Responsiveness of Carnosine Homeostasis Genes in the Pancreas and Brain of Streptozotocin-Treated Mice Exposed to Dietary Carnosine

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Model Validation: Body Weight and Glucose Levels in STZ-Treated Hyperglycemic Mice that Underwent CAR Oral Administration for Two Weeks
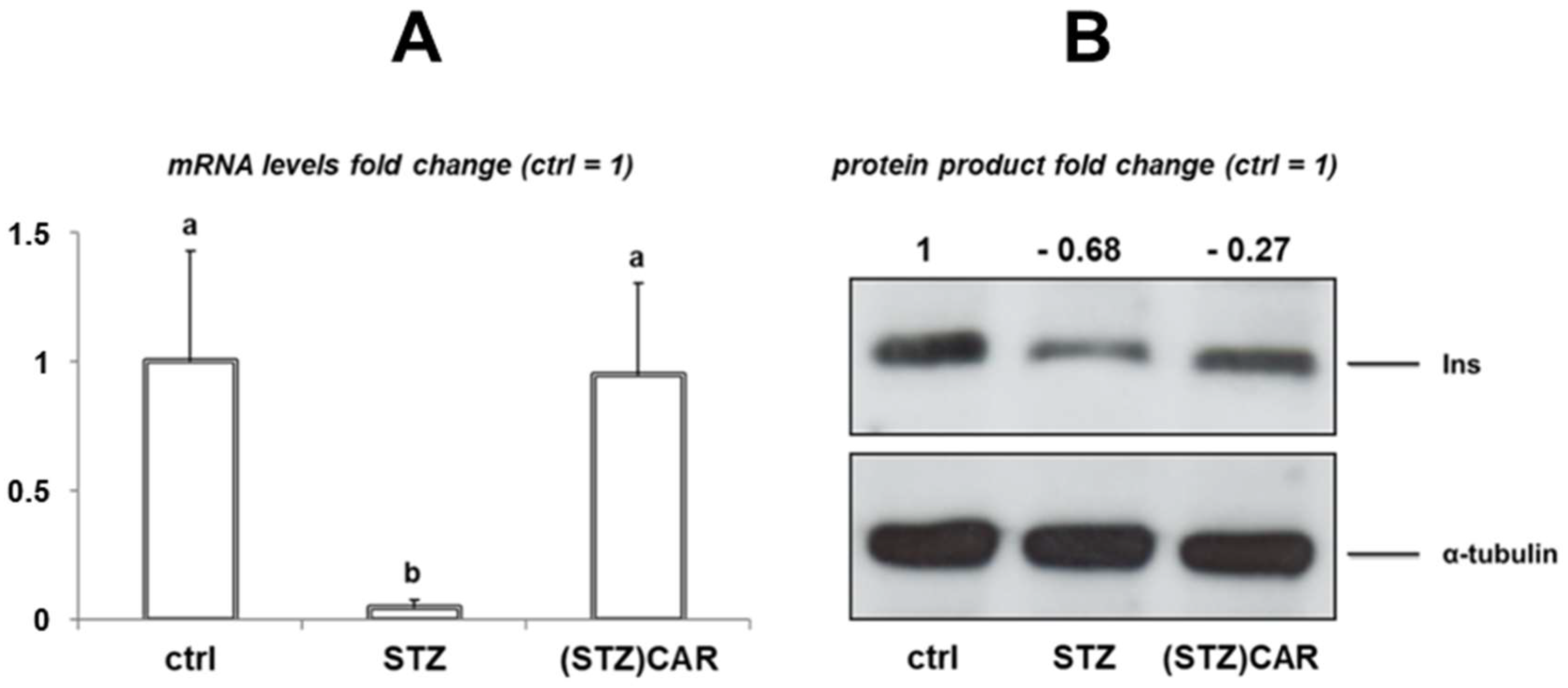
2.2. Model Validation: Expression of the Insulin Gene Products in the Pancreas of STZ-Treated Hyperglycemic Mice that Underwent CAR Oral Administration for Two Weeks
2.3. Expression Variations of mRNAs Related to CAR Homeostasis in the Pancreas and Brain of STZ-Treated Mice that Underwent (Dietary) CAR Oral Administration for Two Weeks
2.4. Detection of CAR Content in Pancreas and Brain Extracts
3. Discussion
4. Materials and Methods
4.1. Animal Handling, Ethical Approval and Treatments
4.2. RNA/Protein Isolation from Tissues
4.3. Primer Design and Real-Time PCR Assays
4.4. Western Blotting
4.5. CAR Content Measurement by High-Performance Liquid Chromatography (HPLC)
4.6. Statistical Analysis
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Boldyrev, A.A. Does carnosine possess direct antioxidant activity? Int. J. Biochem. 1993, 25, 1101–1107. [Google Scholar] [CrossRef]
- Boldyrev, A.A.; Koldobski, A.; Kurella, E.; Maltseva, V.; Stvolinski, S. Natural histidine-containing dipeptide carnosine as a potent hydrophilic antioxidant with membrane stabilizing function. A biomedical aspect. Mol. Chem. Neuropathol. 1993, 19, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Boldyrev, A.A.; Aldini, G.; Derave, W. Physiology and pathophysiology of carnosine. Physiol. Rev. 2013, 93, 1803–1845. [Google Scholar] [CrossRef] [PubMed]
- Nagai, K.; Niijima, A.; Yamano, T.; Otani, H.; Okumra, N.; Tsuruoka, N.; Nakai, M.; Kiso, Y. Possible role of l-carnosine in the regulation of blood glucose through controlling autonomic nerves. Exp. Biol. Med. 2003, 228, 1138–1145. [Google Scholar] [CrossRef]
- Kubomura, D.; Matahira, Y.; Nagai, K.; Niijima, A. Effect of anserine ingestion on hyperglycemia and the autonomic nerves in rats and humans. Nutr. Neurosci. 2010, 13, 183–188. [Google Scholar] [CrossRef] [PubMed]
- Sauerhofer, S.; Yuan, G.; Braun, G.S.; Deinzer, M.; Neumaier, M.; Gretz, N.; Floege, J.; Kriz, W.; van der Woude, F.; Moeller, M.J. l-carnosine, a substrate of carnosinase-1, influences glucose metabolism. Diabetes 2007, 56, 2425–2432. [Google Scholar] [CrossRef] [PubMed]
