Enhancement of Recombinant Protein Production in Transgenic Nicotiana benthamiana Plant Cell Suspension Cultures with Co-Cultivation of Agrobacterium Containing Silencing Suppressors


Abstract
:
1. Introduction
2. Results
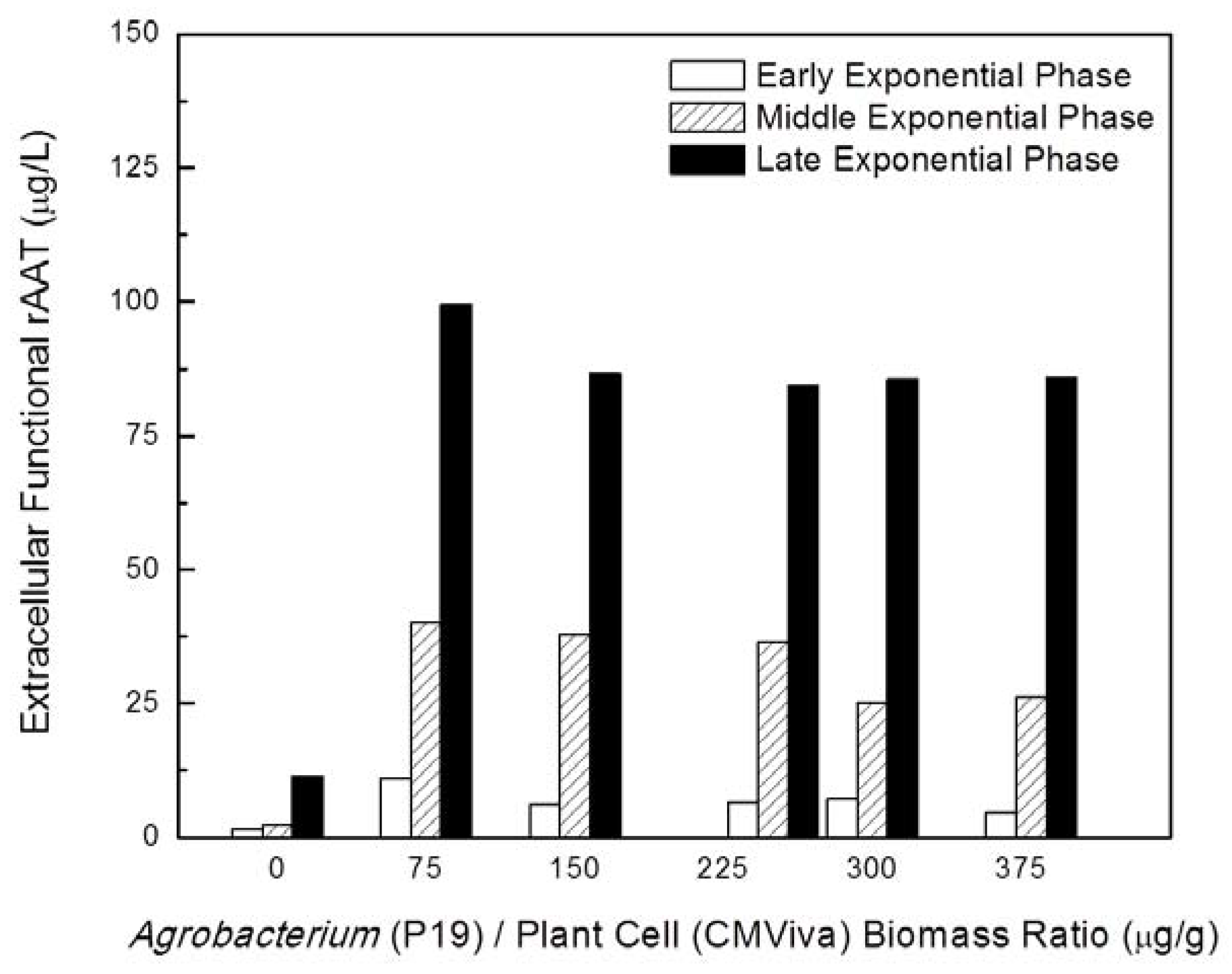
2.1. Development of Transient Expression Co-Cultivation Process
2.2. Effect of Viral Gene Silencing Suppressors on rAAT Production in Transgenic 35S, XVE and CMViva Cell Cultures
2.3. Impact of Co-Expression of Viral Gene Silencing Suppressors on rAAT Production Using Design of Experiment
3. Discussion
4. Materials and Methods
4.1. Expression Systems Construction and Plant Cell Transformation
4.2. Transgenic Plant Cell Cultures and Media
4.3. Biomass Concentration Determination
4.4. Agrobacterium Cultures and Viral Gene Silencing Suppressors
4.5. Co-Culture of Agrobacterium with Transgenic Plant Cell Suspensions
4.6. Induction Treatment
4.7. Protein Quantitative and Qualitative Analysis
4.8. Analysis of the Impact of Viral Gene Silencing on PTGS
4.9. Fractional Factorial Design (FFD)
4.10. Response Surface Methodology (RSM)
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Burgyan, J. Role of silencing suppressor proteins. Methods Mol. Biol. 2008, 451, 69–79. [Google Scholar] [PubMed]
- Hamilton, A.J.; Baulcombe, D.C. A species of small antisense RNA in posttranscriptional gene silencing in plants. Science 1999, 286, 950–952. [Google Scholar] [CrossRef] [PubMed]
- Voinnet, O. Induction and suppression of RNA silencing: Insights from viral infections. Nat. Rev. 2005, 6, 206–220. [Google Scholar] [CrossRef] [PubMed]
- Meins, F., Jr. RNA degradation and models for post-transcriptional gene-silencing. Plant Mol. Biol. 2000, 43, 261–273. [Google Scholar] [CrossRef] [PubMed]
- Bernstein, E.; Caudy, A.A.; Hammond, S.M.; Hannon, G.J. Role for a bidentate ribonuclease in the initiation step of RNA interference. Nature 2001, 409, 363–366. [Google Scholar] [CrossRef] [PubMed]
- Levy, A.; Dafny-Yelin, M.; Tzfira, T. Attacking the defenders: Plant viruses fight back. Trends Microbiol. 2008, 16, 194–197. [Google Scholar] [CrossRef] [PubMed]
- Baulcombe, D. RNA silencing in plants. Nature 2004, 431, 356–363. [Google Scholar] [CrossRef] [PubMed]
- Mlotshwa, S.; Pruss, G.J.; Vance, V. Small RNAs in viral infection and host defense. Trends Plant Sci. 2008, 13, 375–382. [Google Scholar] [CrossRef] [PubMed]
- Voinnet, O. Use, tolerance and avoidance of amplified RNA silencing by plants. Trends Plant Sci. 2008, 13, 317–328. [Google Scholar] [CrossRef] [PubMed]
- Voinnet, O.; Pinto, Y.M.; Baulcombe, D.C. Suppression of gene silencing: A general strategy used by diverse DNA and RNA viruses of plants. Proc. Natl. Acad. Sci. USA 1999, 96, 14147–14152. [Google Scholar] [CrossRef] [PubMed]
