1. Introduction
CCAAT/enhancer-binding proteins (C/EBPs) are transcription factors that activate the expression of target genes through interaction with response elements within their promoters [
1]. There are six members of C/EBP family, and they regulate differentiation process in various tissues [
2]. The first transcription factor in this family, C/EBPα, was isolated from the rat liver and it appeared to be important for adipocyte differentiation [
3]. C/EBPs are modular proteins consisting of an activation domain, a DNA binding domain, and a leucine-rich dimerization domain that is responsible for forming dimers with other members of the family [
4]. In order to activate transcription, the C/EBP dimers bind to the consensus sequence 5′-TT/GNNGNAAT/G-3′ in promoter regions of target genes. For three out of six genes encoding C/EBP family members, alternative protein products are translated, due to a leaky ribosomal scanning mechanism. Some of them lack the N-terminal activation domains and exert inhibitory functions, presumably by a dominant negative mechanism [
5].
Hematopoiesis is a process in which all blood elements are formed from multipotential hematopoietic stem cells (HSCs). In the process of hematopoiesis, the HSCs and their progeny interact with the bone marrow stromal cells and they are stimulated by the numerous growth factors that are secreted in the bone marrow environment. The eventual cell fate during hematopoiesis is governed by spatiotemporal fluctuations in transcription factor concentrations, which either cooperate or compete in driving target gene expression [
6]. Some members of C/EBP family of transcription factors are important at certain steps of hematopoiesis [
7]. C/EBPα appears in differentiating cells at the stage of early progenitors with lymphoid and myeloid potential and then reappears only in the cells that are differentiating into granulocytes [
8]. C/EBPα-deficient mice show disturbances in monocyte and neutrophil development [
9,
10]. High level of C/EBPβ leads to monocyte and macrophage development [
11,
12], while high level of C/EBPε leads to neutrophil differentiation [
13]. The role of C/EBPδ in blood cells development is less defined, since
CEBPD−/− mice did not reveal any blood disturbances [
14]. It has been documented that C/EBPδ regulates expression of genes important for granulocyte function [
15].
However, the most important factors that drive blood cells development are cytokines [
16], some ligands for nuclear receptors can also modulate cell fate during hematopoiesis [
7]. The best described in this respect are ligands for retinoic acid receptors (RARs). Active metabolites of vitamin A are natural ligands for RARs. A dominating retinoic acid (RA) metabolite is all-
trans-RA (ATRA), which binds with high affinity to all RARs (α, β, and γ) [
17]. During embryogenesis, ATRA causes the appearance of hematopoietic progenitors from the hemogenic endothelium [
18], while in adults, it is important for the differentiation of granulocytes, as well as B and T lymphocytes [
19]. This activity of ATRA has been used in clinics. The most clinically significant application of ATRA is to treat a rare subtype of an acute myeloid leukemia (AML), called acute promyelocytic leukemia (APL). At the first description this subtype was considered the most difficult to treat [
20], while it is now considered as highly curable using the combination of ATRA and anthracycline-based chemotherapy [
21]. Another ligand for the nuclear receptor which influences hematopoiesis is an active metabolite of vitamin D. The correct physiological concentrations of 1,25-dihydroxyvitamin D (1,25D), which is a natural ligand for vitamin D receptor (VDR), are necessary to induce markers of monocytic differentiation in HSCs [
22]. The expression of the
VDR gene is higher at the early steps of hematopoiesis than at later stages and in mature blood cells [
23]. However, both these ligands do not seem indispensable for blood cells development since RARα-deficient and VDR-deficient mice show no defects in hematopoiesis [
24,
25]. The possibility that these nuclear receptors can, in some aspects, functionally compensate each other should be considered.
It has been documented in the past that members of C/EBP family of transcription factors can be upregulated in blood cells by an exposure to RA, 1,25D, or to their active analogs. For example, the expression of C/EBPε mRNA and protein increases in AML cells exposed to 9-
cis–RA or 1,25D analog (KH1060) [
26]. The gene encoding C/EBPβ has been shown to be a target for VDR regulation [
27] and all isoforms of this transcription factor are increased in AML cells exposed to 1,25D or to analogs of 1,25D [
11,
28]. This gene is also strongly upregulated in AML cells that were exposed to ATRA [
29]. 1,25D induces a transient increase of C/EBPα [
11], which also participates in the ATRA-induced differentiation of AML cells [
30].
