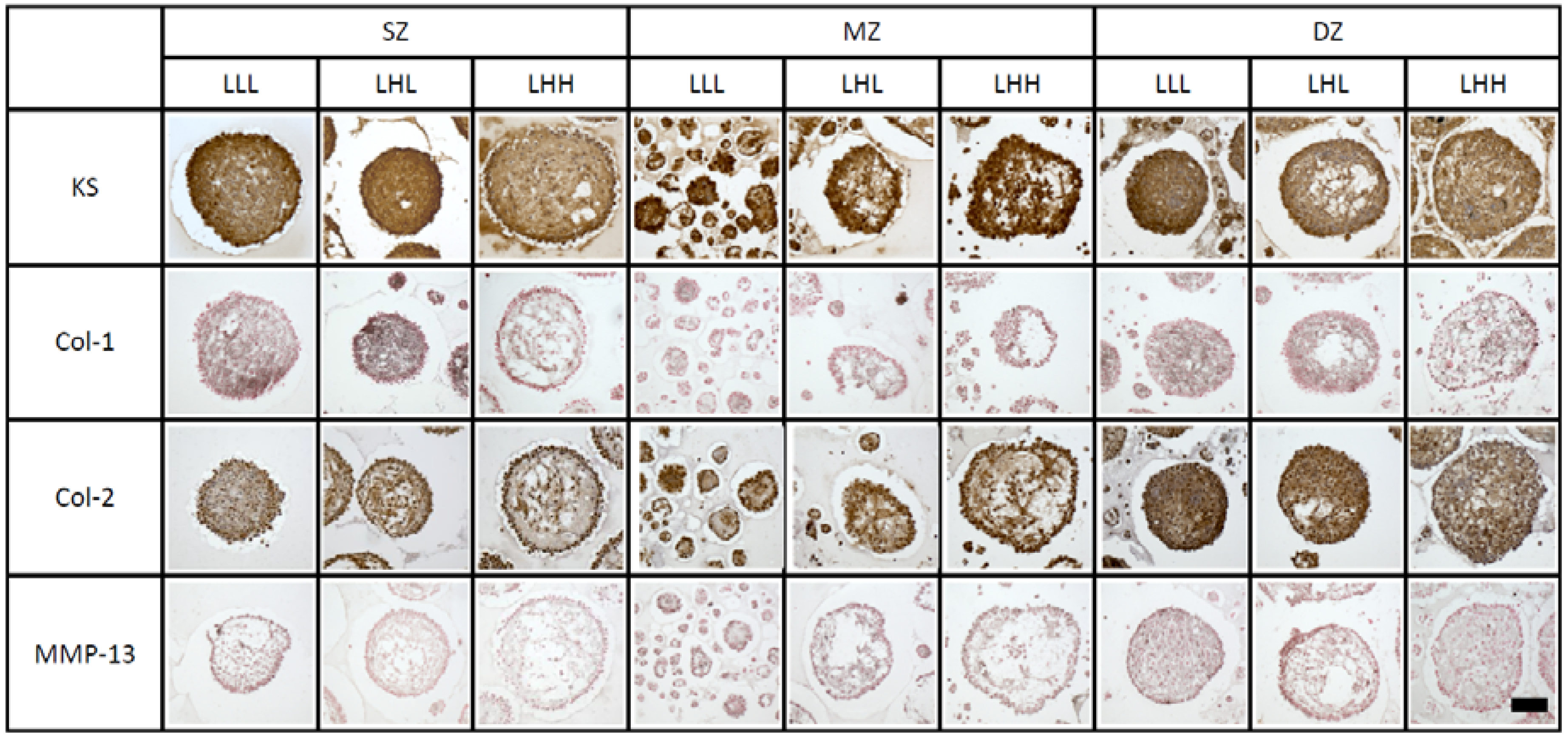
2. Results
2.1. Formation of Spheroids
2.2. Gene Expression Profiles
2.3. Histological Characteristics of Spheroids
3. Discussion
3.1. Morphological Characteristics of Spheroids Formed with Specific Longitudinal Depth Zone-Derived Chondrocytes in Response to ΔOP
3.2. Metabolic Functions of Chondrocytes Isolated from a Specific Zone in Response to ΔOP
4. Materials and Methods
4.1. Cell Isolation from Specific Depth Zones in Articular Cartilage
4.2. Changes in Osmotic Pressure
4.3. Measurement of Gene Expression by Quantitative Polymerase Chain Reaction (qPCR)
4.4. Histological Characteristics
4.5. Data Analysis of Gene Expression Profiles Using a qPCR Assay
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Urban, J.P.G. Present perspectives on cartilage and chondrocyte mechanobiology. Biorheology 2000, 37, 185–190. [Google Scholar] [PubMed]
- Maroudas, A. Physical chemistry of articular cartilage and the intervertebral disc. In The Joints and Synovial Fluid; Sokoloff, L., Ed.; Academic Press: New York, NY, USA, 1981; pp. 240–291. [Google Scholar]
- Schneiderman, R.; Keret, D.; Maroudas, A. Effects of mechanical and osmotic pressure on the rate of glycosaminoglycan synthesis in the human adult femoral head cartilage: An in vitro study. J. Orthop. Res. 1986, 4, 393–408. [Google Scholar] [CrossRef] [PubMed]
- Mow, V.C.; Wang, C.C.; Hung, C.T. The extracellular matrix, interstitial fluid and ions as a mechanical signal transducer in articular cartilage. Osteoarthr. Cartil. 1999, 7, 41–58. [Google Scholar] [CrossRef] [PubMed]
- Lai, W.M.; Gu, W.Y.; Mow, V.C. On the conditional equivalence of chemical loading and mechanical loading on articular cartilage. J. Biomech. 1998, 31, 1181–1185. [Google Scholar] [CrossRef]
- Sanchez, J.C.; Wilkins, R.J. Changes in intracellular calcium concentration in response to hypertonicity in bovine articular chondrocytes. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2004, 137, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.D.; Oloyede, A.; Singh, S.; Gu, Y. Investigation of the effects of extracellular osmotic pressure on morphology and mechanical properties of individual chondrocyte. Cell Biochem. Biophys. 2016, 74, 229–240. [Google Scholar] [CrossRef] [PubMed]
- Negoro, K.; Kobayashi, S.; Takeno, K.; Uchida, K.; Baba, H. Effect of osmolarity on glycosaminoglycan production and cell metabolism of articular chondrocyte under three-dimensional culture system. Clin. Exp. Rheumatol. 2008, 26, 534–541. [Google Scholar] [PubMed]
- Bush, P.G.; Hall, A.C. The osmotic sensitivity of isolated and in situ bovine articular chondrocytes. J. Orthop. Res. 2001, 19, 768–778. [Google Scholar] [CrossRef]
- Mizuno, S.; Takada, E.; Fukai, N. Spherical organoids reproduce characteristics of longitudinal depth zones in bovine articular cartilage. Cells Tissue Organs 2016, 202, 382–392. [Google Scholar] [CrossRef] [PubMed]