- Anandalakshmi, R.; Pruss, G.J.; Ge, X.; Marathe, R.; Mallory, A.C.; Smith, T.H.; Vance, V.B. A viral suppressor of gene silencing in plants. Proc. Natl. Acad. Sci. USA 1998, 95, 13079–13084. [Google Scholar] [CrossRef] [PubMed]
- Roth, B.M.; Pruss, G.J.; Vance, V.B. Plant viral suppressors of RNA silencing. Virus Res. 2004, 102, 97–108. [Google Scholar] [CrossRef] [PubMed]
- Chapman, E.J.; Prokhnevsky, A.I.; Gopinath, K.; Dolja, V.V.; Carrington, J.C. Viral RNA silencing suppressors inhibit the microRNA pathway at an intermediate step. Genes Dev. 2004, 18, 1179–1186. [Google Scholar] [CrossRef] [PubMed]
- Lakatos, L.; Szittya, G.; Silhavy, D.; Burgyan, J. Molecular mechanism of RNA silencing suppression mediated by p19 protein of tombusviruses. EMBO J. 2004, 23, 876–884. [Google Scholar] [CrossRef] [PubMed]
- Fischer, R.; Vaquero-Martin, C.; Sack, M.; Drossard, J.; Emans, N.; Commandeur, U. Towards molecular farming in the future: Transient protein expression in plants. Biotechnol. Appl. Biochem. 1999, 30, 113–116. [Google Scholar] [PubMed]
- Johansen, L.K.; Carrington, J.C. Silencing on the spot. Induction and suppression of RNA silencing in the Agrobacterium-mediated transient expression system. Plant Physiol. 2001, 126, 930–938. [Google Scholar] [CrossRef] [PubMed]
- Marillonnet, S.; Thoeringer, C.; Kandzia, R.; Klimyuk, V.; Gleba, Y. Systemic Agrobacterium tumefaciens-mediated transfection of viral replicons for efficient transient expression in plants. Nat. Biotechnol. 2005, 23, 718–723. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Lai, H.; Hurtado, J.; Stahnke, J.; Leuzinger, K.; Dent, M. Agroinfiltration as an effective and scalable strategy of gene delivery for production of pharmaceutical proteins. Adv. Tech. Biol. Med. 2013, 1, 103. [Google Scholar] [CrossRef] [PubMed]
- McCullen, C.A.; Binns, A.N. Agrobacterium tumefaciens and plant cell interactions and activities required for interkingdom macromolecular transfer. Annu. Rev. Cell Dev. Biol. 2006, 22, 101–127. [Google Scholar] [CrossRef] [PubMed]
- Kapila, J.; DeRycke, R.; VanMontagu, M.; Angenon, G. An Agrobacterium-mediated transient gene expression system for intact leaves. Plant Sci. 1997, 122, 101–108. [Google Scholar] [CrossRef]
- Vaquero, C.; Sack, M.; Chandler, J.; Drossard, J.; Schuster, F.; Monecke, M.; Schillberg, S.; Fischer, R. Transient expression of a tumor-specific single-chain fragment and a chimeric antibody in tobacco leaves. Proc. Natl. Acad. Sci. USA 1999, 96, 11128–11133. [Google Scholar] [CrossRef] [PubMed]