In this study, we addressed a question of whether the lack of one of the nuclear receptors mentioned above could be compensated by the other in terms of
CEBP activation. Therefore, we used four cell lines in our study, with different expression of retinoic acid receptor α (
RARA) or
VDR. In HL60 cells,
VDR expression is on a high level and
RARA is moderate [
31]. For the purpose of this study, we silenced the expression of
VDR in HL60 cells using shRNA. In contrast to HL60 cells, KG1 cell express high levels of
RARA, but low of
VDR [
31]. The effects of
RARA silencing were studied using a sub-line KG1-RARα(−).
3. Discussion
The process of hematopoiesis leads to the acquisition of immune functions by terminally differentiated cells. Lineage selection within hematopoiesis depends on the appropriate levels of key transcription factors, which are regulated in response to numerous hematopoietic cytokines and interactions with bone marrow environment [
38]. Transcription factors C/EBPα, C/EBPβ, and C/EBPδ have been demonstrated in granulocytes, monocytes, and eosinophils, as well as in myeloid progenitor cells [
39]. C/EBPε has been identified as a critical regulator of terminal granulopoiesis [
13]. Many genes that are important for myeloid functions contain in their promoters binding sites for C/EBP transcription factors [
5]. In normal hematopoiesis, C/EBP transcription factors are produced in response to coordinated actions of cytokines and upstream transcription factors, and their activity is further modulated by posttranslational modifications [
5].
C/EBPα seems to be the most important for normal blood development, since mutations in
CEBPA gene lead to AML.
CEBPA is mutated in around 13% of all AML patients [
40], and mutations in this gene appear early, indicating at the driver role in leukemogenesis [
41]. This is why the expression of
CEBPA in AML patients has been extensively studied, and it has been shown to be downregulated by another driver of leukemogenesis, namely the AML-ETO fusion protein, which is present in 5–10% of patients with AML [
42]. Later studies have documented that the downregulation of