- Miceli, V.; Pampalone, M.; Frazziano, G.; Grasso, G.; Rizzarelli, E.; Ricordi, C.; Casu, A.; Iannolo, G.; Conaldi, P.G. Carnosine protects pancreatic beta cells and islets against oxidative stress damage. Mol. Cell. Endocrinol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.T.; Hsu, C.C.; Lin, M.H.; Liu, K.S.; Yin, M.C. Histidine and carnosine delay diabetic deterioration in mice and protect human low density lipoprotein against oxidation and glycation. Eur. J. Pharmacol. 2005, 513, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Soliman, K.; Mohamed, A.; Metwally, N. Attenuation of some metabolic deteriorations induced by diabetes mellitus using carnosine. J. Appl. Sci. 2007, 7, 2252–2260. [Google Scholar]
- Elbarbary, N.S.; Ismail, E.A.R.; El-Naggar, A.R.; Hamouda, M.H.; El-Hamamsy, M. The effect of 12 weeks carnosine supplementation on renal functional integrity and oxidative stress in pediatric patients with diabetic nephropathy: A randomized placebo-controlled trial. Pediatr. Diabetes 2018, 19, 470–477. [Google Scholar] [CrossRef] [PubMed]
- Houjeghani, S.; Kheirouri, S.; Faraji, E.; Jafarabadi, M.A. l-Carnosine supplementation attenuated fasting glucose, triglycerides, advanced glycation end products, and tumor necrosis factor-alpha levels in patients with type 2 diabetes: A double-blind placebo-controlled randomized clinical trial. Nutr. Res. 2018, 49, 96–106. [Google Scholar] [CrossRef] [PubMed]
- Hipkiss, A.R. Carnosine, diabetes and Alzheimer’s disease. Expert Rev. Neurother. 2009, 9, 583–585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pavlin, M.; Rossetti, G.; De Vivo, M.; Carloni, P. Carnosine and Homocarnosine Degradation Mechanisms by the Human Carnosinase Enzyme CN1: Insights from Multiscale Simulations. Biochemistry 2016, 55, 2772–2784. [Google Scholar] [CrossRef] [PubMed]
- Hipkiss, A.R.; Baye, E.; de Courten, B. Carnosine and the processes of ageing. Maturitas 2016, 93, 28–33. [Google Scholar] [CrossRef] [PubMed]
- Riedl, E.; Pfister, F.; Braunagel, M.; Brinkkötter, P.; Sternik, P.; Deinzer, M.; Bakker, S.J.; Henning, R.H.; van den Born, J.; Krämer, B.K.; et al. Carnosine Prevents Apoptosis of Glomerular Cells and Podocyte Loss in STZ Diabetic Rats. Cell Physiol. Biochem. 