- Cazzonelli, C.I.; Velten, J. An in vivo, luciferase-based, Agrobacterium-infiltration assay system: Implications for post-transcriptional gene silencing. Planta 2006, 224, 582–597. [Google Scholar] [CrossRef] [PubMed]
- Johansen, W.; Wilson, R.C. Viral suppressor proteins show varying abilities and effectiveness to suppress transgene-induced post-transcriptional gene silencing of endogenous Chalcone synthase in transgenic Arabidopsis. Plant Cell Rep. 2008, 27, 911–921. [Google Scholar] [CrossRef] [PubMed]
- Ma, P.; Liu, J.; He, H.; Yang, M.; Li, M.; Zhu, X.; Wang, X. A Viral Suppressor P1/HC-Pro Increases the GFP Gene Expression in Agrobacterium-mediated Transient Assay. Appl. Biochem. Biotechnol. 2009, 158, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Sudarshana, M.R.; Plesha, M.A.; Uratsu, S.L.; Falk, B.W.; Dandekar, A.M.; Huang, T.-K.; McDonald, K.A. A chemically inducible cucumber mosaic virus amplicon system for expression of heterologous proteins in plant tissues. Plant Biotechnol. J. 2006, 4, 551–559. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.-K.; Plesha, M.A.; Falk, B.W.; Dandekar, A.M.; McDonald, K.A. Bioreactor strategies for improving production yield and functionality of a recombinant human protein in transgenic tobacco cell cultures. Biotechnol. Bioeng. 2009, 102, 508–520. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.-K.; Plesha, M.A.; McDonald, K.A. Semicontinuous bioreactor production of a recombinant human therapeutic protein using a chemically inducible viral amplicon expression system in transgenic plant cell suspension cultures. Biotechnol. Bioeng. 2010, 106, 408–421. [Google Scholar] [CrossRef] [PubMed]
- Zuo, J.R.; Niu, Q.W.; Chua, N.H. An estrogen receptor-based transactivator XVE mediates highly inducible gene expression in transgenic plants. Plant J. 2000, 24, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Brigneti, G.; Voinnet, O.; Li, W.X.; Ji, L.H.; Ding, S.W.; Baulcombe, D.C. Viral pathogenicity determinants are suppressors of transgene silencing in Nicotiana benthamiana. EMBO J. 1998, 17, 6739–6746. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Yuan, Y.R.; Pei, Y.; Lin, S.S.; Tuschl, T.; Patel, D.J.; Chua, N.H. Cucumber mosaic virus-encoded 2b suppressor inhibits Arabidopsis Argonaute1 cleavage activity to counter plant defense. Genes Dev. 2006, 20, 3255–3268. [Google Scholar] [CrossRef] [PubMed]
- Qu, F.; Ren, T.; Morris, T.J. The coat protein of turnip crinkle virus suppresses posttranscriptional gene silencing at an early initiation step. J. Virol. 2003, 77, 511–522. [Google Scholar] [CrossRef] [PubMed]