CEBPA also accompanies AML cases with inv(16), which creates
CBFB-MYH11 gene fusion and occurs in about 10% of AML patients [
43]. All together, the above data show that more than 30% of patients with AML exhibit disturbances in expression of
CEBPA.
It has been shown that both 1,25D and ATRA are able to upregulate expression of C/EBP factors without the addition of hematopoietic cytokines [
11,
26,
27,
29]. Whether all the
CEBP genes are direct targets for either RARα or VDR is not clear. Retinoic acid response elements (RAREs) have been found in the promoter of
CEBPE gene, and not in other genes of this family [
44], but at present, we know that RARα can bind to big variety of RAREs [
45], which are sometimes located in a long distance from the transcription start [
44].
CEBPA and
CEBPB are direct targets of VDR-dependent transcriptional regulation [
36,
37], but it is not sure whether such a mechanism occurs also for
CEBPD.
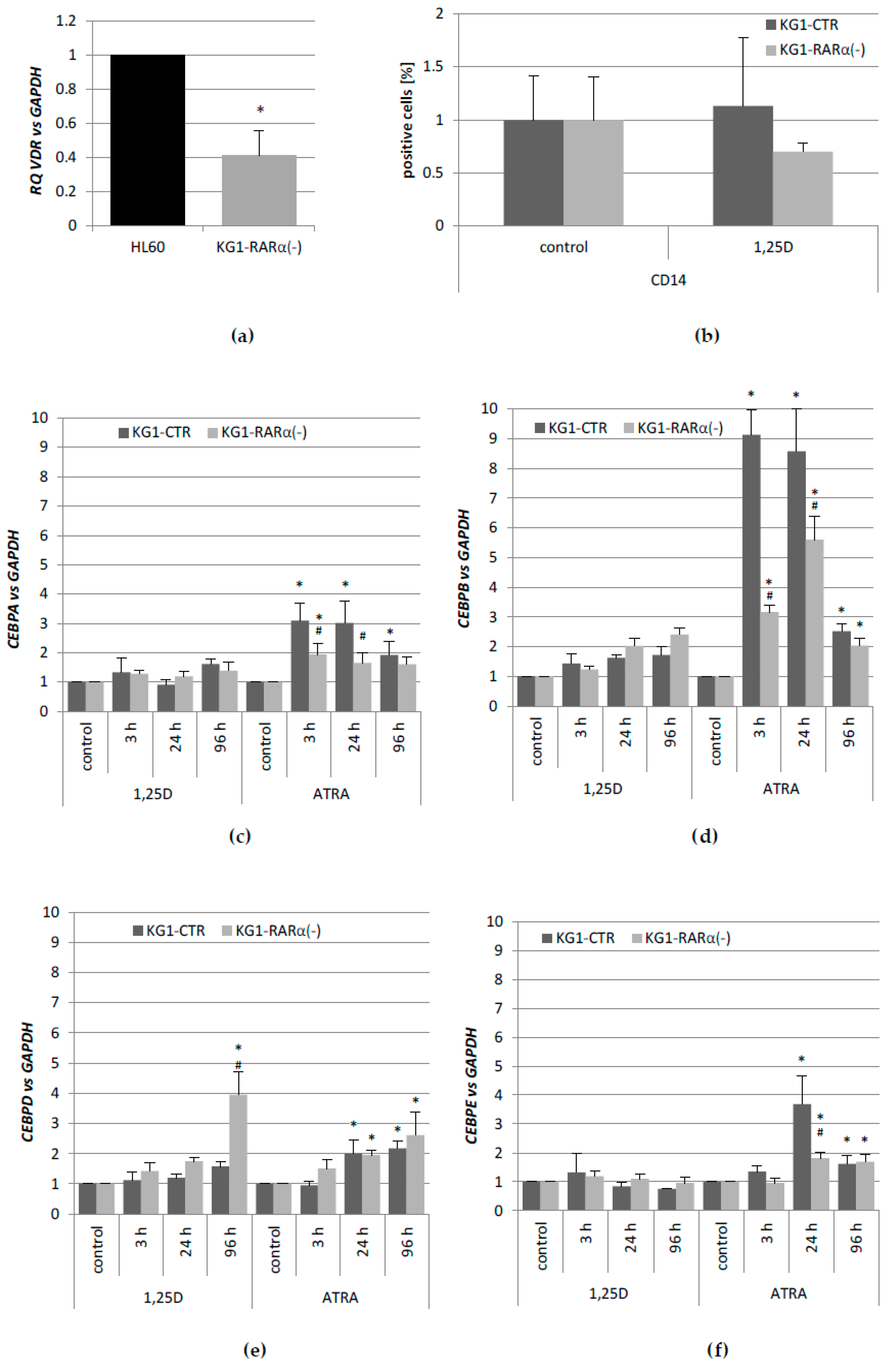
We thus wanted to find out how variable levels of VDR and RARα proteins affect the expression of