2011, 28, 279–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albrecht, T.; Schilperoort, M.; Zhang, S.; Braun, J.D.; Qiu, J.; Rodriguez, A.; Pastene, D.O.; Krämer, B.K.; Köppel, H.; Baelde, H.; et al. Carnosine Attenuates the Development of both Type 2 Diabetes and Diabetic Nephropathy in BTBR ob/ob Mice. Sci. Rep. 2017, 7, 44492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janssen, B.; Hohenadel, D.; Brinkkoetter, P.; Peters, V.; Rind, N.; Fischer, C.; Rychlik, I.; Cerna, M.; Romzova, M.; de Heer, E.; et al. Carnosine as a protective factor in diabetic nephropathy: Association with a leucine repeat of the carnosinase gene CNDP1. Diabetes 2005, 54, 2320–2327. [Google Scholar] [CrossRef] [PubMed]
- Yadav, A.K.; Sinha, N.; Kumar, V.; Bhansali, A.; Dutta, P.; Jha, V. Association of CTG repeat polymorphism in carnosine dipeptidase 1 (CNDP1) gene with diabetic nephropathy in north Indians. Indian J. Med. Res. 2016, 144, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Peters, V.; Lanthaler, B.; Amberger, A.; Fleming, T.; Forsberg, E.; Hecker, M.; Wagner, A.H.; Yue, W.W.; Hoffmann, G.F.; Nawroth, P.; et al. Carnosine metabolism in diabetes is altered by reactive metabolites. Amino Acids 2015, 47, 2367–2376. [Google Scholar] [CrossRef] [PubMed]
- Vardarli, I.; Baier, L.J.; Hanson, R.L.; Akkoyun, I.; Fischer, C.; Rohmeiss, P.; Basci, A.; Bartram, C.R.; van der Woude, F.J.; Janssen, B. Gene for susceptibility to diabetic nephropathy in type 2 diabetes maps to 18q22.3–23. Kidney Int. 2002, 62, 2176–2183. [Google Scholar] [CrossRef] [PubMed]
- Bowden, D.W.; Colicigno, C.J.; Langefeld, C.D.; Sale, M.M.; Williams, A.; Anderson, P.J.; Rich, S.S.; Freedman, B.I. A genome scan for diabetic nephropathy in African Americans. Kidney Int. 2004, 66, 1517–1526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertinaria, M.; Rolando, B.; Giorgis, M.; Montanaro, G.; Guglielmo, S.; Buonsanti, M.F.; Carabelli, V.; Gavello, D.; Daniele, P.G.; Fruttero, R.; et al. Synthesis, physicochemical characterization, and biological activities of new carnosine derivatives stable in human serum as potential neuroprotective agents. J. Med. Chem. 2011, 54, 611–621. [Google Scholar] [CrossRef] [PubMed]