- Bonneau, C.; Brugidou, C.; Chen, L.; Beachy, R.N.; Fauquet, C. Expression of the rice yellow mottle virus P1 protein in vitro and in vivo and its involvement in virus spread. Virology 1998, 244, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Dell’Orco, M.; Saldarelli, P.; Turturo, C.; Minafra, A.; Martelli, G.P. Identification of an RNA-silencing suppressor in the genome of Grapevine virus A. J. Gen. Virol. 2006, 87, 2387–2395. [Google Scholar] [CrossRef] [PubMed]
- Reed, J.C.; Kasschau, K.D.; Prokhnevsky, A.I.; Gopinath, K.; Pogue, G.P.; Carrington, J.C.; Dolja, V.V. Suppressor of RNA silencing encoded by Beet yellows virus. Virology 2003, 306, 203–209. [Google Scholar] [CrossRef]
- Chiba, M.; Reed, J.C.; Prokhnevsky, A.I.; Chapman, E.J.; Mawassi, M.; Koonin, E.V.; Carrington, J.C.; Dolja, V.V. Diverse suppressors of RNA silencing enhance agroinfection by a viral replicon. Virology 2006, 346, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Tamai, A.; Meshi, T. Cell-to-cell movement of potato virus X: The role of p12 and p8 encoded by the second and third open reading frames of the triple gene block. Mol. Plant-Microbe Interact. 2001, 14, 1158–1167. [Google Scholar] [CrossRef] [PubMed]
- Chellappan, P.; Vanitharani, R.; Ogbe, F.; Fauquet, C.M. Effect of temperature on geminivirus-induced RNA silencing in plants. Plant Physiol. 2005, 138, 1828–1841. [Google Scholar] [CrossRef] [PubMed]
- Kameda, T.; Ikegami, K.; Liu, Y.; Terada, K.; Sugiyama, T. A hypothermic-temperature-sensitive gene silencing by the mammalian RNAi. Biochem. Biophys. Res. Commun. 2004, 315, 599–602. [Google Scholar] [CrossRef] [PubMed]
- Szittya, G.; Silhavy, D.; Molnar, A.; Havelda, Z.; Lovas, A.; Lakatos, L.; Banfalvi, Z.; Burgyan, J. Low temperature inhibits RNA silencing-mediated defence by the control of siRNA generation. EMBO J. 2003, 22, 633–640. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, K.M.; Larsen, J.S.; Curtis, W.R. Scale-up of Agrobacterium-mediated transient protein expression in bioreactor-grown Nicotiana glutinosa plant cell suspension culture. Biotechnol. Prog. 2008, 24, 372–376. [Google Scholar] [CrossRef] [PubMed]
- Voinnet, O.; Rivas, S.; Mestre, P.; Baulcombe, D. An enhanced transient expression system in plants based on suppression of gene silencing by the p19 protein of tomato bushy stunt virus. Plant J. 2003, 33, 949–956. [Google Scholar] [CrossRef] [PubMed]
- Wroblewski, T.; Tomczak, A.; Michelmore, R. Optimization of Agrobacterium-mediated transient assays of gene expression in lettuce, tomato and Arabidopsis. Plant Biotechnol. J. 2005, 3, 259–273. [Google Scholar] [CrossRef] [PubMed]
- Plesha, M.A.; Huang, T.-K.; Dandekar, A.M.; Falk, B.W.; McDonald, K.A. High-level transient production of a heterologous protein in plants by optimizing induction of a chemically inducible viral amplicon expression system. Biotechnol. Prog. 2007, 23, 1277–1285. [Google Scholar] [CrossRef] [PubMed]