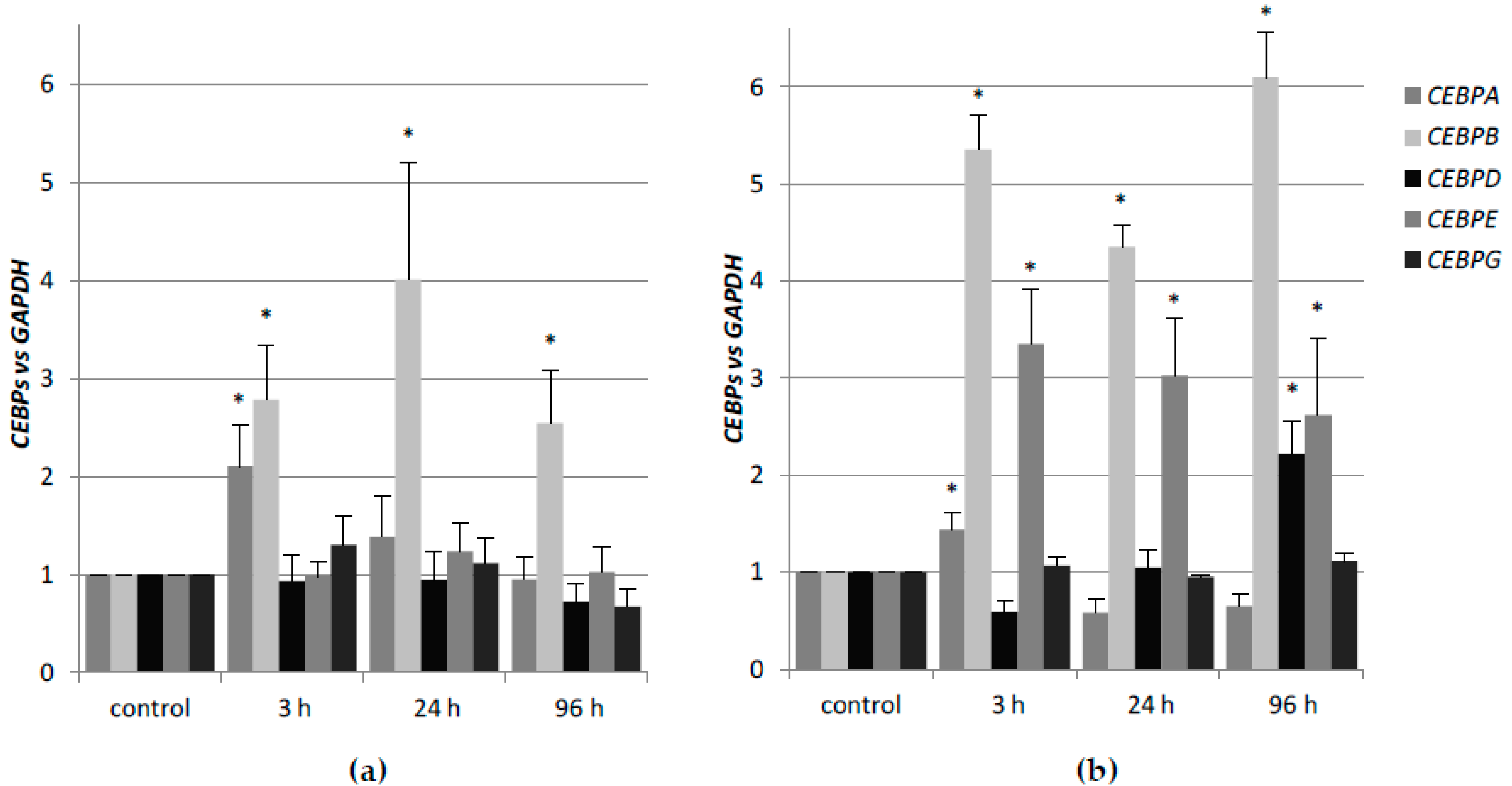
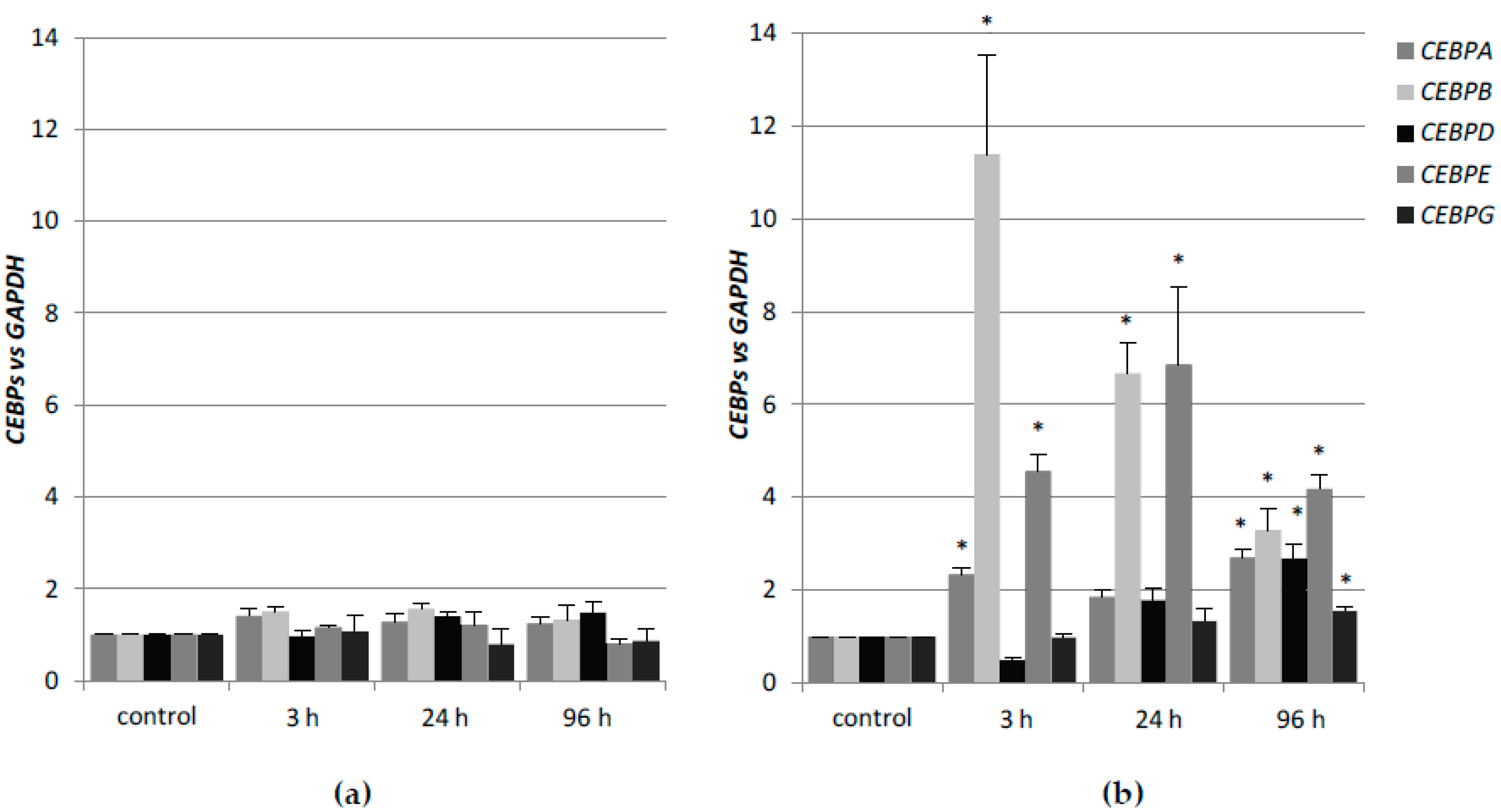
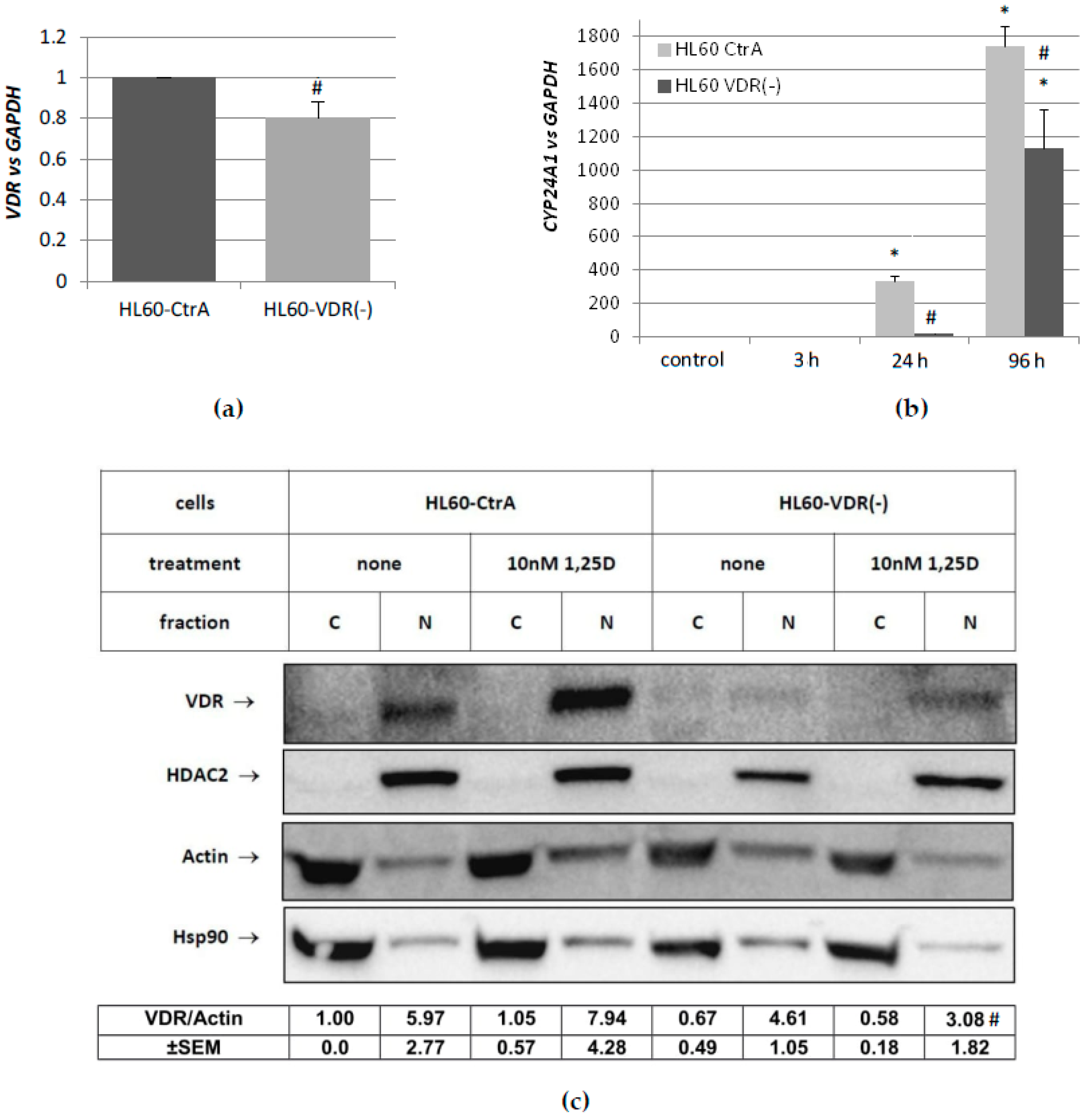
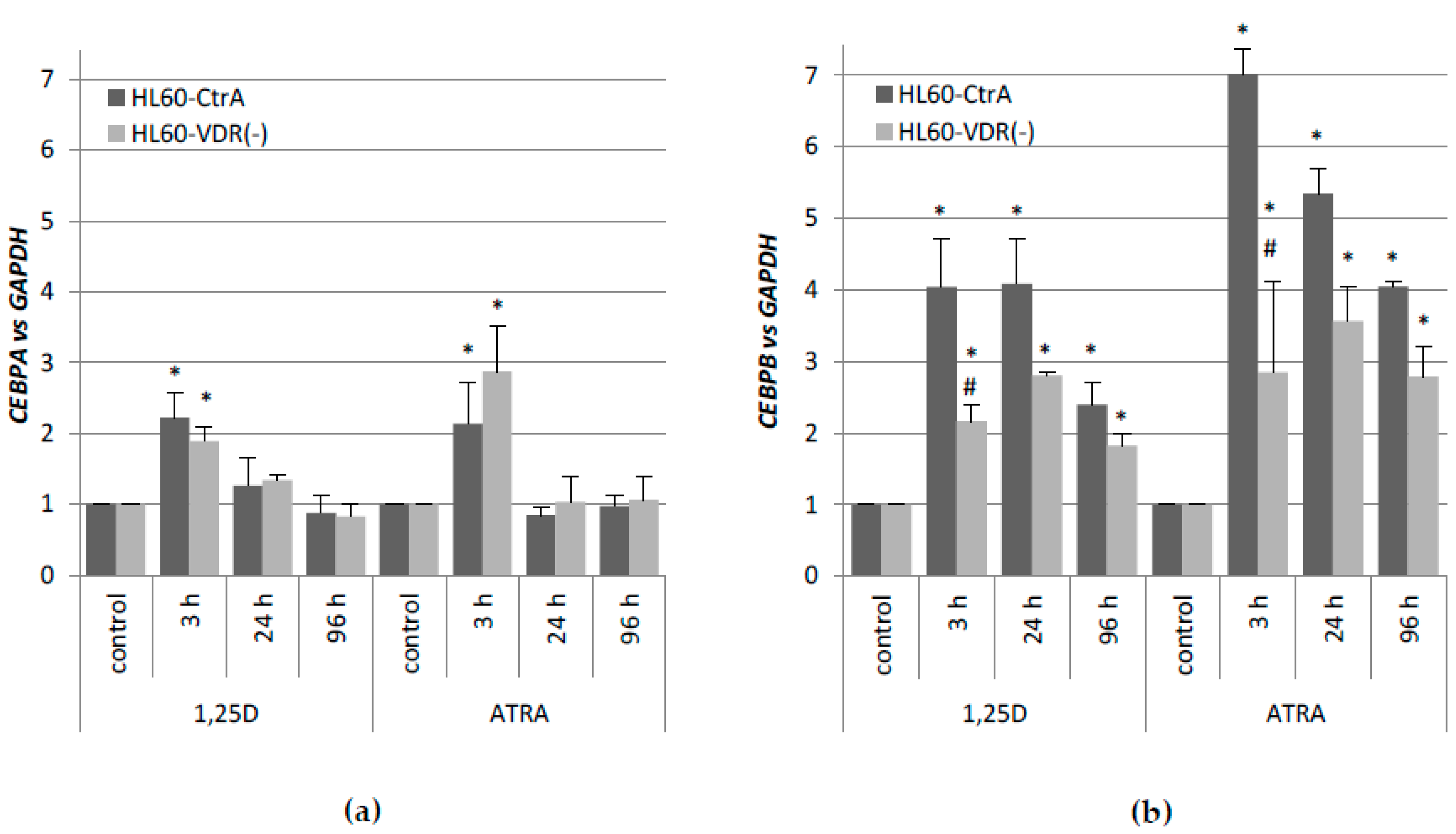
CEBP genes, and whether the lack of one of the nuclear receptors that are mentioned above, could be compensated by the other. In the first place, we determined the expression profiles of these genes after stimulation with 1,25D or ATRA in HL60 and KG1 cells. These cells differ in basal levels of expression of VDR and RARα, and in susceptibility to 1,25D-induced differentiation. The basal levels of
VDR and
RARA mRNA expression in all of the cell lines that were used for the purpose of this research is presented in
Table A5. HL60 cells, which have high level of VDR protein, respond to 1,25D with transient upregulation of
CEBPA, and strong and sustained upregulation of
CEBPB. It appeared that KG1 cells that have low level of VDR protein do not express
CEBP genes in response to 1,25D at all. After the silencing of the
RARA gene in KG1, these cells reduced the responsiveness to ATRA, but started to be responsive to 1,25D, most probably because of an increased expression of VDR [