- Xie, R.X.; Li, D.W.; Liu, X.C.; Yang, M.F.; Fang, J.; Sun, B.L.; Zhang, Z.Y.; Yang, X.Y. Carnosine Attenuates Brain Oxidative Stress and Apoptosis After Intracerebral Hemorrhage in Rats. Neurochem. Res. 2017, 42, 541–551. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.H.; Ma, Q.; Wang, X.; Xu, G.H. Protective effects of beef decoction rich in carnosine on cerebral ischemia injury by permanent middle cerebral artery occlusion in rats. Exp. Ther. Med. 2018, 15, 1321–1329. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Herculano, B.; Tamura, M.; Ohba, A.; Shimatani, M.; Kutsuna, N.; Hisatsune, T. β-alanyl-l-histidine rescues cognitive deficits caused by feeding a high fat diet in a transgenic mouse model of Alzheimer’s disease. J. Alzheimers Dis. 2013, 33, 983–997. [Google Scholar] [CrossRef] [PubMed]
- Manhiani, P.S.; Northcutt, J.K.; Han, I.; Bridges, W.C.; Scott, T.R.; Dawson, P.L. Effect of stress on carnosine levels in brain, breast, and thigh of broilers. Poult. Sci. 2011, 90, 2348–2354. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GENE (Mus musclus) | mRNA RefSeq | Forward Primer 5′–3′ (Tm) | Reverse Primer 5′–3′ (Tm) | PCR Size (bp) |
|---|---|---|---|---|
| Carns1 | NM_134148 | GCGGCGTCAGCAAGAAGTT(59°) | CACCAAGCAGTCATCCCAGAA(60°) | 136 |
| Cndp2 | NM_023149.2 | TTCAAGGTGTACATGGGC(54°) | AAAGGTCAAGGTCACAGGA(55°) | 162 |
| Cndp1 | NM_177450.4 | CCTAGAAGAATACCAGAAGAGC(58°) | GGGACTAGACGGATTGAAA(55°) | 216 |
| Slc15a2/Pept2 | NM_021301.3 | CATGAAATCTGTGCTCCAGG(57°) | AGGAGGCAGGAAAACAAAA(52°) | 126 |
| Ins1 | NM_008386.3 | TCAGAGACCATCAGCAAGCA(56°) | TCTCTACCTGGTGTGTGGG(55°) | 167 |
| 28S | NR_003279.1 | CGTGAGACAGGTTAGTTTTAC(51°) | ATCCCACAGATGGTAGCTTC(53°) | 143 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barca, A.; Gatti, F.; Spagnolo, D.; Ippati, S.; Vetrugno, C.; Verri, T. Responsiveness of Carnosine Homeostasis Genes in the Pancreas and Brain of Streptozotocin-Treated Mice Exposed to Dietary Carnosine. Int. J. Mol. Sci. 2018, 19, 1713. https://doi.org/10.3390/ijms19061713
Barca A, Gatti F, Spagnolo D, Ippati S, Vetrugno C, Verri T. Responsiveness of Carnosine Homeostasis Genes in the Pancreas and Brain of Streptozotocin-Treated Mice Exposed to Dietary Carnosine. International Journal of Molecular Sciences. 2018; 19(6):1713. https://doi.org/10.3390/ijms19061713
Chicago/Turabian StyleBarca, Amilcare, Francesca Gatti, Daniela Spagnolo, Stefania Ippati, Carla Vetrugno, and Tiziano Verri. 2018. "Responsiveness of Carnosine Homeostasis Genes in the Pancreas and Brain of Streptozotocin-Treated Mice Exposed to Dietary Carnosine" International Journal of Molecular Sciences 19, no. 6: 1713. https://doi.org/10.3390/ijms19061713