- Plesha, M.A.; Huang, T.-K.; Dandekar, A.M.; Falk, B.W.; McDonald, K.A. Optimization of the Bioprocessing Conditions for Scale-Up of Transient Production of a Heterologous Protein in Plants Using a Chemically Inducible Viral Amplicon Expression System. Biotechnol. Prog. 2009, 25, 722–734. [Google Scholar] [CrossRef] [PubMed]
- Arzola, L.; Chen, J.; Rattanaporn, K.; Maclean, J.M.; McDonald, K.A. Transient co-expression of post-transcriptional gene silencing suppressors for increased in planta expression of a recombinant anthrax receptor fusion protein. Int. J. Mol. Sci. 2011, 12, 4975–4990. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.-K.; McDonald, K.A. Bioreactor systems for in vitro production of foreign proteins using plant cell cultures. Biotechnol. Adv. 2012, 30, 398–409. [Google Scholar] [CrossRef] [PubMed]
- Tekoah, Y.; Shulman, A.; Kizhner, T.; Ruderfer, I.; Fux, L.; Nataf, Y.; Bartfeld, D.; Ariel, T.; Gingis-Velitski, S.; Hanania, U.; et al. Large-scale production of pharmaceutical proteins in plant cell culture—The protalix experience. Plant Biotechnol. J. 2015, 13, 1199–1208. [Google Scholar] [CrossRef] [PubMed]
- Santos, R.B.; Abranches, R.; Fischer, R.; Sack, M.; Holland, T. Putting the spotlight back on plant suspension culture. Front. Plant Sci. 2016, 7, 297–318. [Google Scholar] [CrossRef] [PubMed]
- Horsch, R.B.; Fry, J.E.; Hoffmann, N.L.; Eichholtz, D.; Rogers, S.G.; Fraley, R.T. A Simple and General-Method for Transferring Genes into Plants. Science 1985, 227, 1229–1231. [Google Scholar]
- Hamilton, C.M. A binary-BAC system for plant transformation with high-molecular-weight DNA. Gene 1997, 200, 107–116. [Google Scholar] [CrossRef]
- Montgomery, D. Design and Analysis of Experiments; Wiley: New York, NY, USA, 1997. [Google Scholar]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Agrobacterium Strain | Gene Silencing Suppressor | Virus | Potential Modes of Action |
|---|---|---|---|
| EHA105 | 2b | Cucumber mosaic virus (CMV) | Required for host-specific movement of PTGS signals [29]; Interacts with components of the RISC machinery to reduce ARGONAUTE (AGO) cleavage activity [30]. |
| GV2260 | coat protein (CP) (also referred to as p38) | Turnip crinkle virus (TCV) | TCV CP functions to suppress RNA silencing at an early initiation step of PTGS by interfering with the function of the Dicer-like RNase in plants [31]. |
| GV2260 | HC-Pro | Tobacco etch virus (TEV) | Functions by binding to double-stranded siRNA (ds siRNA) and inhibits their unwinding to single-stranded siRNA (ss siRNA) [13]. |