31]. The restored
VDR expression level was not high enough to upregulate the expression of
CEBPB, but it was sufficient to upregulate
CEBPD. As presented here, the upregulation of
CEBPD alone was not sufficient to complete the myeloid differentiation process. KG1 cells and HL60 cells are both responsive to ATRA, however, due to higher basal expression of
RARA, KG1 cells respond stronger than HL60. In KG1 cells, the upregulation of
CEBPB and
CEBPE is approximately two times higher than in HL60 cells in response to ATRA; however, the kinetics of expression is similar.
Our results suggest that the ability of 1,25D or ATRA to effectively force the final myeloid differentiation of AML cells strongly depends on effective levels of nuclear receptors for these compounds. It also seems that expression of
CEBPB is indispensable for the final effect of myeloid differentiation, and that VDR and RARα do not compensate each other in terms of the induction of
CEBP expression. Our data are in agreement with the earlier findings that strong and sustained expression of
CEBPB, when accompanied by transient expression of
CEBPA leads to the differentiation towards monocytes [
11], while, when accompanied by the sustained expression of
CEBPE, it leads the differentiation process to granulocytes [
46].
4. Materials and Methods
4.1. Cell Lines and Cultures
HL60 cells were from the local cell bank at the Institute of Immunology and Experimental Therapy in Wrocław, and KG1 cells were purchased from the German Resource Center for Biological Material (DSMZ GmbH, Braunschweig, Germany). The cells were grown in RPMI-1640 medium with 10% fetal bovine serum, 100 units/ml penicillin, and 100 µg/mL streptomycin (Sigma, St. Louis, MO, USA), and maintained at standard cell culture conditions.
4.2. Chemicals and Antibodies
1,25D was purchased from Cayman Europe (Tallinn, Estonia) and ATRA was from Sigma-Aldrich (St. Louis, MO, USA). The compounds were dissolved in an absolute ethanol to 1000× final concentrations, and subsequently, diluted in the culture medium to the required concentration.