- Kato, Y.; Iwamoto, M.; Koike, T.; Suzuki, F.; Takano, Y. Terminal differentiation and calcification in rabbit chondrocyte cultures grown in centrifuge tubes: Regulation by transforming growth factor b and serum factors. Proc. Natl. Acad. Sci. USA 1988, 85, 9552–9556. [Google Scholar] [CrossRef] [PubMed]
- Oster, G.F.; Murray, J.D.; Manini, P.K. A model for chondrogenic condensations in the developing limb: The role of extracellular matrix and cell tractions. J. Embryol. Exp. Morphol. 1985, 89, 93–112. [Google Scholar] [PubMed]
- Roberts, S.; Menage, J.; Sandell, L.J.; Evans, E.H.; Richardson, J.B. Immunohistochemical study of collagen type I and II and procollagen IIA in human cartilage repair tissue following autologous chondrocyte implantation. Knee 2009, 16, 398–404. [Google Scholar] [CrossRef] [PubMed]
- Schattis, O.R.; Markova, M.; Torzilli, P.A.; Gallo, L.M. Mechanical loading of cartilage explants with compression and sliding motion modulates gene expression of lubricin and catabolic enzyme. Cartilage 2015, 6, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Hladky, S.B.; Rink, T.J. Osmotic behavior of human red blood cells: An interpretation in terms if negative intracellular fluid pressure. J. Physiol. 1978, 274, 437–446. [Google Scholar] [CrossRef] [PubMed]
- Weiss, T.F. Equilibration times for regular geometric volumes. In Cellular Biophysics; MIT Press: Cambridge, MA, USA, 1996; pp. 117–118. [Google Scholar]
- Mizuno, S.; Ogawa, R. Using changes in hydrostatic and osmotic pressure to manipulate metabolic function in chondrocytes. Am. J. Physiol. Cell Physiol. 2011, 300, C1234–C1245. [Google Scholar] [CrossRef] [PubMed]
- Jablonski, C.L.; Ferguson, S.; Pozzi, A.; Clark, A.L. Integrin α1β1 participates in chondrocyte transduction of osmotic stress. Biochem. Biophys. Res. Commun. 2014, 445, 184–190. [Google Scholar] [CrossRef] [PubMed]
- Szafranski, J.D.; Grodzinsky, A.J.; Berger, E.; Gaschen, V.; Hung, H.H.; Hunziker, E.B. Chondrocyte mechanotransduction: Effects of compression deformation of intracellular organelles and relevance to cellular biosynthesis. Osteoarthr. Cartil. 2004, 12, 937–946. [Google Scholar] [CrossRef] [PubMed]
- Hui, A.Y.; McCarty, W.J.; Masuda, K.; Firestein, G.S.; Sah, R.L. A system biology approach to synovium joint lubrication in health, injury, and disease. Wiley Interdiscip. Rev. Syst. Biol. Med. 2013, 4, 15–37. [Google Scholar] [CrossRef] [PubMed]
- Hing, W.A.; Sherwin, A.F.; Poole, C.A. The influence of the pericellular microenvironment on the chondrocyte response to osmotic challenge. Osteoarthr. Cartil. 2002, 10, 297–307. [Google Scholar] [CrossRef] [PubMed]
- Korhonen, R.K.; Han, S.K.; Herzog, W. Osmotic loading of articular cartilage modulates cell deformations along primary collagen fibril directions. J. Biomech. 2010, 43, 783–787. [Google Scholar] [CrossRef] [PubMed]