| EHA105 | P1 | Rice yellow mottle virus (RYMV) | P1 of RYMV is required for systemic virus spread and movement [32]. |
| GV2260 | P10 | Grapevine virus A (GVA) | P10 of GVA reduces the levels of ss siRNAs by sequestering ds siRNAs [33]. |
| EHA105 | P19 | Tomato bushy stunt virus (TBSV) | P19 of TBSV functions by binding to and sequestering ds siRNA, reducing the ss siRNA level [3,10,14]. |
| GV2260 | P21 | Beet yellow virus (BYV) | P21 silencing suppression mechanism is similar to P19 for inhibiting silencing pathways by binding ds siRNA [34]. |
| GV2260 | P24 | Grapevine leafroll associated virus-2 (GLRaV-2) | P24 of GLRaV-2 is capable of preventing induction of silencing by double-stranded inverted repeat, reducing the ds RNA levels [35]. |
| EHA105 | P25 | Potato virus X (PVX) | P25 of PVX is responsible for cell-to-cell movement of PTGS signals and blocks systemic silencing [36]. |
| Run No. | Viral Gene Silencing Suppressors (μg-Agrobacterium/g-Plant Cell Biomass) | Extracellular Total rAAT (μg/L) | Extracellular Functional rAAT (μg/L) | Functional/Total rAAT (%) | ||||
|---|---|---|---|---|---|---|---|---|
| CP | P19 | P21 | P24 | P25 | ||||
| 1 | 0 | 0 | 0 | 0 | 10 | 327.7 | 100.1 | 30.54 |
| 2 | 10 | 0 | 0 | 0 | 0 | 227.8 | 92.9 | 40.77 |
| 3 | 0 | 10 | 0 | 0 | 0 | 278.3 | 95.2 | 34.22 |
| 4 | 10 | 10 | 0 | 0 | 10 | 247.4 | 104.4 | 42.22 |
| 5 | 0 | 0 | 10 | 0 | 0 | 255.3 | 120.8 | 47.31 |
| 6 | 10 | 0 | 10 | 0 | 10 | 302.9 | 112.0 | 36.98 |
| 7 | 0 | 10 | 10 | 0 | 10 | 307.7 | 85.7 | 27.86 |
| 8 | 10 | 10 | 10 | 0 | 0 | 326.0 | 134.1 | 41.14 |
| 9 | 0 | 0 | 0 | 10 | 0 | 265.0 | 103.5 | 39.05 |
| 10 | 10 | 0 | 0 | 10 | 10 | 245.4 | 114.5 | 46.65 |
| 11 | 0 | 10 | 0 | 10 | 10 | 311.1 | 108.1 | 34.76 |
| 12 | 10 | 10 | 0 | 10 | 0 | 556.2 | 250.3 | 45.00 |
| 13 | 0 | 0 | 10 | 10 | 10 | 539.4 | 243.1 | 45.07 |
| 14 | 10 | 0 | 10 | 10 | 0 | 347.4 | 194.4 | 55.95 |
| 15 | 0 | 10 | 10 | 10 | 0 | 339.9 | 110.5 | 32.51 |
| 16 | 10 | 10 | 10 | 10 | 10 | 301.8 | 120.1 | 39.79 |
| Control | 0 | 0 | 0 | 0 | 0 | 121.8 | 13.1 | 10.79 |
| Run No. | Viral Gene Silencing Suppressors (μg-Agrobacterium/g-Plant Cell) | Extracellular Total rAAT (μg/L) | Extracellular Functional rAAT (μg/L) | ||||
|---|---|---|---|---|---|---|---|
| CP | P19 | P24 | Experimental Value | Predicted Value | Experimental Value | Predicted Value | |
| 1 | 8 | 8 | 8 | 332.8 | 363.3 | 176.3 | 157.6 |
| 2 | 25 | 8 | 8 | 359.1 | 320.6 | 167.5 | 184.8 |
| 3 | 8 | 25 | 8 | 361.4 | 306.1 | 157.8 | 164.6 |
| 4 | 25 | 25 | 8 | 260.0 | 252.1 | 192.1 | 195.1 |
| 5 | 8 | 8 | 25 | 250.2 | 211.1 | 159.4 | 172.2 |
| 6 | 25 | 8 | 25 | 325.4 | 333.8 | 184.9 | 193.7 |
| 7 | 8 | 25 | 25 | 351.4 | 343.0 | 214.0 | 212.4 |
| 8 | 25 | 25 | 25 | 618.4 | 454.5 | 301.6 | 237.3 |