4.3. cDNA Synthesis and Real-Time PCR
Total RNA was extracted from the cells treated with 1 µM ATRA or 10 nM 1,25D for different time points (3 h, 24 h, 96 h). Briefly, the isolation of total RNA, reverse transcription into cDNA, and Real-time PCR reactions were performed as published before [
33], using CFX Real-time PCR System (Bio-Rad Laboratories Inc., Hercules, CA, USA). The sequences of
GAPDH,
CYP24A1, VDR, and
RARA primers, and the reaction conditions were described previously [
31,
47]. The
CEBPA, CEBPB, CEBPD,
CEBPE, and
CEBPG primers were obtained from RealTimePrimers.com (Real Time Primers, LLC, PA, USA). Their sequences are as follows:
CEBPA: forward 5′-TTGGTGCGTCTAAGATGAGG-3′, reverse 5′-GGCAGGAAACCTCCAAATAA-3′;
CEBPB: forward 5′-AACTCTCTGCTTCTCCCTCTG-3′, reverse 5′-AAGCCCGTAGGAACATCTTT-3′;
CEBPD: forward 5′-ATCGACTTCAGCGCCTACAT-3′, reverse 5′-GCCTTGTGATTGCTGTTGAA-3′;
CEBPE: forward 5′-GAGGAGGTTGCTCAGAGTGG-3′, reverse 5′-TCCTGGCCTATTCAGCAGTT-3′;
CEBPG: forward 5′-GAACAACCCATTTTGCACTC-3′, reverse 5′-TGAAAGCCAGGAACAAAAAG-3′;
APDH: forward 5′-CATGAGAAGTATGACAACAGCCT-3′, reverse 5′-AGTCCTTCCACGATACCAAAGT-3′. Quantification of gene expression was analyzed with either the ∆Cq (to present comparative quantification of expression levels) or with the ∆∆Cq (to present changes in expression induced by treatment) methods using
GAPDH as the endogenous control Primers efficiencies were measured in all of the cell lines using Real-time PCR reaction based on the slope of the standard curve. The results were normalized to primer efficiencies to compare gene expression in different cell lines [
48]. Real-time PCR assays were performed at least in triplicate.
4.4. Flow Cytometry
The expression of CD14 was determined by flow cytometry. The cells were incubated with 10 nM 1,25D for 96 h, then washed, and stained with 1 µL of Phycoerythrin labeled antibody (or the appropriate control immunoglobulins; both from ImmunoTools, Friesoythe, Germany) for 1 h on ice. Next, they were washed with ice-cold PBS supplemented with 0.1% BSA and suspended in 0.5 mL of PBS supplemented with 0.1% BSA prior to analysis on FACS Calibur flow cytometer (Becton–Dickinson, San Jose, CA, USA). Experiments were repeated at least three times. The acquisition parameters were set for an isotype control. Data analysis was performed with the use of WinMDI 2.8 software (freeware by Joseph Trotter).
4.5. Western Blotting
In order to obtain cytosolic and nuclear extracts, 5 × 106 cells/sample were washed and lysed using NE-PER Nuclear and Cytoplasmic Extraction Reagents (Thermo Fisher Scientific Inc., Worcester, MA, USA), according to the user’s manual. Lysates were denatured by adding 5× sample buffer (1/4 volume of the lysate) and boiled for 5 min. 25 µL of each lysate were separated in SDS-PAGE and then electroblotted to PVDF membrane. The membranes were then dried and incubated sequentially with primary and a horseradish peroxidase-conjugated secondary antibody. The protein bands were visualized with a chemiluminescence. Then, the membranes were stripped, dried again, and probed with subsequent antibodies. Western blots were repeated five times.
4.6. Gene Silencing Reagents and Procedure
The
RARA gene silencing in KG1 cells was described before [
31]. The
VDR gene silencing in HL60 cells was performed using shRNA plasmids and Neon
® Transfection System (Invitrogen™, Carlsbad, CA, USA) using control shRNA plasmid-A (sc-108060) and the
VDR shRNA plasmid (sc-106692-SH; both from Santa Cruz Biotechnology, Inc., Dallas, TX, USA). The procedure of electrotransfection by Neon
® Transfection System was described before [
49].
4.7. Statistical Analysis
For statistical analysis one-way ANOVA was used to test the null hypothesis that the samples in two or more groups are drawn from populations with the same mean values. When the ANOVA test had shown that the null hypothesis is not true, the Student’s
t-test for independent samples was used to analyze the differences between the pairs of groups (Excel, Microsoft Office and free ANOVA Calculator:
http://www.danielsoper.com/statcalc3/calc.aspx?id=43).




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}