- Khoshgoftar, M.; Wilson, W.; Ito, K.; van Donkelaar, C.C. Influence of tissue- and cell- scale extracellular matrix distribution on the mechanical properties of tissue-engineered cartilage. Biomech. Model. Mechanobiol. 2013, 12, 901–913. [Google Scholar] [CrossRef] [PubMed]
- Nims, R.J.; Cigan, A.D.; Durney, K.M.; Jones, B.K.; O’Neil, J.D.; Law, W.A.; Vunjak-Novakovic, G.; Hung, C.T.; Ateshian, G.A. Constrained Cage culture improves engineered cartilage functional properties by enhancing collagen network stability. Tissue Eng. Part A 2017, 15–16, 847–858. [Google Scholar] [CrossRef] [PubMed]
- Thronemo, M.; Tallheden, T.; Sjogren-Hasson, E.; Larsson, A.; Lovstedt, K.; Nannmark, U.; Brittberg, M.; Lindahl, A. Clonal populations of chondocytes with progenitor properties identified within human articular cartilage. Cell Tissue Organ 2005, 180, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Lorenzo, P.; Bayliss, M.T.; Heinegard, D. A novel cartilage protein (CILP) present in the mid-zone of human articular cartilage increases with age. J. Biol. Chem. 1998, 273, 23463–23468. [Google Scholar] [CrossRef] [PubMed]
- Poole, C.A.; Flint, M.H.; Beaumont, B.W. Chondrons in cartilage: Ultrastructural analysis of the pericellular microenvironment in adult human articular cartilage. J. Orthop. Res. 1987, 5, 509–522. [Google Scholar] [CrossRef] [PubMed]
- Dowthwaite, G.P.; Bishop, J.C.; Redman, S.N. The surface of articular cartilage contains a progenitor cell population. J. Cell Sci. 2004, 117, 889–897. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, S. A novel method for assessing effects of hydrostatic fluid pressure on intracellular calcium: A study with bovine articular chondrocytes. Am. J. Physiol. Cell Physiol. 2005, 288, C329–C337. [Google Scholar] [CrossRef] [PubMed]




© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Takada, E.; Mizuno, S. Reproduction of Characteristics of Extracellular Matrices in Specific Longitudinal Depth Zone Cartilage within Spherical Organoids in Response to Changes in Osmotic Pressure. Int. J. Mol. Sci. 2018, 19, 1507. https://doi.org/10.3390/ijms19051507
Takada E, Mizuno S. Reproduction of Characteristics of Extracellular Matrices in Specific Longitudinal Depth Zone Cartilage within Spherical Organoids in Response to Changes in Osmotic Pressure. International Journal of Molecular Sciences. 2018; 19(5):1507. https://doi.org/10.3390/ijms19051507
Chicago/Turabian StyleTakada, Eiichiro, and Shuichi Mizuno. 2018. "Reproduction of Characteristics of Extracellular Matrices in Specific Longitudinal Depth Zone Cartilage within Spherical Organoids in Response to Changes in Osmotic Pressure" International Journal of Molecular Sciences 19, no. 5: 1507. https://doi.org/10.3390/ijms19051507
APA StyleTakada, E., & Mizuno, S. (2018). Reproduction of Characteristics of Extracellular Matrices in Specific Longitudinal Depth Zone Cartilage within Spherical Organoids in Response to Changes in Osmotic Pressure. International Journal of Molecular Sciences, 19(5), 1507. https://doi.org/10.3390/ijms19051507