| 9 | 0 | 17 | 17 | 321.9 | 302.2 | 193.7 | 164.9 |
| 10 | 34 | 17 | 17 | 304.3 | 371.0 | 203.9 | 217.0 |
| 11 | 17 | 0 | 17 | 288.6 | 252.1 | 184.1 | 144.8 |
| 12 | 17 | 34 | 17 | 232.1 | 315.6 | 171.9 | 195.4 |
| 13 | 17 | 17 | 0 | 231.5 | 211.2 | 176.1 | 142.8 |
| 14 | 17 | 17 | 34 | 194.0 | 261.3 | 181.9 | 199.5 |
| 15 | 17 | 17 | 17 | 323.7 | 435.5 | 168.2 | 226.6 |
| Control | 0 | 0 | 0 | 216.5 | 238.1 | 15.5 | 30.2 |
| CMViva System | Functional rAAT/TSP (%) | Total rAAT/TSP (%) | Functional AAT/Total AAT (%) |
|---|---|---|---|
| Transient expression in whole intact plant leaves, without p19, topical infiltration and induction [25] | 0.16 | 0.57 | 28.1 |
| Transient expression in whole intact plant leaves, with p19, topical infiltration and induction [25] | 1.2 | 1.7 | 70.5 |
| Transient expression in detached plant leaves, without p19, vacuum infiltration and induction [43] | 0.061 | 0.364 | 16.7 |
| Transient expression in detached plant leaves, with p19, vacuum infiltration and induction [43] | 1.196 | 4.068 | 29.4 |
| Transient expression in detached plant leaves, with p19, optimized vacuum infiltration and induction [44] | 2.6 | 4.1 | 63.4 |
| Transgenic plant cell cultures in shake flask, without p19 transient expression (extracellular rAAT yield), this work | 0.0195 | 0.163 | 11.9 |
| Transgenic plant cell cultures in shake flask, with p19 transient expression (extracellular rAAT yield), this work | 0.110 | 0.422 | 26.1 |
| Transgenic plant cell cultures in shake flask, with transient co-expression of CP, P19 and P24 (extracellular rAAT yield), this work | 0.336 | 0.673 | 49.9 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, T.-K.; Falk, B.W.; Dandekar, A.M.; McDonald, K.A. Enhancement of Recombinant Protein Production in Transgenic Nicotiana benthamiana Plant Cell Suspension Cultures with Co-Cultivation of Agrobacterium Containing Silencing Suppressors. Int. J. Mol. Sci. 2018, 19, 1561. https://doi.org/10.3390/ijms19061561
Huang T-K, Falk BW, Dandekar AM, McDonald KA. Enhancement of Recombinant Protein Production in Transgenic Nicotiana benthamiana Plant Cell Suspension Cultures with Co-Cultivation of Agrobacterium Containing Silencing Suppressors. International Journal of Molecular Sciences. 2018; 19(6):1561. https://doi.org/10.3390/ijms19061561
Chicago/Turabian StyleHuang, Ting-Kuo, Bryce W. Falk, Abhaya M. Dandekar, and Karen A. McDonald. 2018. "Enhancement of Recombinant Protein Production in Transgenic Nicotiana benthamiana Plant Cell Suspension Cultures with Co-Cultivation of Agrobacterium Containing Silencing Suppressors" International Journal of Molecular Sciences 19, no. 6: 1561. https://doi.org/10.3390/ijms19061561